
A Non-Destructive Method, Micro-CT, Supports the Identification of Three New Casmara Species from Sumatra and Taiwan (Lepidoptera: Ashinagidae) †




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
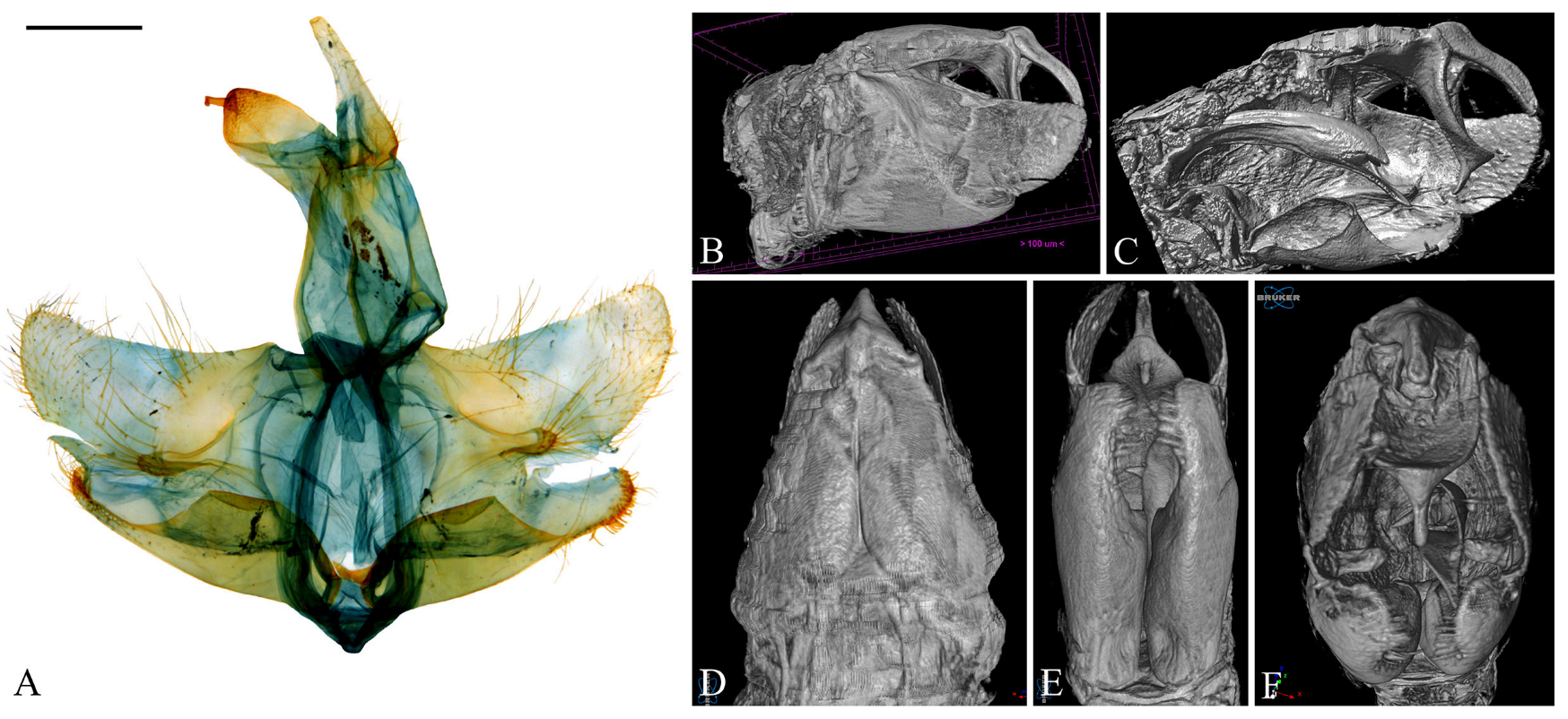
2.1. Specimen Preparation
2.2. Specimen Scanning
2.3. Genitalia Dissection
2.4. 3D Reconstruction of Genitalia
3. Results
- Order Lepidoptera Linnaeus, 1758
- Family Ashinagidae Matsumura, 1929
- Genus Casmara Walker, 1863 [23]
- Type species: Casmara infaustella Walker, 1863
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- May, R.M. Biological diversity: How many species are there? Nature 1986, 324, 514–515. [Google Scholar] [CrossRef] [PubMed]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. Bioscience 2006, 54, 311–323. [Google Scholar] [CrossRef]
- Rasoarimalala, N.F.; Ramiadantsoa, T.; Rakotonirina, J.C.; Fisher, B.L. Linear Morphometry of Male Genitalia Distinguishes the Ant Genera Monomorium and Syllophopsis (Hymenoptera: Formicidae) in Madagascar. Insects 2024, 15, 605. [Google Scholar] [CrossRef] [PubMed]
- Deans, A.R.; Mikó, I.; Wipfler, B.; Friedrich, F. Evolutionary phenomics and the emerging enlightenment of arthropod systematics. Invertebr. Syst. 2012, 26, 323–330. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Kitching, I.J. Virtual dissections through micro-CT scanning: A method for non-destructive genitalia ‘dissections’ of valuable Lepidoptera material. Syst. Entomol. 2014, 39, 606–618. [Google Scholar] [CrossRef]
- Sartori, M.; Kubiak, M.; Michalik, P. Deciphering genital anatomy of rare, delicate and precious specimens: First study of two type specimens of mayflies using micro-computed X-ray tomography (Ephemeroptera; Heptageniidae). Zoosymposia 2016, 11, 28–32. [Google Scholar] [CrossRef]
- Elliott, J.C.; Dover, S.D. X-ray microtomography. J. Microsc. 1982, 126, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Ritman, E.L. Micro-computed tomography—Current status and developments. Annu. Rev. Biomed. Eng. 2004, 6, 185–208. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Saksena, M.; Coopey, S.B.; Fernandez, L.; Buckley, J.M.; Lei, L.; Aftreth, O.; Koerner, F.; Michaelson, J.; Rafferty, E.; et al. Intraoperative micro-computed tomography (micro-CT): A novel method for determination of primary tumour dimensions in breast cancer specimens. Br. J. Radiol. 2016, 89, 20150581. [Google Scholar] [CrossRef] [PubMed]
- Rosin, P.L.; Lai, Y.K.; Liu, C.; Davis, G.R.; Mills, D.; Tuson, G.; Russell, Y. Virtual recovery of content from X-ray micro-tomography scans of damaged historic scrolls. Sci. Rep. 2018, 8, 11901. [Google Scholar] [CrossRef] [PubMed]
