
Dietary Sodium Butyrate Supplementation Enhances Silkworm Silk Yield by Simultaneously Promoting Larval Growth and Silk Gland Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insect Material
2.2. Assessment of NaB on Silkworm Growth
2.3. Silk Gland Phenotype Observation and Measurement
2.4. DNA Content Determination
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Comparison of Economic Characteristics
2.7. Statistical Analysis
3. Results
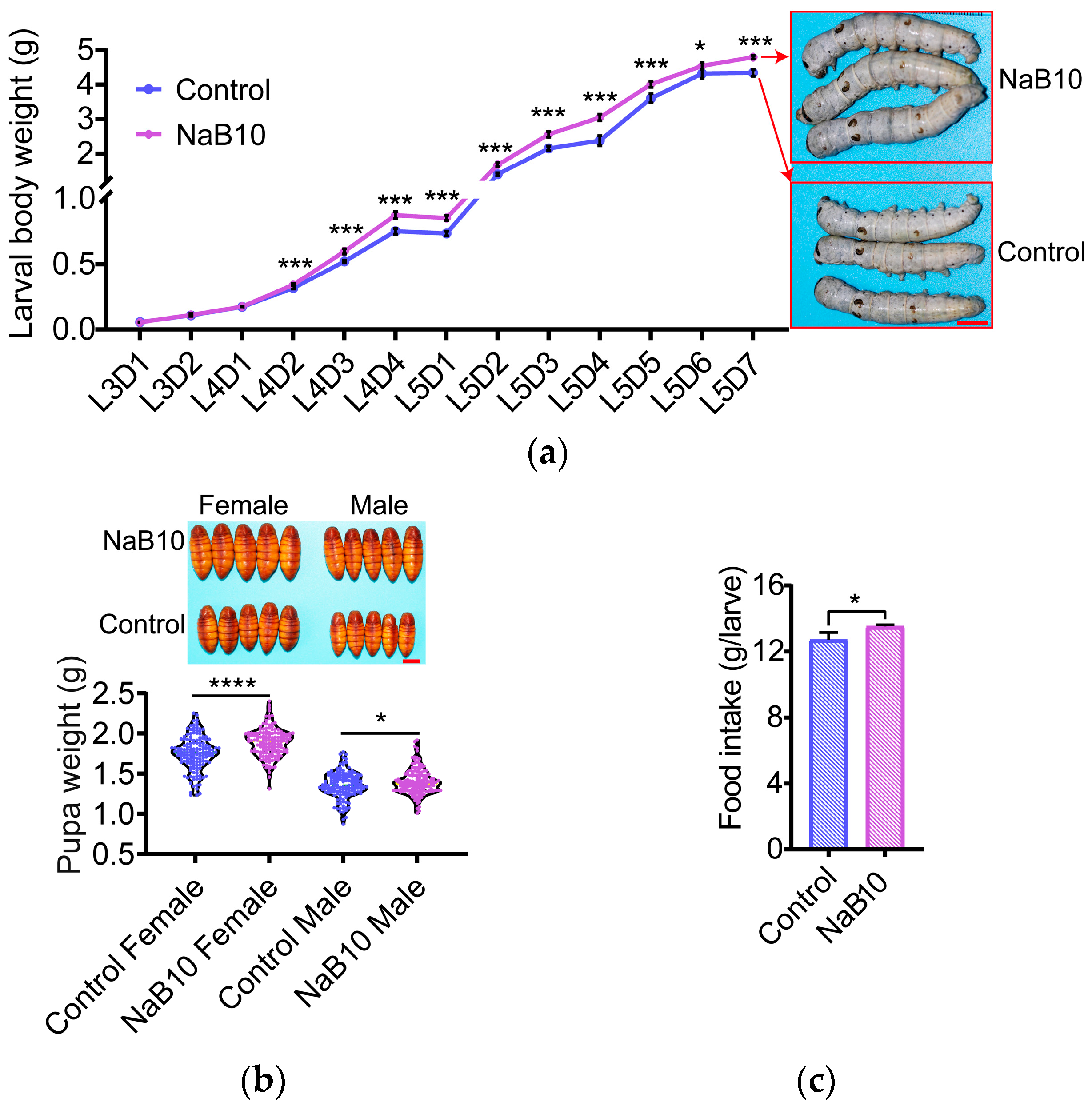
3.1. Effects of Dietary NaB on Silkworm Growth
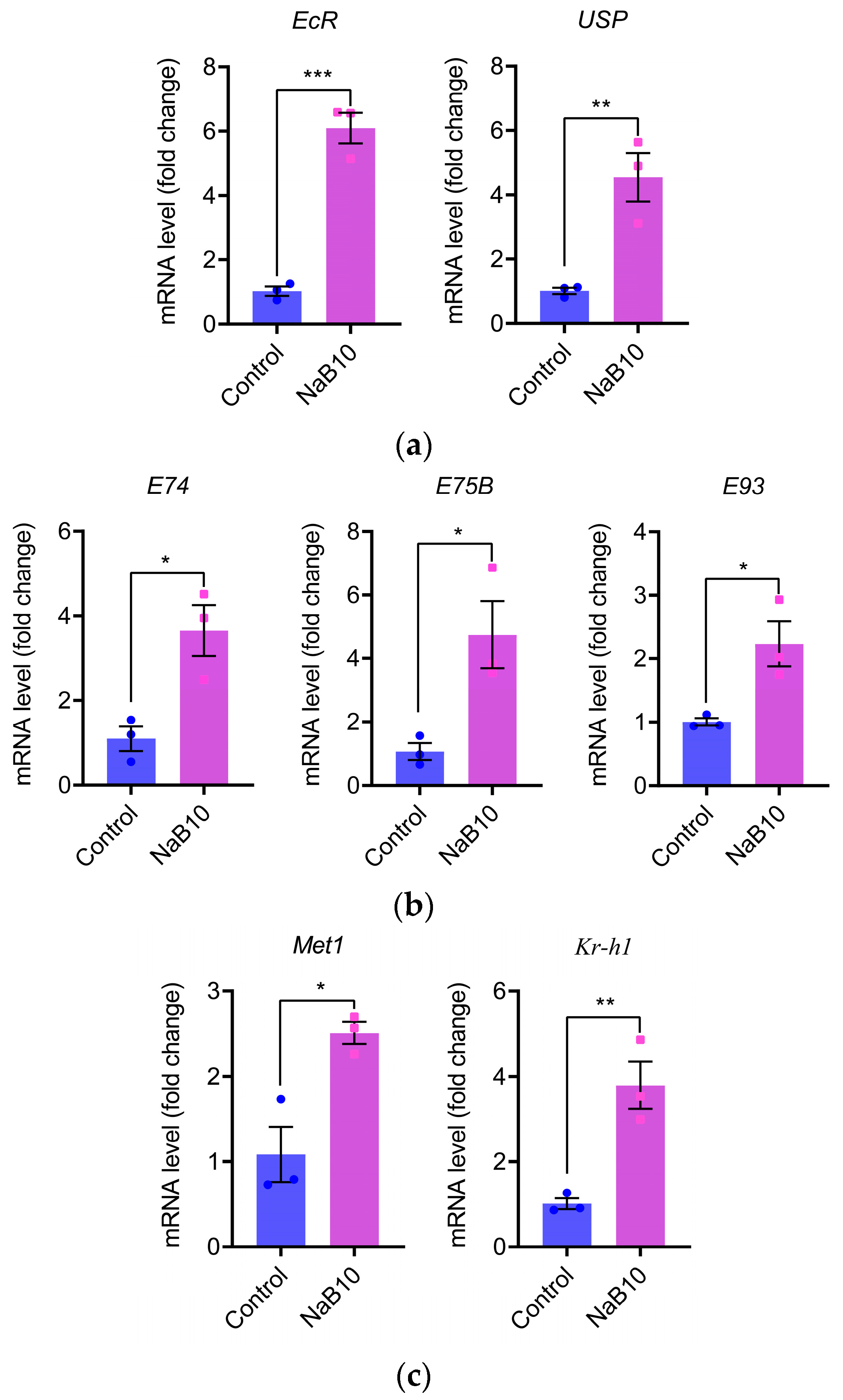
3.2. Effects of NaB on Hormonal Regulatory Pathways in Silkworms
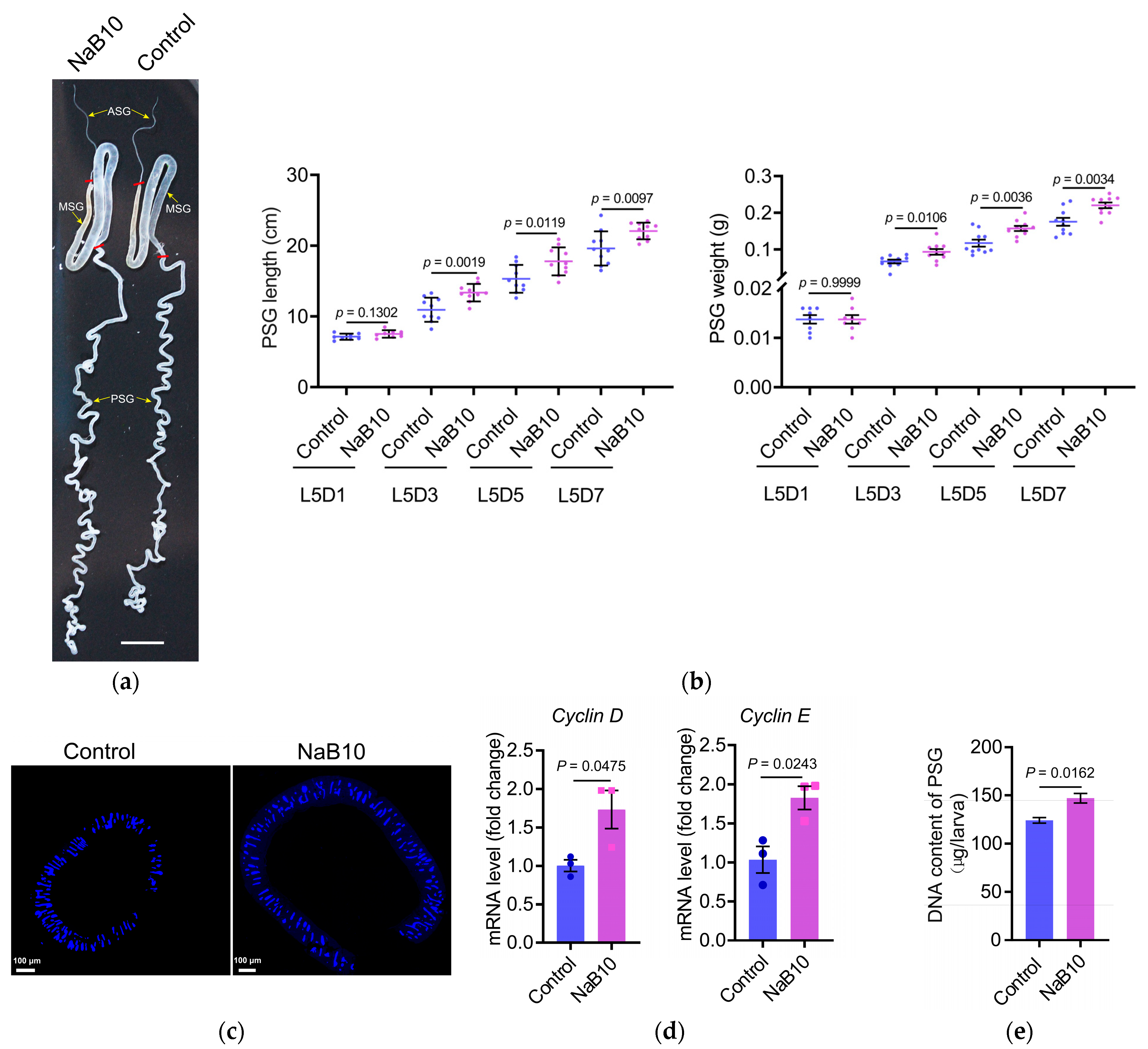
3.3. Effects of Dietary NaB on Silk Gland Development
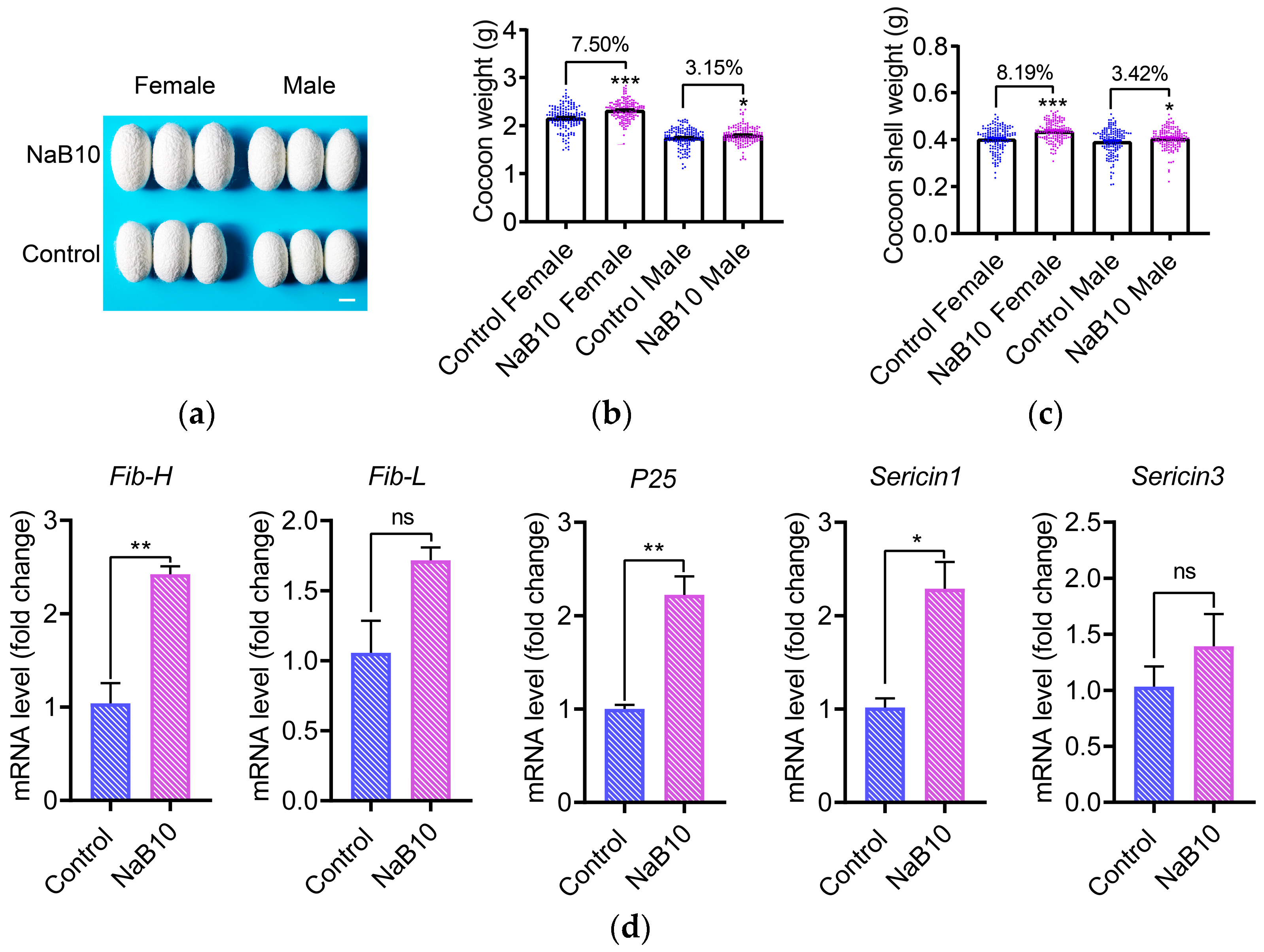
3.4. Effects of Dietary NaB on Economic Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NaB | Sodium butyrate |
| SG | Silk gland |
| MSG | Middle silk gland |
| PSG | Posterior silk gland |
References
- Li, S.; Lao, J.; Sun, Y.; Hua, X.; Lin, P.; Wang, F.; Shen, G.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated editing of BmEcKL1 gene sequence affected silk gland development of silkworms (Bombyx mori). Int. J. Mol. Sci. 2024, 25, 1907. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, X.; Zhao, P.; Sun, Y.; Zhao, X.; Xiong, Y.; Xu, G.; Xia, Q. GC/MS-based metabolomic studies reveal key roles of glycine in regulating silk synthesis in silkworm, Bombyx mori. Insect Biochem. Mol. 2015, 57, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Luo, C.; Pu, Q.; Yin, Q.; Xu, L.; Peng, X.; Ma, S.; Xia, Q.; Liu, S. Let-7 microRNA is a critical regulator in controlling the growth and function of silk gland in the silkworm. RNA. Biol. 2020, 17, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Zuo, W.; Li, C.; Gao, R.; Zhang, H.; Tong, X.; Han, M.; Hu, H.; Lu, C.; Dai, F. Identification of genes that control silk yield by RNA sequencing analysis of silkworm (Bombyx mori) strains of variable silk yield. Int. J. Mol. Sci. 2018, 19, 3718. