
Low-Cost–High-Efficacy Control of Faba Bean Aphids by Fungal and Chemical Insecticides Co-Applied at Low and Sublethal Rates

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal and Chemical Insecticides
2.2. Faba Bean Crop
2.3. Setting up Field Trials
2.4. Sampling of Aphid Density for Control Efficacy
2.5. Weather Data
2.6. Statistical Analysis
3. Results
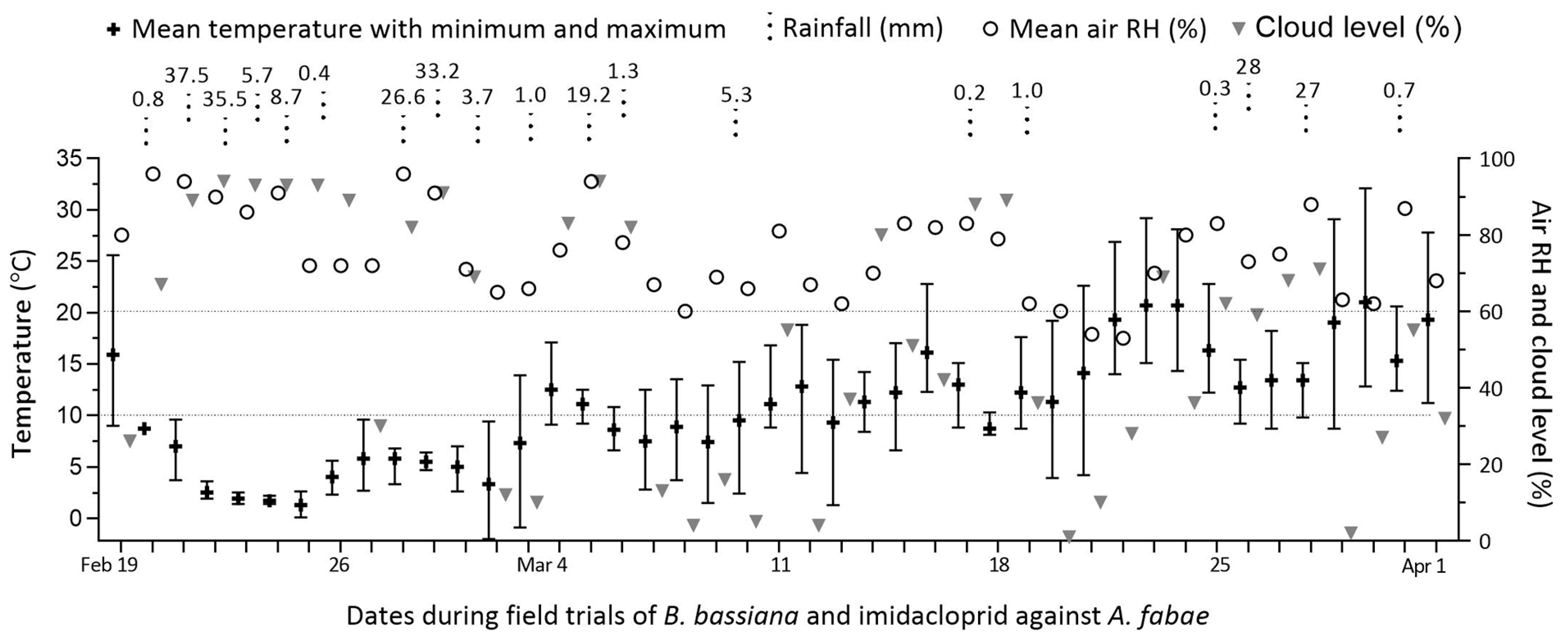
3.1. Weather Changes During Field Trials
3.2. Suppression of Aphid Population
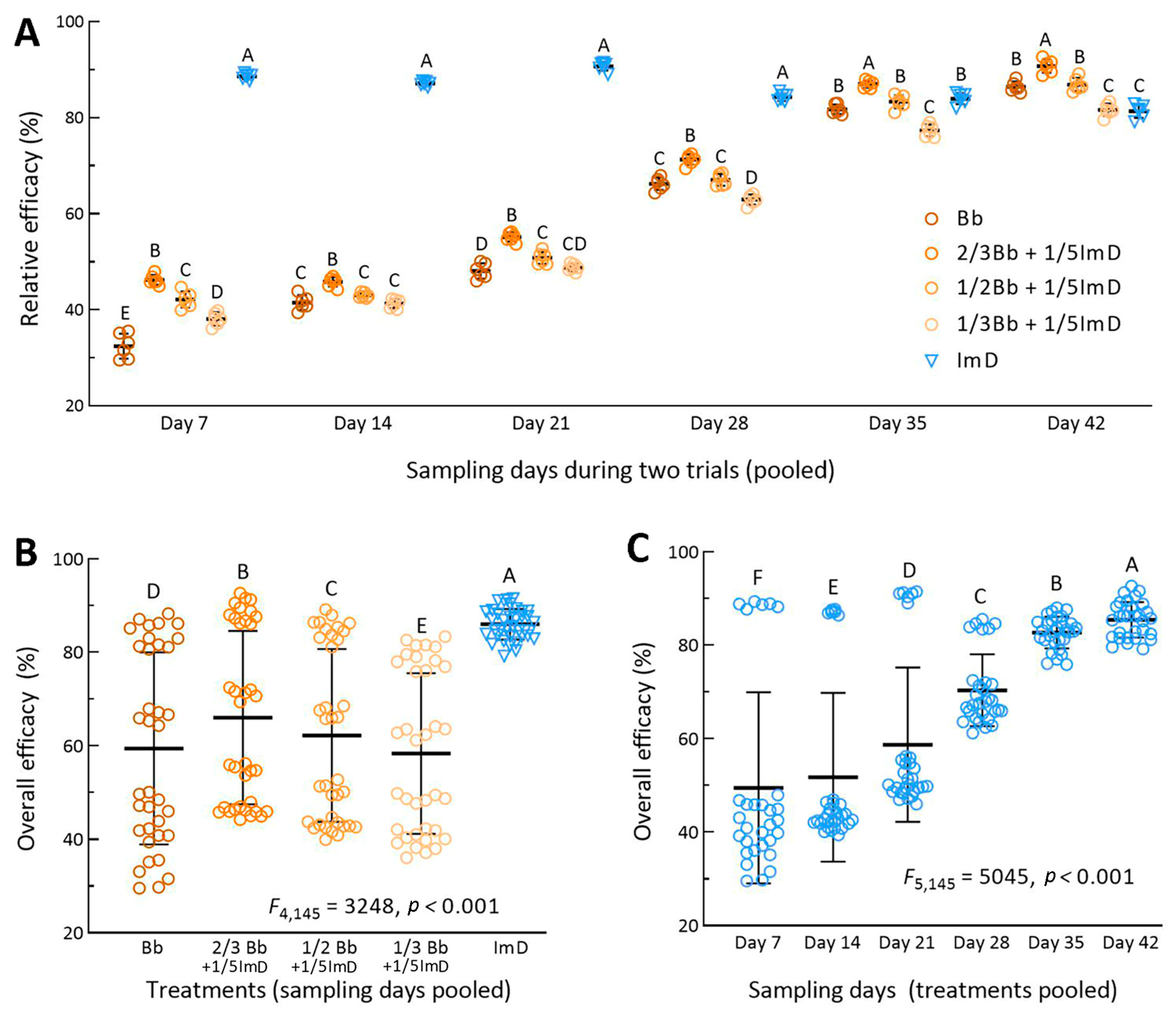
3.3. Efficacies of Different Treatments Against A. fabae
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akca, I.; Ayvaz, T.; Yazici, E.; Smith, C.L.; Chi, H. Demography and population projection of Aphis fabae (Hemiptera: Aphididae): With additional comments on life table research criteria. J. Econ. Entomol. 2015, 108, 1466–1478. [Google Scholar] [CrossRef]
- Shih, P.Y.; Sugio, A.; Simon, J.C. Molecular mechanisms underlying host plant specificity in aphids. Annu. Rev. Entomol. 2023, 68, 431–450. [Google Scholar] [CrossRef]
- Almogdad, M.; Lavrukaite, K.; Semaskiene, R. Temporal analysis of the relationship between black bean aphid (Aphis fabae) infestation and meteorological conditions in faba bean (Vicia faba). Agronomy 2024, 14, 1182. [Google Scholar] [CrossRef]
- Edwards, O.R.; Franzmann, B.; Thackray, D.; Micic, S. Insecticide resistance and implications for future aphid management in Australian grains and pastures: A review. Aust. J. Exp. Agric. 2008, 48, 1523–1530. [Google Scholar] [CrossRef]
- Bass, C.; Nauen, R. The molecular mechanisms of insecticide tesistance in aphid crop pests. Insect Biochem. Mol. Biol. 2023, 156, 103937. [Google Scholar] [CrossRef]
- Mc Namara, L.; Gauthier, K.; Walsh, L.; Thebaud, G.; Gaffney, M.; Jacquot, E. Management of yellow dwarf Disease in Europe in a post-neonicotinoid agriculture. Pest Manag. Sci. 2020, 76, 2276–2285. [Google Scholar] [CrossRef]
- Ndakidemi, B.J.; Mbega, E.R.; Ndakidemi, P.A.; Stevenson, P.C.; Belmain, S.R.; Arnold, S.E.J.; Woolley, V.C. Natural pest regulation and its compatibility with other crop protection practices in smallholder bean farming systems. Biology 2021, 10, 805. [Google Scholar] [CrossRef]
