
Histopathological and Transcriptional Changes in Silkworm Larval Gonads in Response to Chlorfenapyr Exposure

Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Silkworm Samples and Chlorfenapyr Stress
2.2. Ovary and Testis Histopathology
2.3. Transcriptome Sequencing (RNA-Seq) and Analysis
2.4. Differential Gene Expression and Gene Enrichment Analyses
2.5. Creating a Protein–Protein Interaction (PPI) Network
2.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
3. Results
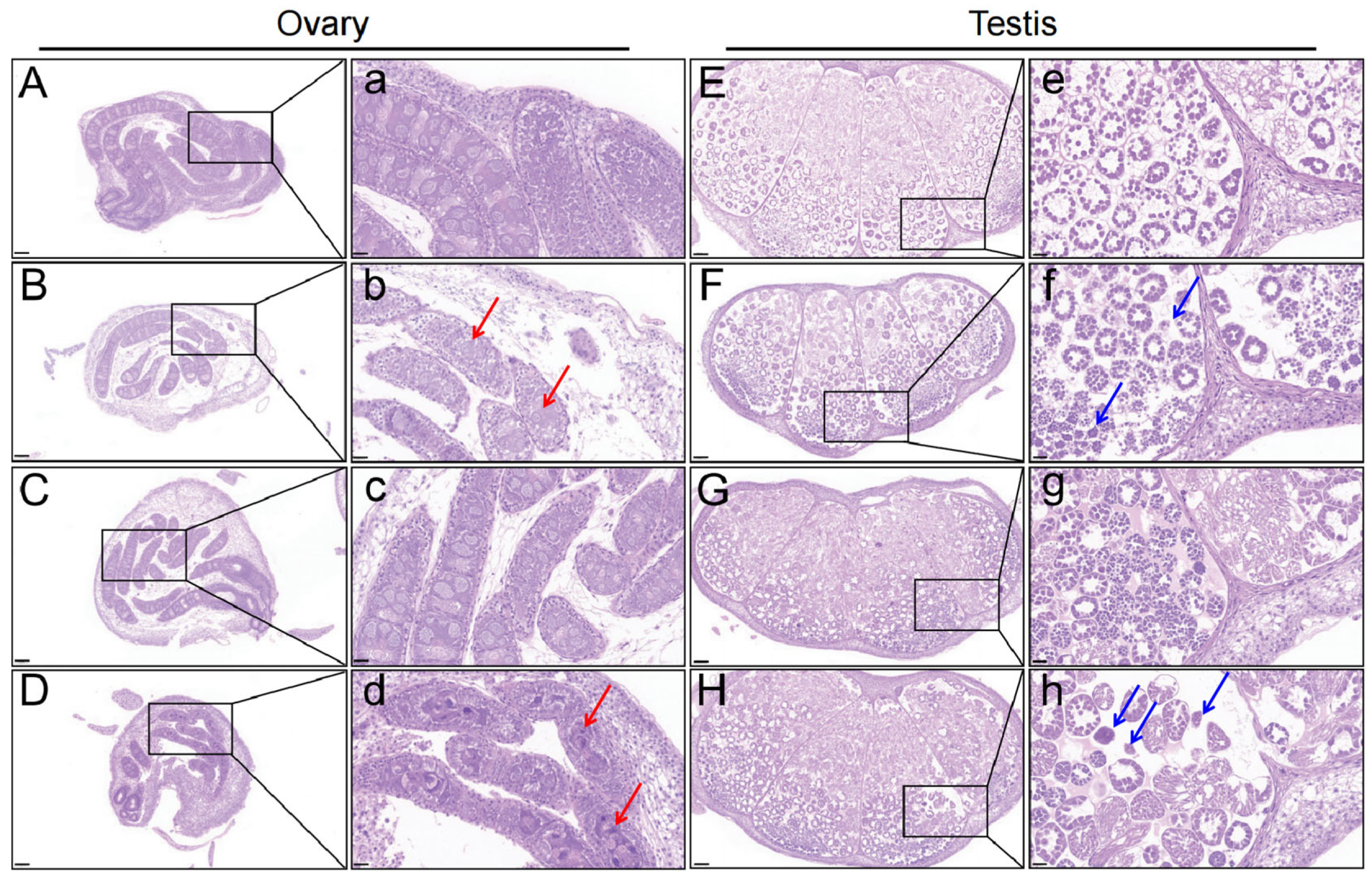
3.1. Structure of Larval Silkworm Gonad After Chlorfenapyr Stress
3.2. Analysis of Differentially Expressed Genes Between Treatments
3.3. KEGG Pathway Enrichment Analysis of DEGs
3.4. Transcriptional Changes of P450, Glutathione S-Transferase (GST), and Insect Hormones
3.5. Prediction of Transcription Factors and Their Expression Patterns
3.6. Analysis of PPI Network
3.7. Validation of Gene Expression Patterns After Exposure to Chlorfenapyr
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEGs | Differentially expressed genes |
| HE | Hematoxylin–eosin |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene ontology |
| qPCR | Quantitative real-time polymerase chain reaction |
| PPI | Protein–protein interaction |
| PCA | Principal component analysis |
| HCA | Hierarchical cluster analysis |
| MCC | Maximal clique centrality |
| JH | Juvenile hormone |
| JHAMT | Juvenile hormone acid methyltransferase |
| JHEH | Juvenile hormone epoxide hydrolase |
| CYP | Cytochrome P450 |
| GST | Glutathione S-transferase |
References
- Rodriguez-Ortiz, L.M.; Hincapie, C.A.; Hincapie-Llanos, G.A.; Osorio, M. Potential uses of silkworm pupae (Bombyx mori L.) in food, feed, and other industries: A systematic review. Front. Insect Sci. 2024, 4, 1445636. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.L.; Han, M.J.; Lu, K.P.; Tai, S.S.; Liang, S.B.; Liu, Y.C.; Hu, H.; Shen, J.H.; Long, A.X.; Zhan, C.Y.; et al. High-resolution silkworm pan-genome provides genetic insights into artificial selection and ecological adaptation. Nat. Commun. 2022, 13, 5619. [Google Scholar] [CrossRef]
- Ma, L.; Xie, Y.; Gu, Z.Y.; Wang, B.B.; Li, F.C.; Xu, K.Z.; Shen, W.D.; Li, B. Characteristics of phoxim-exposed gene transcription in the silk gland of silkworms. Pestic. Biochem. Physiol. 2013, 107, 391–397. [Google Scholar] [CrossRef]
- Chen, Q.Q.; Sun, S.M.; Yang, X.; Yan, H.H.; Wang, K.Y.; Ba, X.C.; Wang, H.Y. Sublethal effects of neonicotinoid insecticides on the development, body weight and economic characteristics of silkworm. Toxics 2023, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Su, Y.; Liu, X.Y.; Qi, R.N.; Li, F.C.; Li, B.; Sun, H.N. Low concentration of indoxacarb interferes with the growth and development of silkworm by damaging the structure of midgut cells. Pestic. Biochem. Physiol. 2023, 195, 105567. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.Z.; Lan, H.L.; He, C.H.; Wei, Y.T.; Lu, Q.Y.; Cai, K.P.; Yu, D.L.; Yin, X.C.; Li, Y.Z.; Lv, J.C. Toxicological effects of trace amounts of pyriproxyfen on the midgut of non-target insect silkworm. Pestic. Biochem. Physiol. 2022, 188, 105266. [Google Scholar] [CrossRef]