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, Y.; Zhang, G.; Zhang, Y. Sericin from fibroin-deficient silkworms served as a promising resource for biomedicine. Polymers 2023, 15, 2941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, S.; Song, H.-S.; Zhang, G.; Jia, Q.; Li, S. YorkieCA overexpression in the posterior silk gland improves silk yield in Bombyx mori. J. Insect Physiol. 2017, 100, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, F.; Guo, K.; Xu, J.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated gene editing of the let-7 seed sequence improves silk yield in the silkworm, Bombyx mori. Int. J. Biol. Macromol. 2023, 243, 124793. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhan, L.; Chen, X.; Li, J.; Zhang, M.; Xu, P. Comparative transcriptome analysis of midgut of Bombyx mori under a sucrose-supplemented diet. Sci. Rep. 2025, 15, 15643. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.-J.; Hu, F.; Hong, T.-T.; Thakur, K.; Tang, S.-M.; Zhang, J.-G.; Wei, Z.-J. MiRNA omics reveal the mechanisms of the dual effects of selenium supplementation on the development of the silkworm (Bombyx mori). Int. J. Mol. Sci. 2025, 26, 3394. [Google Scholar] [CrossRef] [PubMed]
- Alfazairy, A.A.; Elsakhawy, D.A.; El-Meniawi, F.A.; Hashem, M.; Rawash, I.A. Effects of mulberry leaf enrichment with Lepidium sativum L. seed powder suspension on the economic parameters of Bombyx mori L. Sci. Rep. 2024, 14, 19600. [Google Scholar] [CrossRef] [PubMed]
- Lattala, G.M.; Kandukuru, K.; Gangupantula, S.; Mamillapalli, A. Spermidine enhances the silk production by mulberry silkworm. J. Insect Sci. 2014, 14, 207. [Google Scholar] [CrossRef] [PubMed]
