
Function of Vivid Coloration of Terrestrial Isopods from the Point of View of an Avian Predator



Simple Summary
Abstract

1. Introduction
- All isopod species are protected against attack by birds better than edible roach.
- Isopod species with conspicuous coloration are better protected against attack by birds than inconspicuous species.
- Birds do not show any disgust after eating isopods.
- Birds prefer attacking roaches rather than isopods when encountering both simultaneously.
2. Materials and Methods
2.1. Prey
- The Common Rough Woodlouse Porcellio scaber Latreille, 1804 (coded as scaber throughout the text): Body size up to 17 mm, nearly twice as long as wide. The body is strongly irregularly tuberculate. Variable in color. Usually, dark slate grey, sometimes brightly mottled cream, brown, orange, or red forms are found [66]. Distributed across Central and Western Europe but introduced to most countries [67]. Common in the Czech Republic.
- The Common Shiny Woodlouse, Oniscus asellus Linnaeus, 1758 (coded as asellus throughout the text): Body size up to 18 mm, about twice as long as wide. Usually, shiny slate grey with irregular lighter patterns and two rows of yellow patches [66]. Distributed in Northern and Western Europe, east to Finland, Poland, and Ukraine. Also found in Madeira and the Azores. Introduced to America [68]. Common in the Czech Republic.
- Armadillidium versicolor Stein, 1859 (coded as versicolor throughout the text): Body size 10 mm long. Dark coloring varies from black-brown to olive-brown, with light spots forming 5 regular rows on the dorsal side and pale-yellow margins of epimeres [70]. Found in Eastern Europe [68]. Rare in the Czech Republic, but locally abundant.
- Armadillidium gestroi Tua, 1900 (coded as gestroi throughout the text): Body size up to 20 mm. Color dark or light brown with vivid yellow spots forming 4 regular rows. Found in northwest Italy [68]. Not occurring in the Czech Republic.
2.2. Predator
2.3. Experiment
2.4. Data Analyses
3. Results
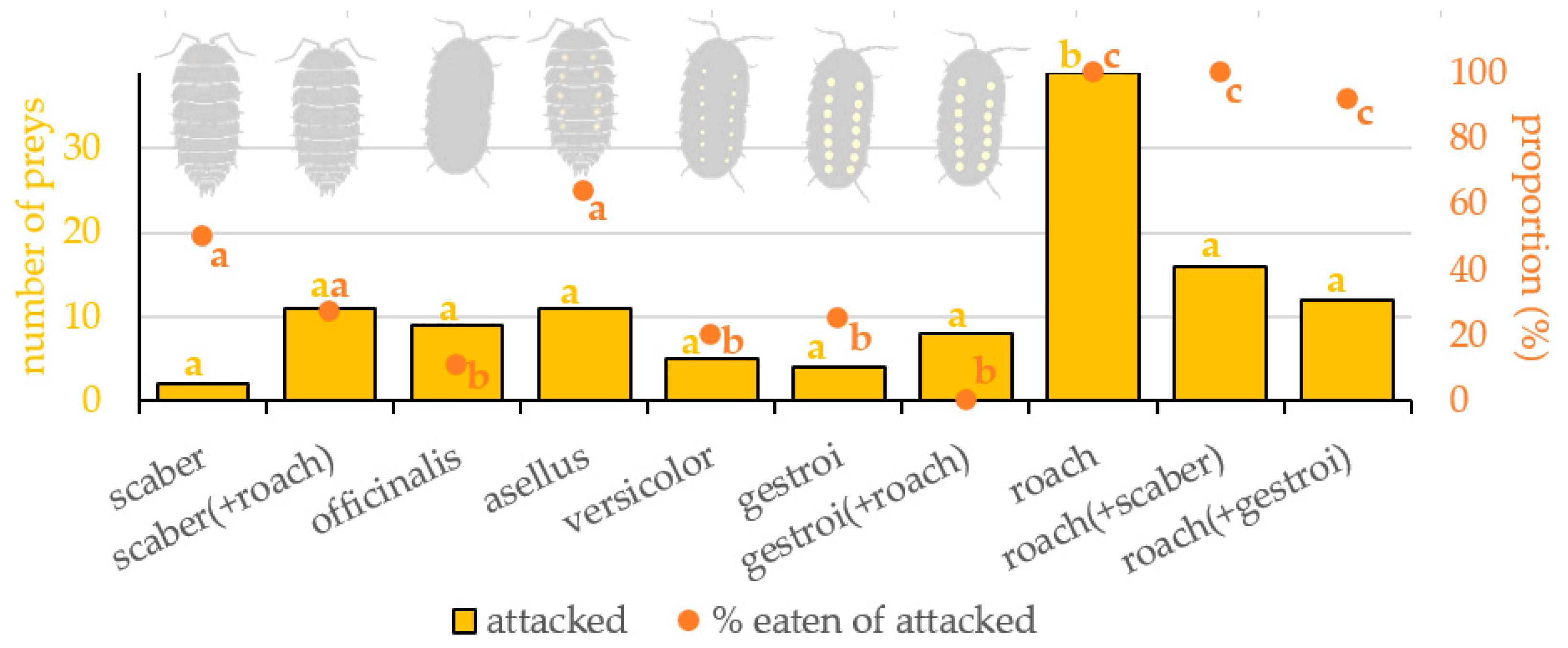
3.1. Attacking of Prey
3.2. Eating of Prey
3.3. Observation of Prey
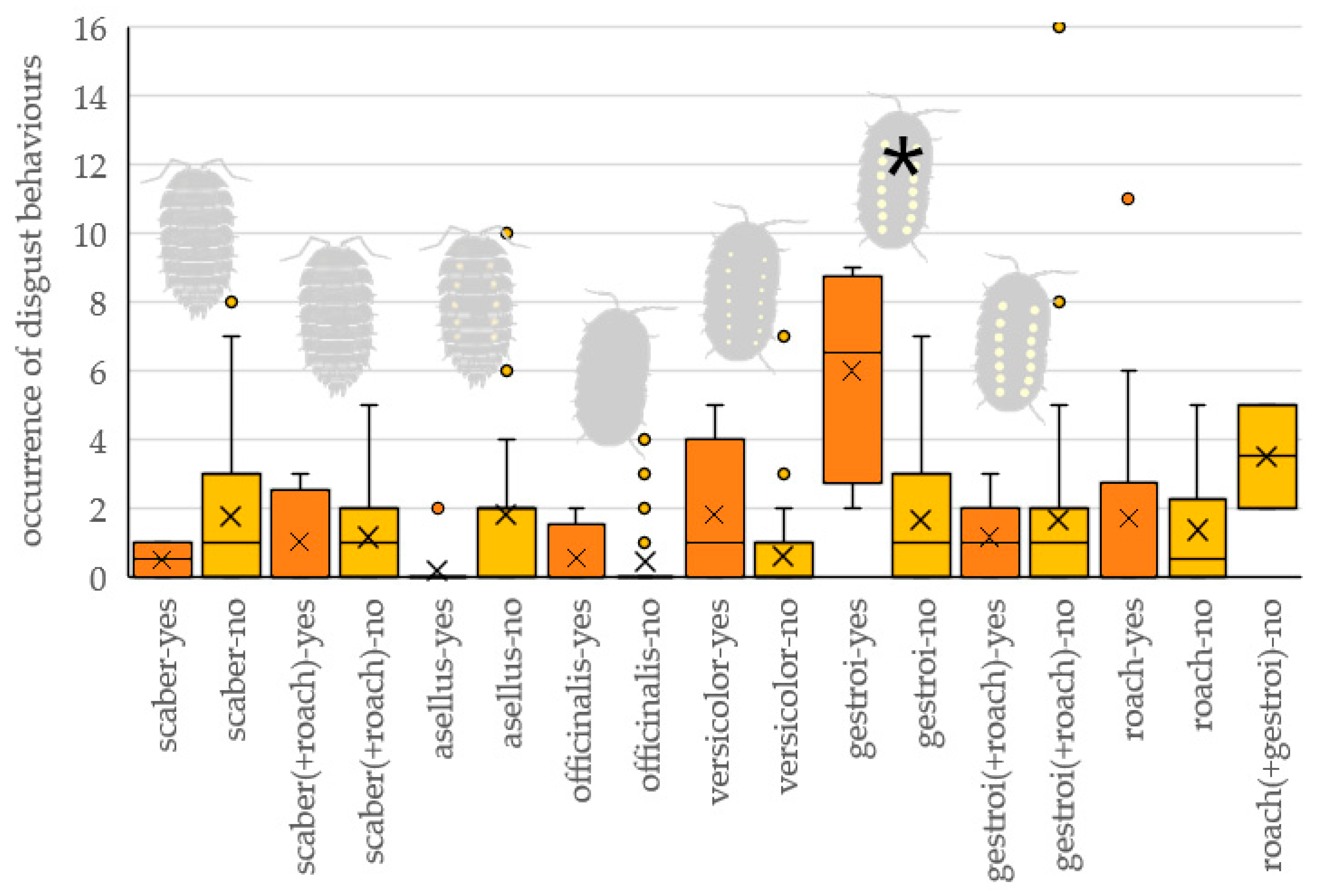
3.4. Signs of Discomfort
4. Discussion
4.1. Attacking and Eating Prey
4.2. Observing Prey
4.3. Discomfort Behavior
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Eisner, T.; Alsop, D.; Hicks, K.; Meinwald, J. Defensive secretion of millipedes. In Arthropod Venoms, Handbook of Experimental Pharmacology/Handbuch der Experimentellen Pharmakologie; Bettini, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; Volume 48, pp. 41–72. [Google Scholar] [CrossRef]
- Roth, L.M.; Eisner, T. Chemical defenses of arthropods. Annu. Rev. Entomol. 1962, 7, 107–136. [Google Scholar] [CrossRef]