- Cheng, J.; Chen, Y.L.; Wang, W.D.; Zhu, X.Q.; Jiang, Z.L.; Liu, P.; Du, L.W. Chlorfenapyr poisoning: Mechanisms, clinical presentations, and treatment strategies. World J. Emerg. Med. 2024, 15, 214–219. [Google Scholar] [CrossRef]
- Black, B.C.; Hollingworth, R.M.; Ahammadsahib, K.I.; Kukel, C.D.; Donovan, S. Insecticidal action and mitochondrial uncoupling activity of AC-303,630 and related halogenated pyrroles. Pestic. Biochem. Physiol. 1994, 50, 115–128. [Google Scholar] [CrossRef]
- Saito, S. Effects of pyridalyl on ATP concentrations in cultured Sf9 cells. J. Pestic. Sci. 2005, 30, 403–405. [Google Scholar] [CrossRef]
- Wang, Q.G.; Xu, M.C.; An, E.; Zhang, X.; Li, X.; Wei, J.Z.; Liu, X.G. Sublethal and transgenerational effects of chlorfenapyr on physiological and developmental traits of Spodoptera frugiperda. Entomol. Gen. 2024, 44, 1341–1349. [Google Scholar] [CrossRef]
- Jia, B.T.; Zhang, J.L.; Hong, S.S.; Chang, X.L.; Li, X.C. Sublethal effects of chlorfenapyr on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2023, 79, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Wang, Q.H.; Ding, J.F.; Wang, Y.; Zhang, Z.Q.; Liu, F.; Mu, W. Sublethal effects of chlorfenapyr on the life table parameters, nutritional physiology and enzymatic properties of Bradysia odoriphaga (Diptera: Sciaridae). Pestic Biochem Physiol 2018, 148, 93–102. [Google Scholar] [CrossRef]
- Aljabr, A.M.; Rizwan-ul-Haq, M.; Hussain, A.; Al-Mubarak, A.I.; Al-Ayied, H.Y. Establishing midgut cell culture from Rhynchophorus ferrugineus (Olivier) and toxicity assessment against ten different insecticides. In Vitro Cell Dev. Biol. Anim. 2014, 50, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; Wang, X.; Yang, X.; Ma, Z.Y.; Liu, P.; Tang, S.Y.; Zhao, M.; Chen, H.J.; Qiu, Q.; Tang, M.H.; et al. Toxicokinetics, in vivo metabolic profiling and tissue distribution of chlorfenapyr in mice. Arch. Toxicol. 2024, 98, 3763–3775. [Google Scholar] [CrossRef]
- Shao, Y.; Xin, X.D.; Liu, Z.X.; Wang, J.; Zhang, R.; Gui, Z.Z. Transcriptional response of detoxifying enzyme genes in Bombyx mori under chlorfenapyr exposure. Pestic. Biochem. Physiol. 2021, 177, 104899. [Google Scholar] [CrossRef]
- Li, T.; Liu, M.; Liu, Z.H.; Fan, Y.H.; Liao, P.F.; Dong, Z.P. Effects of chlorfenapyr stress on economic traits and fecundity of Bombyx mori. Guizhou Agr. Sci. 2023, 51, 93–100. (In Chinese) [Google Scholar]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods. 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Bu, D.C.; Luo, H.T.; Huo, P.P.; Wang, Z.H.; Zhang, S.; He, Z.H.; Wu, Y.; Zhao, L.H.; Liu, J.J.; Guo, J.C.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.; Pyysalo, S.; et al. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2022, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Li, T.; Qin, S.; Sun, X.; Zhang, K.X.; Ding, X.Y.; Wang, X.Y.; Li, M.W. Transcriptome analysis reveals distinct innate immunity and ribosomal response at early stage of AcMNPV infection in haemocyte of silkworm resistant and susceptible strains. J. Asia-Pac. Entomol. 