- Francis, F.; Then, C.; Francis, A.; Gbangbo, Y.A.C.; Iannello, L.; Ben Fekih, I. Complementary strategies for biological control of aphids and related virus transmission in sugar beet to replace neonicotinoids. Agriculture 2022, 12, 1663. [Google Scholar] [CrossRef]
- Qubbaj, T.; Samara, R. Efficacy of three entomopathogenic fungi Beauveria bassiana, Metarhizium anisopliae and Lecanicillium lecanii Isolates against black bean aphid, Aphis fabae (Scop.) (Hemiptera: Aphididae) on faba bean (Vicia faba L.). Legume Res. 2022, 45, 1572–1579. [Google Scholar] [CrossRef]
- Mohammed, A.A. Lecanicillium muscarium and Adalia bipunctata combination for the control of black bean aphid, Aphis fabae. BioControl 2018, 63, 277–287. [Google Scholar] [CrossRef]
- Seiedy, M.; Heydari, S.; Tork, M. Orientation of Hippodamia variegata (Coleoptera: Coccinellidae) to healthy and Beauveria bassiana-Infected Aphis fabae (Hemiptera: Aphididae) in an olfactometer system. Turk. J. Zool. 2015, 39, 53–58. [Google Scholar] [CrossRef]
- Nordey, T.; Boni, S.B.; Agbodzavu, M.K.; Mwashimaha, R.; Mlowe, N.; Ramasamy, S.; Deletre, E. Comparison of biological methods to control Aphis fabae Scopoli (Hemiptera: Aphididae) on kalanchoe crops in East Africa. Crop Prot. 2021, 142, 105520. [Google Scholar] [CrossRef]
- Fernández-Grandon, G.M.; Harte, S.J.; Ewany, J.; Bray, D.; Stevenson, P.C. Additive effect of botanical insecticide and entomopathogenic fungi on pest mortality and the behavioral response of its natural rnemy. Plants 2020, 9, 173. [Google Scholar] [CrossRef] [PubMed]
- Benbelkhir, F.Z.; Allali, K.; Benadjila, A.; Goudjal, Y.; Medjekal, S.; Zamoum, M. Development of bioinsecticide based on Streptomyces griseoflavus PAL114 for control of black bean aphids Aphis fabae. Biocontrol Sci. Technol. 2024, 34, 736–753. [Google Scholar] [CrossRef]
- Jensen, R.E.; Enkegaard, A.; Steenberg, T. Increased fecundity of Aphis fabae on Vicia faba plants following seed or leaf inoculation with the entomopathogenic fungus Beauveria bassiana. PLoS ONE 2019, 14, e0223616. [Google Scholar] [CrossRef]
- Frank, S.D. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biol. Control 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Pell, J.K.; Hannam, J.J.; Steinkraus, D.C. Conservation biological control using fungal entomopathogens. BioControl 2010, 55, 187–198. [Google Scholar] [CrossRef]