- Minelli, A. Secretions of centipedes. In Arthropod Venoms, Handbook of Experimental Pharmacology/Handbuch der Experimentellen Pharmakologie; Bettini, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; Volume 48, pp. 73–85. [Google Scholar] [CrossRef]
- Pekár, S.; Raspotnig, G. Defences of Arachnids: Diversified arsenal used against range of enemies. Entomol. Gen. 2022, 42, 663–679. [Google Scholar] [CrossRef]
- Schildknecht, H. The defensive chemistry of land and water beetles. Angew. Chem. Int. Ed. 1970, 9, 1–9. [Google Scholar] [CrossRef]
- Talbot, P.; Demers, D. Tegumental glands of Crustacea. In The Crustacean Integument: Morphology and Biochemistry; Horst, M.N., Freeman, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 151–191. [Google Scholar]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef]
- Sugiura, S. Anti-predator defences of a bombardier beetle: Is bombing essential for successful escape from frogs? PeerJ 2018, 6, e5942. [Google Scholar] [CrossRef]
- Leimar, O.; Enquist, M.; Sillen-Tullberg, B. Evolutionary stability of aposematic coloration and prey unprofitability: A theoretical analysis. Am. Nat. 1986, 128, 469–490. [Google Scholar] [CrossRef]
- Ruxton, G.; Sherratt, T.; Speed, M. Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry; Oxford University Press: Oxford, UK, 2004. [Google Scholar] [CrossRef]
- Pekár, S.; Garcia, L.F.; Bulbert, M.W. Spiders mimic the acoustic signalling of mutillid wasps to avoid predation: Startle signalling or Batesian mimicry? Anim. Behav. 2020, 170, 157–166. [Google Scholar] [CrossRef]
- Gamberale-Stille, G.; Guilford, T. Automimicry destabilizes aposematism: Predator sample-and-reject behaviour may provide a solution. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, 2621–2625. [Google Scholar] [CrossRef]
- Guilford, T. The evolution of aposematism. In Insect Defense: Adaptive Mechanisms and Strategies of Prey and Predators; Evans, D.I., Schmidt, J.O., Eds.; State University of New York Press: New York, NY, USA, 1990; pp. 23–61. [Google Scholar]
- Prudic, K.L.; Skemp, A.K.; Papaj, D.R. Aposematic coloration, luminance contrast, and the benefits of conspicuousness. Behav. Ecol. 2007, 18, 41–46. [Google Scholar] [CrossRef]
- Wyszecki, G.; Stiles, W.S. Color Science: Concepts and Methods, Quantitative Data and Formulae; John Wiley & Sons, Inc.: New York, NY, USA, 1982. [Google Scholar]
- Adelson, E.H. Lightness perception and lightness illusions. In The New Cognitive Neurosciences; Gazzaniga, M., Ed.; MIT Press: Cambridge, MA, USA, 2000; pp. 339–351. [Google Scholar]
- Goldsmith, T.H. Optimization, constraint, and history in the evolution of eyes. Q. Rev. Biol. 1990, 65, 281–322. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection; Dover Publication: New York, NY, USA, 1930. [Google Scholar] [CrossRef]