- Perego, S.; Sansoni, V.; Banfi, G.; Lombardi, G. Sodium butyrate has anti-proliferative, pro-differentiating, and immunomodulatory effects in osteosarcoma cells and counteracts the TNFα-induced low-grade inflammation. Int. J. Immunopathol. Pharmacol. 2018, 31, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, Z.; Shao, K.; Wang, X.; Huang, F.; Qi, J.; Duan, Y.; Jia, Y.; Xu, M. Effects of sodium butyrate supplementation in milk on the growth performance and intestinal microbiota of preweaning holstein calves. Animals 2023, 13, 2069. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yang, Y.; Liu, W.; Bu, D. Sodium butyrate supplementation impacts the gastrointestinal bacteria of dairy calves before weaning. Appl. Microbiol. Biotechnol. 2023, 107, 3291–3304. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Yu, Y.; Wen, X.; Xiao, H.; Liu, K.; Liu, Z.; Liu, S.; Li, Q.; Wang, X.; Deng, Z.; et al. Effects of dietary sodium butyrate on growth performance, immune function, and intestinal microflora of Chinese soft-shelled turtle (Pelodiscus sinensis). Front. Cell. Infect. Microbiol. 2023, 13, 1271912. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.D.; Lindemann, M.D.; Monegue, H.J.; Monegue, J.S. The effect of coated sodium butyrate supplementation in sow and nursery diets on lactation performance and nursery pig growth performance. Life Sci. 2017, 195, 13–20. [Google Scholar] [CrossRef]
- Zeng, T.; Sun, H.; Huang, M.; Guo, R.; Gu, T.; Cao, Y.; Li, C.; Tian, Y.; Chen, L.; Li, G.; et al. Dietary supplementation of coated sodium butyrate improves growth performance of laying ducks by regulating intestinal health and immunological performance. Front. Immunol. 2023, 14, 1142915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, K.; Wang, J.; Bai, S.; Zeng, Q.; Peng, H.; Zhang, B.; Xuan, Y.; Ding, X. Effects of coated sodium butyrate on performance, egg quality, nutrient digestibility, and intestinal health of laying hens. Poult. Sci. 2022, 101, 102020. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ye, A.; Wu, X.; Qu, Z.; Xu, S.; Sima, Y.; Wang, Y.; He, R.; Jin, F.; Zhan, P.; et al. Combined analysis of silk synthesis and hemolymph amino acid metabolism reveal key roles for glycine in increasing silkworm silk yields. Int. J. Biol. Macromol. 2022, 209, 1760–1770. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Fujimoto, Z.; Shiotsuki, T.; Tsuchiya, W.; Momma, M.; Tase, A.; Miyazawa, M.; Yamazaki, T. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm, Bombyx mori. Sci. Rep. 2011, 1, 133. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, S.; Li, C.; Zhou, W.; Dai, F.; Tong, X. BmE2F1 regulates endoreplication of silk gland cells in silkworm, Bombyx mori. Int. J. Biol. Macromol. 2025, 291, 138916. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, J.; Zhao, M.; Liu, M.; Yue, Z.; Liu, L.; Li, F. Effect of sodium butyrate on slaughter performance, serum indexes and intestinal barrier of rabbits. J. Anim. Physiol. Anim. Nutr. 2021, 106, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Tang, S.; Zhao, H.; Zhong, R.; Liu, L.; Meng, Q.; Zhang, H.; Chen, L. Combined effects of sodium butyrate and xylo-oligosaccharide on growth performance, anti-inflammatory and antioxidant capacity, intestinal morphology and microbiota of broilers at early stage. Poultry Sci. 2023, 102, 102585. [Google Scholar] [CrossRef] [PubMed]
- Elnesr, S.S.; Ropy, A.; Abdel-Razik, A.H. Effect of dietary sodium butyrate supplementation on growth, blood biochemistry, haematology and histomorphometry of intestine and immune organs of Japanese quail. Animal 2019, 13, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Li, M.; Li, P.; Chen, B.; Huang, W.; Guo, H.; Cao, J.; Zhao, H. Effects of sodium butyrate on growth performance, antioxidant status, inflammatory response and resistance to hypoxic stress in juvenile largemouth bass (Micropterus salmoides). Front. Immunol. 