- Azimi, S.; Amini, R. Population density of Aphis fabae Scopoli (Hemiptera, Aphididae) and its natural enemies in intercropping of faba bean (Vicia faba L.) and dragonhead (Dracocephalum moldavica L.). J. Biodivers. Environ. Sci. 2015, 6, 380–388. [Google Scholar]
- Emaru, A.; Nyaanga, J.G.; Saidi, M. Integrating Metarhizium anisopliae entomopathogenic fungi with border cropping reduces black bean aphids (Aphis fabae) damage and enhances yield and quality of French bean. Heliyon 2024, 10, e33037. [Google Scholar] [CrossRef]
- Meradsi, F.; Laamari, M. Population dynamics and biological parameters of Aphis fabae Scopoli on five broad bean cultivars. Int. J. Biosci. 2016, 9, 58–68. [Google Scholar]
- Wamonje, F.O.; Tungadi, T.D.; Murphy, A.M.; Pate, A.E.; Woodcock, C.; Caulfield, J.C.; Mutuku, J.M.; Cunniffe, N.J.; Bruce, T.J.A.; Gilligan, C.A.; et al. Three aphid-transmitted viruses encourage vector migration from infected common bean (Phaseolus vulgaris) plants through a combination of volatile and surface cues. Front. Plant Sci. 2020, 11, 613772. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Stahl, A.; Rostás, M.; Will, T. Temporal and species-specific resistance of sugar beet to green peach aphid and black bean aphid: Mechanisms and implications for breeding. Pest Manag. Sci. 2024, 80, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Skovgård, H.; Stoddard, F.L. Reproductive potential of the black bean aphid (Aphis fabae Scop.) on a range of faba bean (Vicia faba L.) accessions. Legume Sci. 2023, 5, e199. [Google Scholar] [CrossRef]
- de Faria, M.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Arthurs, S.; Dara, S.K. Microbial Biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13–21. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Parker, P.E.; Tsai, J.H. Laboratory and field evaluation of hyphomycete insect pathogenic fungi for control of brown citrus aphid (Homoptera: Aphididae). Environ. Entomol. 1999, 28, 315–321. [Google Scholar] [CrossRef]
- Vandenberg, J.D.; Sandvol, L.E.; Jaronski, S.T.; Jackson, M.A.; Souza, E.J.; Halbert, S.E. Efficacy of fungi for control of Russian wheat aphid (Homoptera: Aphididae) in irrigated wheat. Southwest Entomol. 2001, 26, 73–85. [Google Scholar]