- Edie, S.M.; Collins, K.S.; Jablonski, D. High-throughput micro-CT scanning and deep learning segmentation workflow for analyses of shelly invertebrates and their fossils: Examples from marine Bivalvia. Front. Ecol. Evol. 2023, 11, 1127756. [Google Scholar] [CrossRef]
- Kim, S.; Kaila, L.; Lee, S. Evolution of larval life mode of Oecophoridae (Lepidoptera: Gelechioidea) inferred from molecular phylogeny. Mol. Phylogenet. Evol. 2016, 101, 314–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Li, H.H. Phylogeny of the superfamily Gelechioidea (Lepidoptera: Obtectomera), with an exploratory application on geometric morphometrics. Zool. Scr. 2020, 49, 307–328. [Google Scholar] [CrossRef]
- Lvovsky, A.L. New and little known species of the genus Casmara Walker, 1863 (Lepidoptera: Oecophoridae) from Southeast Asia. Zoosyst. Ross. 2013, 22, 107–110. [Google Scholar] [CrossRef]
- Wineriter-Wright, S.A.; Smith, M.C.; Metz, M.A.; Makinson, J.R.; Brown, B.T.; Purcell, M.F.; Barr, K.L.; Pratt, P.D. The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate. Insects 2020, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, L.; Wang, J. Four new species of the genus Casmara Walker, 1863 (Lepidoptera: Oecophoridae) from China. Zootaxa 2012, 3239, 58–63. [Google Scholar] [CrossRef]
- Meyrick, E. Second contribution to our knowledge about the Pyralidae and Microlepidoptera of Kwansien. Bull. Sect. Sci. Acad. Roumaine 1931, 14, 59–75. [Google Scholar]
- Meyrick, E. Descriptions of Indian micro-lepidoptera. J. Bombay Nat. Hist. Soc. 1907, 17, 732. [Google Scholar]
- Bradley, J.D. A new species of Casmara Walker, 1863 from Malaya. Ann. Mag. Nat. His. 1953, 12, 319–320. [Google Scholar] [CrossRef]
- Meyrick, E. Exotic Microlepidoptera 2; Marlborough: Bolton, UK, 1922; Volume 17, p. 544. [Google Scholar]
- Meyrick, E. Exotic Microlepidoptera 1; Marlborough: Bolton, UK, 1914; Volume 8, p. 237. [Google Scholar]
- Moriuti, S. A new and an unrecorded species of Casmara Walker (Lepidoptera: Oecophoridae) from Thailand. Tinea 1985, 12, 11–16. [Google Scholar]
- Walker, F. List of the Specimens of Lepidopterous Insects in the Collection of the British Museum; British Museum: London, UK, 1863; Volume 28, p. 518. [Google Scholar]
- Diakonoff, A. Records and descriptions of South Asiatic Microlepidoptera. Tijdschr. Entomol. 1966, 109, 49–88. [Google Scholar]
- Meyrick, E. Materialien zu einer Microlepidopteren-fauna Kwangtungs, Deutsche entomologische. Zeitschrift Iris 1934, 48, 38. [Google Scholar]
- Heppner, J.B. Classification of Lepidoptera: Part 1: Introduction; Holarctic Lepidoptera; Association for Tropical Lepidoptera: Statesboro, GA, USA, 1998; pp. 28–31. [Google Scholar]