2022, 25, 101938. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Raghavendra, K.; Barik, T.K.; Sharma, P.; Bhatt, R.M.; Srivastava, H.C.; Sreehari, U.; Dash, A.P. Chlorfenapyr: A new insecticide with novel mode of action can control pyrethroid resistant malaria vectors. Malar. J. 2011, 10, 16. [Google Scholar] [CrossRef]
- Lu, K.; Song, Y.Y.; Zeng, R.S. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef]
- Wang, R.L.; Liu, S.W.; Baerson, S.R.; Qin, Z.; Ma, Z.H.; Su, Y.J.; Zhang, J.E. Identification and functional analysis of a novel cytochrome P450 gene CYP9A105 associated with pyrethroid detoxification in Spodoptera exigua Hubner. Int. J. Mol. Sci. 2018, 19, 737. [Google Scholar] [CrossRef]
- Liu, Z.X.; Xing, X.R.; Liang, X.H.; Ding, J.H.; Li, Y.-J.C.; Shao, Y.; Wu, F.A.; Wang, J.; Sheng, S. The role of Glutathione-S-transferases in phoxim and chlorfenapyr tolerance in a major mulberry pest, Glyphodes pyloalis walker (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2022, 181, 105004. [Google Scholar] [CrossRef]
- Bolt, H.M.; Thier, R. Relevance of the deletion polymorphisms of the glutathione S-transferases GSTT1 and GSTM1 in pharmacology and toxicology. Curr. Drug Metab. 2006, 7, 613–628. [Google Scholar] [CrossRef]
- Krishnamurthy, H.K.; Rajavelu, I.; Pereira, M.; Jayaraman, V.; Krishna, K.; Wang, T.; Bei, K.; Rajasekaran, J.J. Inside the genome: Understanding genetic influences on oxidative stress. Front. Genet. 2024, 15, 1397352. [Google Scholar] [CrossRef]
- Guo, P.C.; Zhang, Y.S.; Zhang, L.; Xu, H.Y.; Zhang, H.; Wang, Z.; Jiang, Y.L.; Molloy, D.; Zhao, P.; Xia, Q.Y. Structural basis for juvenile hormone biosynthesis by the juvenile hormone acid methyltransferase. J. Biol. Chem. 2021, 297, 101234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, H.Y.; Zhang, Y.S.; Zhang, H.; Wang, Z.; Guo, P.C.; Zhao, P. Structural characterization and functional analysis of juvenile hormone acid methyltransferase JHAMT3 from the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2022, 151, 103863. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.V.; Maestro, J.L. Expression of juvenile hormone acid O-methyltransferase and juvenile hormone synthesis in Blattella germanica. Insect Sci. 2018, 25, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.Y.; Li, Q.; Zhou, L.T.; Meng, Q.W.; Lu, F.G.; Guo, W.C.; Li, G.Q. Knockdown of juvenile hormone acid methyl transferase severely affects the performance of Leptinotarsa decemlineata (Say) larvae and adults. Pest Manag. Sci. 2016, 72, 1231–1241. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Huang, Z.S.; Zheng, X.Y.; Zheng, Z.H.; Yao, D.; Zhang, Y.L.; Aweya, J.J. The juvenile hormone epoxide hydrolase homolog in Penaeus vannamei plays immune-related functions. Dev. Comp. Immunol. 2022, 132, 104410. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Liu, X.J.; Shiotsuki, T.; Wang, Z.S.; Xu, X.; Huang, Y.P.; Li, M.W.; Li, K.; Tan, A.J. Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect Biochem. Mol. Biol. 2017, 81, 72–79. [Google Scholar] [CrossRef]