- Wraight, S.P.; Carruthers, R.I.; Jaronski, S.T.; Bradley, C.A.; Garza, C.J.; Galaini-Wraight, S. Evaluation of the entomopathogenic fungi Beauveria bassiana and Paecilomyces fumosoroseus for microbial control of the silverleaf whitefly, Bemisia argentifolii. Biol. Control 2000, 17, 203–217. [Google Scholar] [CrossRef]
- Tang, J.F.; Liu, X.Y.; Ding, Y.C.; Jiang, W.J.; Xie, J.Q. Evaluation of Metarhizium anisopliae for rice planthopper control and its synergy with selected insecticides. Crop Prot. 2019, 121, 132–138. [Google Scholar] [CrossRef]
- Peng, G.X.; Xie, J.Q.; Guo, R.; Keyhani, N.O.; Zeng, D.Y.; Yang, P.Y.; Xia, Y.X. Long-term field evaluation and large-scale application of a Metarhizium anisopliae strain for controlling major rice pests. J. Pest Sci. 2021, 94, 969–980. [Google Scholar] [CrossRef]
- Peng, Y.F.; Tang, J.F.; Hong, M.S.; Xie, J.Q. Suppression of rice planthopper populations by the entomopathogenic fungus Metarhizium anisopliae without affecting the rice microbiota. Appl. Environ. Microbiol. 2020, 86, e01337-20. [Google Scholar] [CrossRef]
- Qi, D.Y.; Xu, W.Y.; Shao, Y.Z.; Feng, J.R.; Feng, M.G.; Tong, S.M. Mycoinsecticides applied in late afternoon are more efficacious against rice leaf-rolling insect pests than those in the morning. Biol. Control 2023, 186, 105352. [Google Scholar] [CrossRef]
- Xu, W.Y.; Wen, Z.X.; Li, X.J.; Hu, E.Z.; Qi, D.Y.; Feng, M.G.; Tong, S.M. Timing of fungal insecticide application to avoid solar ultraviolet irradiation enhances field control of rice planthoppers. Insects 2023, 14, 307. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.Y.; Shao, Y.Z.; Yang, R.; Liu, C.L.; Feng, G.H.; Pan, W.Y.; Feng, M.G.; Tong, S.M. Emulsifiable oil-formulated Beauveria bassiana competes with imidacloprid for seasonal control of cereal aphids in Zhejiang, China. Pest Manag. Sci. 2024, 80, 2929–2936. [Google Scholar] [CrossRef]
- Tong, S.M.; Qi, D.Y.; Liu, C.L.; Feng, G.H.; Pan, W.Y.; Shao, Y.Z.; Yang, R.; Feng, M.G. Comparative efficacies of two fungal insecticides in seasonal control of cereal aphids heavily infesting winter wheat. BioControl 2024, 69, 437–448. [Google Scholar] [CrossRef]