- Latief, F.D.E.; Sari, D.S.; Fitri, L.A. Applications of Micro-CT scanning in medicine and dentistry: Microstructural analyses of a Wistar Rat mandible and a urinary tract stone. J. Phys. Conf. Ser. 2017, 884, 012042. [Google Scholar] [CrossRef]
- Han, S.C.; Chung, Y.E.; Lee, Y.H.; Park, K.K.; Kim, M.J.; Kim, K.W. Metal artifact reduction software used with abdominopelvic dual-energy CT of patients with metal hip prostheses: Assessment of image quality and clinical feasibility. AJR Am. J. Roentgenol. 2014, 203, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, W.; Lee, S. Estimation of a new molecular marker of the genus Stathmopoda (Lepidoptera: Stathmopodidae): Comparing EF1a and COI sequences. J. Asia Pac. Entomol. 2017, 20, 269–280. [Google Scholar] [CrossRef]
- Lima, M.G.; Jussiani, E.I.; Andrello, A.C.; Zequi, J.A.C.; Kawabata, E.K. Potentialities of iodine-enhanced micro-CT imaging in the morphological study of adult Aedes (Stegomyia) aegypti (Linnaeus, 1762) mosquitoes. Micron 2023, 103518. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species of Casmara | Distribution | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| KOR | JPN | CHN | TWN | THA | HKG | VIE | MAS | INA | IND | AUS | |
| C. acantha Wang [16] Zootaxa 3239: 58–63. TL: China. | ● | ● | |||||||||
| C. aduncata Wang [16] Zootaxa 3239: 58–63. TL: China. | ● | ||||||||||
| C. agronoma Meyrick [17] Bulletin de la Section Scientifique de l’Academie Roumaine 14: 59–75. TL: China. | ● | ● | ● | ● | |||||||
| C. demotarcha (Meyrick) [18] Journal of the Bombay Natural History Society 17(3): 742. TL: Khasi Hills. | ● | ||||||||||
| C. diabolella Bradley [19] Annals and magazine of natural history 12(6): 319–320. TL: Malaysia. | ● | ||||||||||
| C. epicompsa Meyrick [20] Exotic Microlepidoptera 2(17): 544. TL: Bengal, Darjiling. | ● | ||||||||||
| C. exculta (Meyrick) [21] Exotic Microlepidoptera 1(8): 237. TL: Assam, Khasis. | ● | ● | ● | ● | ● | ● | |||||
| C. falcatussica sp. nov. * TL: New Taipei City. | ● | ||||||||||
| C. fulvacorona sp. nov. * TL: Sumatra. | ● | ||||||||||
| C. fuscatulipa sp. nov. * TL: Taoyuan City. | ● | ||||||||||
| C. grandipennata Moriuti [22] Tinea 12(2): 11–16. TL: Thailand. | ● | ||||||||||
| C. infaustella Walker [23] List of the specimens of lepidopterous insects in the collection of the British Museum 28: 518. TL: North Hindostan. | ● | ||||||||||
| C. kalshoveni Diakonoff [24] Tijdschrift voor Entomologie 109(3): 69. TL: Central Java. | ● | ||||||||||
| C. longiclavata Wang [16] Zootaxa 3239: 58–63. TL: China. | ● | ● | ● | ● | |||||||
| C. nedoshivinae Lvovsky [14] Zoosystematic Rossica 22(1): 108. TL: Thua Thien Hue. | ● | ● | |||||||||