2023, 14, 1265963. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.D.; Bayir, H.O.; Cosby, D.E.; Cox, N.A.; Williams, S.M.; Fowler, J. Evaluation of encapsulated sodium butyrate on growth performance, energy digestibility, gut development, and Salmonella colonization in broilers. Poultry Sci. 2017, 96, 3638–3644. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.X.; Li, S.Q.; Zhao, Z.; An, L.L. Sodium butyrate as an effective feed additive to improve growth performance and gastrointestinal development in broilers. Vet. Med. Sci. 2020, 6, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.Z.; Everaert, N.; Dunisławska, A. In ovo sodium butyrate administration differentially impacts growth performance, intestinal barrier function, immune response, and gut microbiota characteristics in low and high hatch-weight broilers. J. Anim. Sci. Biotechnol. 2024, 15, 165. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wang, X.; Liu, Y.; Liu, J.; Xiao, S.; Yang, C.; Zhong, Y. Effects of dietary microcapsule sustained-release sodium butyrate on the growth performance, immunity, and gut microbiota of yellow broilers. Animals 2023, 13, 3598. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, Z.; Song, Q.; Jia, Y.; Qi, J.; Xu, M. Modulating gastrointestinal microbiota in preweaning dairy calves: Dose-dependent effects of milk-based sodium butyrate supplementation. Microorganisms 2024, 12, 333. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Ma, X.; Yang, X.; Wang, L.; Jiang, Z. Effect of sodium butyrate on cell proliferation and cell cycle in porcine intestinal epithelial (IPEC-J2) cells. Vitr. Cell. Dev. Biol.