- Ye, S.D.; Dun, Y.H.; Feng, M.G. Time and concentration dependent interactions of Beauveria bassiana with sublethal rates of imidacloprid against the aphid pests Macrosiphoniella sanborni and Myzus persicae. Ann. Appl. Biol. 2005, 146, 459–468. [Google Scholar] [CrossRef]
- Skouras, P.J.; Darras, A.I.; Mprokaki, M.; Demopoulos, V.; Margaritopoulos, J.; Delis, C.; Stathas, G.J. Toxicity, sublethal and low dose effects of imidacloprid and deltamethrin on the aphidophagous predator Ceratomegilla undecimnotata (Coleoptera: Coccinellidae). Insects 2021, 12, 696. [Google Scholar] [CrossRef]
- Gao, Z.S.; Zhang, X.F.; Liu, H.T.; Zhang, W.J.; Mu, W. Feasibility for controlling wheat aphids by seed dressing with neonicotinoid insecticides. Acta Phytophyl. Sin. 2016, 43, 864–872. [Google Scholar]
- Zhang, P.; Zhang, X.F.; Zhao, Y.H.; Wei, Y.; Mu, W.; Liu, F. Effects of imidacloprid and clothianidin seed treatments on wheat aphids and their natural enemies on winter wheat. Pest Manag. Sci. 2016, 72, 1141–1149. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.P.; Li, X.R.; Zhu, X.; Zhang, Y.H. Efficacy of imidacloprid seed treatments against four wheat aphids under laboratory and field conditions. Plants 2023, 12, 238. [Google Scholar] [CrossRef]
- Xu, T.Y.; Zhang, S.; Liu, Y.; Ma, L.; Li, X.Q.; Zhang, Y.X.; Fan, Y.J.; Song, D.L.; Gao, X.W. Slow resistance evolution to neonicotinoids in field populations of wheat aphids revealed by insecticide resistance monitoring in China. Pest Manag. Sci. 2022, 78, 1428–1437. [Google Scholar] [CrossRef] [PubMed]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. BioControl 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Zhang, B.; Leonard, S.P.; Li, Y.Y.; Moran, N.A. Obligate bacterial endosymbionts limit thermal tolerance of insect host species. Proc. Natl. Acad. Sci. USA 2019, 116, 24712–24718. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xu, S.Y.; Luo, X.C.; Ying, S.H.; Feng, M.G. High photoreactivation activities of Rad2 and Rad14 in recovering insecticidal Beauveria bassiana from solar UV damage. J. Photochem. Photobiol. B-Biol. 2024, 251, 112849. [Google Scholar] [CrossRef]