- Sherratt, T.N.; Beatty, C.D. The evolution of warning signals as reliable indicators of prey defense. Am. Nat. 2003, 162, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Skelhorn, J.; Halpin, C.G.; Rowe, C. Learning about aposematic prey. Behav. Ecol. 2016, 27, 955–964. [Google Scholar] [CrossRef]
- Smith, K.E.; Halpin, C.G.; Rowe, C. Body size matters for aposematic prey during predator aversion learning. Behav. Process. 2014, 109, 173–179. [Google Scholar] [CrossRef]
- Roberts, S.M.; Stuart-Fox, D.; Medina, I. The evolution of conspicuousness in frogs: When to signal toxicity? J. Evol. Biol. 2022, 35, 1455–1464. [Google Scholar] [CrossRef]
- White, T.E.; Umbers, K.D. Meta-analytic evidence for quantitative honesty in aposematic signals. Proc. R. Soc. B Biol. Sci. 2021, 288, 20210679. [Google Scholar] [CrossRef] [PubMed]
- Rojas, B.; Luis-Martínez, A.; Mappes, J. Frequency-dependent flight activity in the colour polymorphic wood tiger moth. Curr. Zool. 2015, 61, 765–772. [Google Scholar] [CrossRef]
- Marek, P.E.; Bond, J.E. A Müllerian mimicry ring in Appalachian millipedes. Proc. Natl. Acad. Sci. USA 2009, 106, 9755–9760. [Google Scholar] [CrossRef]
- Gorvett, H. The tegumental glands in the land Isopoda: A. The rosette glands. J. Cell Sci. 1946, 2, 209–235. [Google Scholar] [CrossRef]
- Gorvett, H. The Tegumental glands in the land Isopoda: B. The lobed glands: Structure and distribution. J. Cell Sci. 1951, 3, 275–296. [Google Scholar] [CrossRef]
- Gorvett, H. The tegumental glands in the land Isopoda: C. The lobed glands: The properties of their secretion and their mode of action. J. Cell Sci. 1952, 3, 17–29. [Google Scholar] [CrossRef]
- Gorvett, H. Tegumental glands and terrestrial life in woodlice. Proc. Zool. Soc. Lond. 1956, 126, 291–314. [Google Scholar] [CrossRef]
- Herold, W. Beiträge zur Anatomie und Physiologie einiger Landisopoden. Zool. Jahrbücher Abt. Anat. Ontog. Tiere 1913, 35, 455–526. [Google Scholar]
- Paris, O.H. The ecology of Armadillidium vulgare (Isopoda: Oniscoidea) in California grassland: Food, enemies, and weather. Ecol. Monogr. 1963, 33, 1–22. [Google Scholar] [CrossRef]
- Deslippe, R.; Jelinski, L.; Eisner, T. Defense by use of a proteinaceous glue: Woodlice vs. ants. Zool. Anal. Complex Syst. 1996, 99, 205–210. [Google Scholar]
- Brereton, J.L.G. The distribution of woodland isopods. Oikos 1957, 8, 85–106. [Google Scholar] [CrossRef]
- Crowcroft, P. The Life of the Shrew; M. Reinhardt: London, UK, 1957. [Google Scholar]
- Schmalfuss, H.; Ferrara, F. Observations on the distribution and ecology of terrestrial isopods (Oniscoidea) in South-west Cameroon. Monit. Zool. Ital. 1982, 17, 243–265. [Google Scholar] [CrossRef]
- Levi, H. An unusual case of mimicry. Evolution 1965, 19, 26–262. [Google Scholar] [CrossRef]
- Schmalfuss, H. Revision of the Armadillidium klugii-group (Isopoda: Oniscidea). Stuttg. Beiträge Naturkunde A 2013, 6, 1–12. [Google Scholar]