| C. patrona Meyrick [25] Deutsche entomologische Zeitschrift Iris 48: 38. TL: China. | ● | ● | ● | ||||||||
| C. phobographa Diakonoff [24] Tijdschrift voor Entomologie 109(3): 62. TL: Sumatra. | ● | ||||||||||
| C. quadrilatera Wang [16] Zootaxa 3239: 58–63. TL: China. | ● | ||||||||||
| C. regalis Diakonoff [24] Tijdschrift voor Entomologie 109(3): 61. TL: Celebes, Pangean nr Maros. | ● | ● | |||||||||
| C. rhodotrachys Diakonoff [24] Tijdschrift voor Entomologie 109(3): 67. TL: Borneo, Balikpapan. | ● | ||||||||||
| C. rodochalca Meyrick [25] Deutsche entomologische Zeitschrift Iris 48: 38. TL: China. | ● | ||||||||||
| C. rufipes Diakonoff [24] Tijdschrift voor Entomologie 109(3): 65. TL: Java. | ● | ||||||||||
| C. subagronoma Lvovsky [14] Zoosystematic Rossica 22(1): 108. TL: Vinh Phuc, Ngoc Thanh Vill. | ● | ● | ● | ||||||||
| C. uniata Diakonoff [24] Tijdschrift voor Entomologie 109(3): 64. TL: Celebes, Paloe District. | ● | ||||||||||
| System Settings | Acquisition Settings |
|---|---|
| Software Version = 1.6 | Partial Width = 75% |
| Magnification Drive Version = 3.4 | Image Rotation = 0.03000 |
| Camera Drive Version = 3.4 | Source Voltage (kV) = 40 |
| Filter Drive Version = 3.4 | Source Current (μA) = 100 |
| Animal Drive Version = 3.4 | Image Pixel Size (μm) = 4.0 |
| Source Type = HAMAMATSU L10321-67 | Exposure (ms) = 1200 |
| Filter = No Filter | |
| Camera Type = XIMCA MH110XC-KK-TP | Camera Binning = 1 × 1 |
| Frame Averaging = ON (2) | |
| Camera Pixel Size (μm) = 17.250 | Scanning Trajectory = ROUND |
| Camera X/Y Ration = 1.0032 | FF Updating Interval = 172 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, I.-W.; Kim, S.; Heppner, J.B. A Non-Destructive Method, Micro-CT, Supports the Identification of Three New Casmara Species from Sumatra and Taiwan (Lepidoptera: Ashinagidae). Insects 2025, 16, 747. https://doi.org/10.3390/insects16080747
Jeong I-W, Kim S, Heppner JB. A Non-Destructive Method, Micro-CT, Supports the Identification of Three New Casmara Species from Sumatra and Taiwan (Lepidoptera: Ashinagidae). Insects. 2025; 16(8):747. https://doi.org/10.3390/insects16080747
Chicago/Turabian StyleJeong, In-Won, Sora Kim, and John B. Heppner. 2025. "A Non-Destructive Method, Micro-CT, Supports the Identification of Three New Casmara Species from Sumatra and Taiwan (Lepidoptera: Ashinagidae)" Insects 16, no. 8: 747. https://doi.org/10.3390/insects16080747
APA StyleJeong, I.-W., Kim, S., & Heppner, J. B. (2025). A Non-Destructive Method, Micro-CT, Supports the Identification of Three New Casmara Species from Sumatra and Taiwan (Lepidoptera: Ashinagidae). Insects, 16(8), 747. https://doi.org/10.3390/insects16080747