- Li, C.L.; Zhang, H.; Gao, R.; Zuo, W.D.; Liu, Y.Y.; Hu, H.; Luan, Y.; Lu, C.; Tong, X.L.; Dai, F.Y. Identification and effect of Zf-AD-containing C2H2 zinc finger genes on BmNPV replication in the silkworm (Bombyx mori). Pestic. Biochem. Physiol. 2020, 170, 104678. [Google Scholar] [CrossRef]
- Dhillon, B.; Aleithan, F.; Abdul-Sater, Z.; Abdul-Sater, A.A. The evolving role of TRAFs in mediating inflammatory responses. Front. Immunol. 2019, 10, 104. [Google Scholar] [CrossRef]
- Han, X.F.; Che, M.Y.; Su, X.; Tian, J.Y.; Liu, L.H.; Nie, P.; Wang, S. Molecular characterization of TRAF gene family in snakehead (Channa argus). Fish Shellfish Immunol. 2025, 158, 110135. [Google Scholar] [CrossRef]
- Wang, X.F.; Gao, S.; Hao, Z.X.; Tang, T.; Liu, F.S. Involvement of TRAF6 in regulating immune defense and ovarian development in Musca domestica. Int. J. Biol. Macromol. 2020, 153, 1262–1271. [Google Scholar] [CrossRef] [PubMed]
- Rumbo, M.; Pagone, V.; Piulachs, M.D. Diverse functions of the ecdysone receptor (EcR) in the panoistic ovary of the German cockroach. Insect Biochem. Mol. Biol. 2023, 156, 103935. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Song, H.Y.; Wang, Y.; Liu, Z.G.; Wang, H.F.; Xu, B.H. 20-hydroxyecdysone upregulates ecdysone receptor (EcR) gene to promote pupation in the honeybee, Apis mellifera ligustica. Integr. Comp. Biol. 2023, 63, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Liu, C.B.; Zhang, J.F.; Lu, Y.Y.; Jiang, B.T.; Xiong, H.B.; Li, C.X. Pyruvate dehydrogenase kinase regulates macrophage polarization in metabolic and inflammatory diseases. Front. Immunol. 2023, 14, 1296687. [Google Scholar] [CrossRef]
- He, Q.Y.; Zhang, Y.X. Kr-h1, a cornerstone gene in insect life history. Front. Physiol. 2022, 13, 905441. [Google Scholar] [CrossRef]
- Myers, T.D.; Palladino, M.J. Newly discovered roles of triosephosphate isomerase including functions within the nucleus. Mol. Med. 2023, 29, 18. [Google Scholar] [CrossRef]
- Han, J.H.; Lee, E.J.; Park, W.; Ha, K.T.; Chung, H.S. Natural compounds as lactate dehydrogenase inhibitors: Potential therapeutics for lactate dehydrogenase inhibitors-related diseases. Front. Pharmacol. 2023, 14, 1275000. [Google Scholar] [CrossRef]
- Zeng, B.S.; Huang, Y.P.; Xu, J.; Shiotsuki, T.; Bai, H.; Palli, S.R.; Huang, Y.P.; Tan, A.J. The FOXO transcription factor controls insect growth and development by regulating juvenile hormone degradation in the silkworm, Bombyx mori. J. Biol. Chem. 2017, 292, 11659–11669. [Google Scholar] [CrossRef]
- Song, J.B.; Li, Z.Q.; Zhou, L.; Chen, X.; Sew, W.Q.G.; Herranz, H.; Ye, Z.L.; Olsen, J.V.; Li, Y.; Nygaard, M.; et al. FOXO-regulated OSER1 reduces oxidative stress and extends lifespan in multiple species. Nat. Commun. 2024, 15, 7144. [Google Scholar] [CrossRef]