- Hart, N.S. The visual ecology of avian photoreceptors. Prog. Retin. Eye Res. 2001, 20, 675–703. [Google Scholar] [CrossRef]
- Walls, G.L. The Vertebrate Eye and Its Adaptive Radiation; Cranbrook Institute of Science: Bloomfield Hills, MI, USA, 1942. [Google Scholar]
- Meyer, D.B. The avian eye and its adaptations. In Handbook of Sensory Physiology, The Visual System in Vertebrates; Crescitelli, F., Ed.; Springer: Berlin/Heidelberg, Germany, 1977; Volume VII. [Google Scholar] [CrossRef]
- Hodos, W. The visual capabilities of birds. In Avian Vision, Brain and Behavior; Ziegler, H.P., Bischof, H.J., Eds.; MIT Press: Cambridge, MA, USA, 1993; pp. 63–76. [Google Scholar]
- Jones, M.P.; Pierce, K.E., Jr.; Ward, D. Avian vision: A review of form and function with special consideration to birds of prey. J. Exot. Pet Med. 2007, 16, 69–87. [Google Scholar] [CrossRef]
- Hunt, D.M.; Carvalho, L.S.; Cowing, J.A.; Davies, W.L. Evolution and spectral tuning of visual pigments in birds and mammals. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2941–2955. [Google Scholar] [CrossRef] [PubMed]
- Lind, O.; Mitkus, M.; Olsson, P.; Kelber, A. Ultraviolet vision in birds: The importance of transparent eye media. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132209. [Google Scholar] [CrossRef]
- Kelber, A. Bird colour vision-from cones to perception. Curr. Opin. Behav. Sci. 2019, 30, 34–40. [Google Scholar] [CrossRef]
- Roper, T.J. Responses of domestic chicks to articially coloured insect prey: Effects of previous experience and background colour. Anim. Behav. 1990, 39, 466–473. [Google Scholar] [CrossRef]
- Roper, T.J.; Cook, S.E. Responses of chicks to brightly coloured insect prey. Behaviour 1989, 110, 276–293. [Google Scholar] [CrossRef]
- Gamberale-Stille, G.; Tullberg, B. Experienced chicks show biased avoidance of stronger signals: An experiment with natural colour variation in live aposematic prey. Evol. Ecol. 1999, 13, 579–589. [Google Scholar] [CrossRef]
- Schuler, W.; Hesse, E. On the function of warning coloration: A black and yellow pattern inhibits prey-attack by naive domestic chicks. Behav. Ecol. Sociobiol. 1985, 16, 249–255. [Google Scholar] [CrossRef]
- Osorio, D.; Miklósi, A.; Gonda, Z. Visual ecology and perception of coloration patterns by domestic chicks. Evol. Ecol. 1999, 13, 673–689. [Google Scholar] [CrossRef]
- Wiklund, C.; Järvi, T. Survival of distasteful insects after being attacked by naïve birds: A reappraisal of the theory of aposematic coloration evolving through individual selection. Evolution 1982, 36, 998–1002. [Google Scholar] [CrossRef]
- Sillén-Tullberg, B. Higher survival of an aposematic than of a cryptic form of a distasteful bug. Oecologia 1985, 67, 411–415. [Google Scholar] [CrossRef]