—Animal 2017, 53, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sun, D.; Mao, S.; Zhu, W.; Liu, J. Infusion of sodium butyrate promotes rumen papillae growth and enhances expression of genes related to rumen epithelial VFA uptake and metabolism in neonatal twin lambs. J. Anim. Sci. 2019, 97, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Upadhaya, S.D.; Jiao, Y.; Kim, Y.M.; Lee, K.Y.; Kim, I.H. Coated sodium butyrate supplementation to a reduced nutrient diet enhanced the performance and positively impacted villus height and faecal and digesta bacterial composition in weaner pigs. Anim. Feed Sci. Tech. 2020, 265, 114534. [Google Scholar] [CrossRef]
- Dahiya, R.; Berwal, R.S.; Sihag, S.; Patil, C.S.; Lalit. The effect of dietary supplementation of salts of organic acid on production performance of laying hens. Vet. World 2016, 9, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, G.; Wang, H.; Mo, W.; Huang, Y.; Cao, J.; Li, P. Effects of dietary sodium butyrate on growth, digestive enzymes, body composition and nutrient retention-related gene expression of juvenile yellow catfish (Pelteobagrus fulvidraco). Anim. Nutr. 2021, 7, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Smidt, H.; Wu, W.; Xiao, Z.; An, W.; Dong, Y.; Zhang, B. Dietary sodium butyrate improves intestinal development and function by modulating the microbial community in broilers. PLoS ONE 2018, 13, e0197762. [Google Scholar]
- Layalle, S.; Arquier, N.; Léopold, P. The TOR pathway couples nutrition and developmental timing in Drosophila. Dev. Cell 2008, 15, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.; Li, Y.; Xu, G.; Deng, H.; King-Jones, K.; Li, S. Nutrient status alters developmental fates via a switch in mitochondrial homeodynamics. Nat. Commun. 2025, 16, 1258. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.T.; Tang, C.K.; Wu, C.P.; Wu, P.C.; Yang, E.C.; Tai, C.C.; Wu, Y.L. Histone deacetylase inhibitor treatment restores memory-related gene expression and learning ability in neonicotinoid-treated Apis mellifera. Insect Mol. Biol. 2018, 27, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Kmetzsch, L. Histone deacetylases: Targets for antifungal drug development. Virulence 2015, 6, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Tong, S.