- Dulce, S.; Thomas-Wolf, V.; Lina, M.; Stijn, V.d.B.; Bart, G.; Filip, V.N.; Anna, K.; Luc, S.; Niels, W.; Jozef, V.B. PIWI proteins play an antiviral role in lepidopteran cell lines. Viruses 2022, 14, 1442. [Google Scholar] [CrossRef]
- Kiuchi, T.; Shoji, K.; Izumi, N.; Tomari, Y.; Katsuma, S. Non-gonadal somatic piRNA pathways ensure sexual differentiation, larval growth, and wing development in silkworms. PLoS Genet. 2023, 19, e1010912. [Google Scholar] [CrossRef] [PubMed]
- Anger, A.M.; Armache, J.-P.; Berninghausen, O.; Habeck, M.; Subklewe, M.; Wilson, D.N.; Beckmann, R. Structures of the human and Drosophila 80S ribosome. Nature 2013, 497, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.L.; Gang, X.X.; Zhang, T.L.; Wei, L.; Yang, X.X.; Li, Z.; Yang, Y.; Song, L.; Wang, P.; Peng, J.; et al. Protein kinase A-mediated phosphorylation of the Broad-Complex transcription factor in silkworm suppresses its transcriptional activity. J. Biol. Chem. 2017, 292, 12460–12470. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.L.; Ye, W.T.; Jiang, X.J.; Feng, P.; Zhu, Q.Y.; Sun, H.N.; Li, F.C.; Wei, J.; Li, B. Effect of tachinid parasitoid Exorista japonica on the larval development and pupation of the host silkworm Bombyx mori. Front. Physiol. 2022, 13, 824203. [Google Scholar] [CrossRef]
- Wu, J.J.; Cheng, M.D.; Ze, L.J.; Shen, C.H.; Jin, L.; Li, G.Q. Dissecting the isoform-specific roles of FTZ-F1 in the larval-larval and larval-pupal ecdyses in Henosepilachna vigintioctopunctata. Insects 2022, 13, 228. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Li, H.J.; Liu, X.M.; Li, H.L.; Lan, Q.Y.; Wu, H.H.; Wang, Y.L.; Zhang, J.Z.; Zhao, X.M. Nuclear receptor FTZ-F1 controls locust molt by regulating the molting process of Locusta migratoria. Insects 2024, 15, 237. [Google Scholar] [CrossRef]
- Petibon, C.; Ghulam, M.M.; Catala, M.; Elela, S.A. Regulation of ribosomal protein genes: An ordered anarchy. WIREs RNA 2020, 12, e1632. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Hu, C.; Liu, Z.; Li, Q.; Fan, Y.; Liao, P.; Liu, M.; Yang, W.; Li, X.; Dong, Z. Histopathological and Transcriptional Changes in Silkworm Larval Gonads in Response to Chlorfenapyr Exposure. Insects 2025, 16, 619. https://doi.org/10.3390/insects16060619
Li T, Hu C, Liu Z, Li Q, Fan Y, Liao P, Liu M, Yang W, Li X, Dong Z. Histopathological and Transcriptional Changes in Silkworm Larval Gonads in Response to Chlorfenapyr Exposure. Insects. 2025; 16(6):619. https://doi.org/10.3390/insects16060619
Chicago/Turabian StyleLi, Tao, Changxiong Hu, Zenghu Liu, Qiongyan Li, Yonghui Fan, Pengfei Liao, Min Liu, Weike Yang, Xingxing Li, and Zhanpeng Dong. 2025. "Histopathological and Transcriptional Changes in Silkworm Larval Gonads in Response to Chlorfenapyr Exposure" Insects 16, no. 6: 619. https://doi.org/10.3390/insects16060619
APA StyleLi, T., Hu, C., Liu, Z., Li, Q., Fan, Y., Liao, P., Liu, M., Yang, W., Li, X., & Dong, Z. (2025). Histopathological and Transcriptional Changes in Silkworm Larval Gonads in Response to Chlorfenapyr Exposure. Insects, 16(6), 619. https://doi.org/10.3390/insects16060619