- Hutchison, L.V.; Wenzel, B.M. Olfactory guidance in foraging by procellariiforms. Condor 1980, 82, 314–319. [Google Scholar] [CrossRef]
- Clark, L.; Mason, J.R. Use of nest material as insecticidal and anti-pathogenic agents by the European starling. Oecologia 1985, 67, 169–176. [Google Scholar] [CrossRef]
- Nevitt, G.A.; Veit, R.R.; Kareiva, P. Dimethyl sulphide as a foraging cue for Antarctic procellariiform seabirds. Nature 1995, 376, 680–682. [Google Scholar] [CrossRef]
- Petit, C.; Hossaert-McKey, M.; Perret, P.; Blondel, J.; Lambrechts, M. Blue Tits use selected plants and olfaction to maintain an aromatic environment for nestlings. Ecol. Lett. 2002, 5, 585–589. [Google Scholar] [CrossRef]
- Wallraff, H.G. Avian olfactory navigation: Its empirical foundation and conceptual state. Anim. Behav. 2004, 67, 189–204. [Google Scholar] [CrossRef]
- Roper, T.J. Olfaction in birds. In Advances in the Study of Behavior; Slater, P.J.B., Rosenblatt, J.S., Snowdon, C.T., Roper, T.J., Eds.; Academic Press: Boston, MA, USA, 1999; Volume 28, pp. 247–332. [Google Scholar] [CrossRef]
- Castro, I.; Cunningham, S.J.; Gsell, A.C.; Jaffe, K.; Cabrera, A.; Liendo, C. Olfaction in birds: A closer look at the kiwi (Apterygidae). J. Avian Biol. 2010, 41, 213–218. [Google Scholar] [CrossRef]
- Kare, M. Comparative study of taste. In Handbook of Sensory Physiology 4; Beidler, L.M., Ed.; Springer: New York, NY, USA, 1971; pp. 278–292. [Google Scholar]
- Roura, E.; Baldwin, M.W.; Klasing, K.C. The avian taste system: Potential implications in poultry nutrition. Anim. Feed. Sci. Technol. 2013, 180, 1–9. [Google Scholar] [CrossRef]
- Heyers, D.; Zapka, M.; Hoffmeister, M.; Wild, J.M.; Mouritsen, H. Magnetic field changes activate the trigeminal brainstem complex in a migratory bird. Proc. Natl. Acad. Sci. USA 2010, 107, 9394–9399. [Google Scholar] [CrossRef]
- Van Heezik, Y.M.; Gerritsen, A.F.C.; Swennen, C. The influence of chemoreception on the foraging behaviour of two species of sandpiper, Calidris alba and Calidris alpina. Neth. J. Sea Res. 1983, 17, 47–56. [Google Scholar] [CrossRef]
- Skelhorn, J.; Rowe, C. Taste-rejection by predators and the evolution of unpalatability in prey. Behav. Ecol. Sociobiol. 2006, 60, 550–555. [Google Scholar] [CrossRef]
- Skelhorn, J.; Rowe, C. Tasting the difference: Do multiple defence chemicals interact in Müllerian mimicry? Proc. R. Soc. B Biol. Sci. 2005, 272, 339–345. [Google Scholar] [CrossRef]
- Hopkin, S.P. A key to the woodlice of Britain and Ireland. Field Stud. 1991, 7, 599–650. [Google Scholar]
- Hopkin, S.P.; Jones, D.T.; Dietrich, D. The isopod Porcellio scaber as a monitor of the bioavailability of metals in terrestrial ecosystems: Towards a global ‘woodlouse watch’ scheme. Sci. Total Environ. 1993, 134, 357–365. [Google Scholar] [CrossRef]