-M.; Shao, W.; Ying, S.-H.; Feng, M.-G. Pleiotropic effects of the histone deacetylase Hos2 linked to H4-K16 deacetylation, H3-K56 acetylation, and H2A-S129 phosphorylation in Beauveria bassiana. Cell. Microbiol. 2018, 20, e12839. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Gaddelapati, S.C.; Palli, S.R. Histone deacetylase 1 suppresses Krüppel homolog 1 gene expression and influences juvenile hormone action in Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2019, 116, 17759–17764. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Palli, S.R. Epigenetic modifications acetylation and deacetylation play important roles in juvenile hormone action. BMC Genom. 2018, 19, 934. [Google Scholar] [CrossRef] [PubMed]
- Gaddelapati, S.C.; Palli, S.R. Histone deacetylases synergistically regulate juvenile hormone signaling in the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2025, 177, 104256. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Xu, Z.; Li, W.; Cai, C.; Wang, W.; Ge, P.; Jia, X.; Li, Y.; Ding, T.; Ma, W.; et al. HDAC3 knockdown dysregulates juvenile hormone and apoptosis-related nenes in Helicoverpa armigera. Int. J. Mol. Sci. 2022, 23, 14820. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Li, H.; Zhang, X.; Tang, Y.; Yuan, D.; Huang, Z.; Cheng, D. Fzr regulates silk gland growth by promoting endoreplication and protein synthesis in the silkworm. PLoS Genet. 2023, 19, e1010602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bu, L.; Liu, Y.; Huo, W.; Xia, C.; Pei, C.; Liu, Q. Dietary supplementation of sodium butyrate enhances lactation performance by promoting nutrient digestion and mammary gland development in dairy cows. Anim. Nutr. 2023, 15, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Zhang, L.; Zhang, B.; Kong, L.; Pan, X.; Goossens, T.; Song, Z. Dietary sodium butyrate improves female broiler breeder performance and offspring immune function by enhancing maternal intestinal barrier and microbiota. Poult. Sci. 2023, 102, 102658. [Google Scholar] [CrossRef] [PubMed]