- Feng, M.G.; Pu, X.Y. Time-concentration-mortality modeling of the synergistic interaction of Beauveria bassiana and imidacloprid against Nilaparvata lugens. Pest Manag. Sci. 2005, 61, 363–370. [Google Scholar] [CrossRef]
- Tian, L.; Feng, M.G. Evaluation of the time-concentration-mortality responses of Plutella xylostella larvae to the interaction of Beauveria bassiana with a nereistoxin analogue insecticide. Pest Manag. Sci. 2006, 62, 69–76. [Google Scholar] [CrossRef]
- Malsam, O.; Kilian, M.; Oerke, E.C.; Dehne, H.W. Oils for increased efficacy of Metarhizium anisopliae to control whiteflies. Biocontrol Sci. Technol. 2002, 12, 337–348. [Google Scholar] [CrossRef]
- Shi, W.B.; Feng, M.G.; Liu, S.S. Sprays of emulsifiable Beauveria bassiana formulation are ovicidal towards Tetranychus urticae (Acari: Tetranychidae) at various regimes of temperature and humidity. Exp. Appl. Acarol. 2008, 46, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pest. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- Goulson, D. REVIEW: An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, S.-M.; Qi, D.-Y.; Tang, X.-Y.; Shao, Y.-Z.; Hu, S.-T.; Zheng, Y.-P.; Zheng, X.-Y.; Yan, L.-L.; Feng, M.-G. Low-Cost–High-Efficacy Control of Faba Bean Aphids by Fungal and Chemical Insecticides Co-Applied at Low and Sublethal Rates. Insects 2025, 16, 565. https://doi.org/10.3390/insects16060565
Tong S-M, Qi D-Y, Tang X-Y, Shao Y-Z, Hu S-T, Zheng Y-P, Zheng X-Y, Yan L-L, Feng M-G. Low-Cost–High-Efficacy Control of Faba Bean Aphids by Fungal and Chemical Insecticides Co-Applied at Low and Sublethal Rates. Insects. 2025; 16(6):565. https://doi.org/10.3390/insects16060565
Chicago/Turabian StyleTong, Sen-Miao, Dan-Yi Qi, Xin-Yue Tang, Yan-Zhi Shao, Shi-Tong Hu, Yu-Piao Zheng, Xin-Yun Zheng, Ling-Li Yan, and Ming-Guang Feng. 2025. "Low-Cost–High-Efficacy Control of Faba Bean Aphids by Fungal and Chemical Insecticides Co-Applied at Low and Sublethal Rates" Insects 16, no. 6: 565. https://doi.org/10.3390/insects16060565
APA StyleTong, S.-M., Qi, D.-Y., Tang, X.-Y., Shao, Y.-Z., Hu, S.-T., Zheng, Y.-P., Zheng, X.-Y., Yan, L.-L., & Feng, M.-G. (2025). Low-Cost–High-Efficacy Control of Faba Bean Aphids by Fungal and Chemical Insecticides Co-Applied at Low and Sublethal Rates. Insects, 16(6), 565. https://doi.org/10.3390/insects16060565