- Schmalfuss, H. World catalog of terrestrial isopods (Isopoda: Oniscidea). Stuttg. Beiträge Naturkunde. Ser. A Biol. 2003, 654, 1–341. [Google Scholar]
- Schmalfuss, H. The terrestrial isopod genus Armadillo in western Asia (Oniscidea: Armadillidae), with descriptions of five new species. Stuttg. Beiträge Naturkunde Ser. A 1996, 544, 1–43. [Google Scholar]
- Frankenberger, Z. Stejnonožci suchozemští—Oniscoidea. [Terrestrial Isopods]; Nakladatelství ČSAV: Prague, Czech Republic, 1959. (In Czech) [Google Scholar]
- Karlíková, Z.; Veselý, P.; Beránková, J.; Fuchs, R. Low ability of Great Tits to discriminate similarly inconspicuous edible and inedible prey. Ethology 2016, 122, 180–191. [Google Scholar] [CrossRef]
- Veselý, P.; Fuchs, R. Newly emerged Batesian mimicry protects only unfamiliar prey. Evol. Ecol. 2009, 23, 919–929. [Google Scholar] [CrossRef]
- Veselý, P.; Luhanová, D.; Prášková, M.; Fuchs, R. Generalization of mimics imperfect in colour patterns: The point of view of wild avian predators. Ethology 2013, 119, 138–145. [Google Scholar] [CrossRef]
- Dolenská, M.; Nedvěd, O.; Veselý, P.; Tesařová, M.; Fuchs, R. What constitutes optical warning signals of ladybirds (Coleoptera: Coccinellidae) towards bird predators: Colour, pattern or general look? Biol. J. Linn. Soc. 2009, 98, 234–242. [Google Scholar] [CrossRef]
- Hotová Svádová, K.; Exnerová, A.; Kopećková, M.; Štys, P. Predator dependent mimetic complexes: Do passerine birds avoid Central European red-and-black Heteroptera? Eur. J. Entomol. 2010, 107, 349–355. [Google Scholar] [CrossRef]
- Průchová, A.; Nedvěd, O.; Veselý, P.; Ernestová, B.; Fuchs, R. Visual warning signals of the ladybird Harmonia axyridis: The avian predators’ point of view. Entomol. Exp. Applicata 2014, 151, 128–134. [Google Scholar] [CrossRef]
- Exnerová, A.; Landová, E.; Štys, P.; Fuchs, R.; Prokopová, M.; Cehláriková, P. Reactions of passerine birds to aposematic and nonaposematic firebugs (Pyrrhocoris apterus; Heteroptera). Biol. J. Linn. Soc. 2003, 78, 517–525. [Google Scholar] [CrossRef]
- Marples, N.M.; Kelly, D.J. Neophobia and dietary conservatism: Two distinct processes? Evol. Ecol. 1999, 13, 641–653. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, N.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Alatalo, R.V.; Mappes, J. Tracking the evolution of warning signals. Nature 1996, 382, 708–710. [Google Scholar] [CrossRef]
- Chouteau, M.; Dezeure, J.; Sherratt, T.N.; Llaurens, V.; Joron, M. Similar predator aversion for natural prey with diverse toxicity levels. Anim. Behav. 2019, 153, 49–59. [Google Scholar] [CrossRef]
- Chipanovska, N.; Župunski, V.; Vittori, M. Differences in lobed gland structure and distribution in terrestrial isopods (Oniscidea). Nauplius 2025, 33, e20250532. [Google Scholar] [CrossRef]