- Alhaj, H.W.; Li, Z.; Shan, T.; Dai, P.; Zhu, P.; Li, Y.; Alsiddig, M.A.; Abdelghani, E.; Li, C. Effects of dietary sodium butyrate on reproduction in adult breeder roosters. Anim. Reprod. Sci. 2018, 196, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, T.; Tomita, S.; Uchino, K.; Kimoto, M.; Takiya, S.; Kajiwara, H.; Yamazaki, T.; Sezutsu, H. A Hox gene, Antennapedia, regulates expression of multiple major silk protein genes in the silkworm Bombyx mori. J. Biol. Chem. 2016, 291, 7087–7096. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; He, X.; Ye, Z.; Qing, Z.; Yang, W.; Luo, C.; Xing, D. Dietary Sodium Butyrate Supplementation Enhances Silkworm Silk Yield by Simultaneously Promoting Larval Growth and Silk Gland Development. Insects 2025, 16, 761. https://doi.org/10.3390/insects16080761
Ren X, He X, Ye Z, Qing Z, Yang W, Luo C, Xing D. Dietary Sodium Butyrate Supplementation Enhances Silkworm Silk Yield by Simultaneously Promoting Larval Growth and Silk Gland Development. Insects. 2025; 16(8):761. https://doi.org/10.3390/insects16080761
Chicago/Turabian StyleRen, Xiaoxiao, Xingjiang He, Zhanfeng Ye, Zhuo Qing, Wanjun Yang, Chaobin Luo, and Dan Xing. 2025. "Dietary Sodium Butyrate Supplementation Enhances Silkworm Silk Yield by Simultaneously Promoting Larval Growth and Silk Gland Development" Insects 16, no. 8: 761. https://doi.org/10.3390/insects16080761
APA StyleRen, X., He, X., Ye, Z., Qing, Z., Yang, W., Luo, C., & Xing, D. (2025). Dietary Sodium Butyrate Supplementation Enhances Silkworm Silk Yield by Simultaneously Promoting Larval Growth and Silk Gland Development. Insects, 16(8), 761. https://doi.org/10.3390/insects16080761