- Ďurajková, B.; Tuf, I.H.; Vittori, M. Repugnatorial glands and aposematism in terrestrial isopods (Isopoda: Oniscidea: Armadillidiidae, Porcellionidae). J. Crustac. Biol. 2025, 45, ruaf014. [Google Scholar] [CrossRef]
- Exnerová, A.; Svádová, K.; Fousová, P.; Fučíková, E.; Ježová, D.; Niederlová, A.; Kopečková, M.; Štys, P. European birds and aposematic Heteroptera: Review of comparative experiments. Bull. Insectology 2008, 61, 163–165. [Google Scholar]
- Kazemi, B.; Gamberale-Stille, G.; Tullberg, B.S.; Leimar, O. Stimulus salience as an explanation for imperfect mimicry. Curr. Biol. 2014, 24, 965–969. [Google Scholar] [CrossRef]
- Fabricant, S.A.; Exnerová, A.; Ježová, D.; Štys, P. Scared by shiny? The value of iridescence in aposematic signalling of the hibiscus harlequin bug. Anim. Behav. 2014, 90, 315–325. [Google Scholar] [CrossRef]
- MacDougall, A.; Dawkins, M.S. Predator discrimination error and the benefits of Müllerian mimicry. Anim. Behav. 1998, 55, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Andreu, J.; Barba, E. Breeding dispersal of Great Tits Parus major in a homogeneous habitat: Effects of sex, age, and mating status. Ardea-Wageningen 2006, 94, 45–58. [Google Scholar]
- Adamová-Ježová, D.; Hospodková, E.; Fuchsová, L.; Štys, P.; Exnerová, A. Through experience to boldness? Deactivation of neophobia towards novel and aposematic prey in three European species of tits (Paridae). Behav. Process. 2016, 131, 24–31. [Google Scholar] [CrossRef]
- Tuf, I.H.; Drábková, L.; Šipoš, J. Personality affects defensive behaviour of Porcellio scaber (Isopoda, Oniscidea). ZooKeys 2015, 515, 159–171. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Predictor | Data Type | Data Included | Chi | DF | p |
|---|---|---|---|---|---|---|
| attacking | prey type | binomial | all | 37.575 | 9 | <0.001 |
| eating | prey type | binomial | only attacking | 62.303 | 9 | <0.001 |
| observing | prey type | gaussian | all | 31.97 | 7 | <0.001 |
| disgust | prey type * attack | poisson | all | 12.256 | 9 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ďurajková, B.; Veselý, P.; Tuf, I.H. Function of Vivid Coloration of Terrestrial Isopods from the Point of View of an Avian Predator. Insects 2025, 16, 662. https://doi.org/10.3390/insects16070662
Ďurajková B, Veselý P, Tuf IH. Function of Vivid Coloration of Terrestrial Isopods from the Point of View of an Avian Predator. Insects. 2025; 16(7):662. https://doi.org/10.3390/insects16070662
Chicago/Turabian StyleĎurajková, Barbora, Petr Veselý, and Ivan Hadrián Tuf. 2025. "Function of Vivid Coloration of Terrestrial Isopods from the Point of View of an Avian Predator" Insects 16, no. 7: 662. https://doi.org/10.3390/insects16070662
APA StyleĎurajková, B., Veselý, P., & Tuf, I. H. (2025). Function of Vivid Coloration of Terrestrial Isopods from the Point of View of an Avian Predator. Insects, 16(7), 662. https://doi.org/10.3390/insects16070662