Simple Summary
The aim of the study was to determine the effect of different doses of tested pyrethroids on the colony growth and metabolic activity of EPF from the Beauveria genus. In vitro, the effect of three pyrethroids (deltamethrin, λ-cyhalothrin, and α-cypermethrin) added to SDA medium at a dose 10 times lower than the recommended field dose (A), the recommended field dose (B), and 10 times higher than the recommended field dose (C) on colony growth and metabolic activity of B. bassiana and B. brongniartii was tested. The studies conducted showed that on the 20th day of the observation, λ-cyhalothrin used in the recommended field dose limited the growth of B. bassiana to the least extent in relation to the other tested pyrethroids. However, with respect to the fungus B. brongniartii, no toxic effect of this pyrethroid was found. λ-cyhalothrin used in the recommended field dose and 10 times lower than recommended significantly increased the metabolic activity of B. bassiana. Deltamethrin used in each of the tested concentrations significantly affected B. brongniartii viability.
Abstract
Pyrethroids are chemical insecticides used on a large scale in agriculture, horticulture, and forest protection. In order to reduce their use in IPM, alternative methods of controlling insect pests are introduced, such as the use of biopesticides based on entomopathogenic fungi (EPF). Species of the Beauveria genus are characterized by a very broad spectrum of action, which is why they are often used to produce preparations based on EPF. The aim of the study was to determine the effect of different doses of tested pyrethroids on the colony growth and metabolic activity of EPF from the Beauveria genus. In vitro, the effect of three pyrethroids (deltamethrin, λ-cyhalothrin, and α-cypermethrin) added to SDA medium at a dose 10 times lower than the recommended field dose (A), the recommended field dose (B), and 10 times higher than the recommended field dose (C) on colony growth and metabolic activity of B. bassiana and B. brongniartii was tested. The research carried out showed that pyrethroid insecticides used in the experiment showed various toxic effects towards the tested EPF of the genus Beauveria. The studies conducted showed that on the 20th day of the observation, λ-cyhalothrin used in the recommended field dose limited the growth of B. bassiana to the least extent in relation to the other tested pyrethroids. However, with respect to the fungus B. brongniartii, no toxic effect of this pyrethroid was found. Based on the results obtained, it was found that λ-cyhalothrin used in the recommended field dose and 10 times lower than recommended significantly increased the metabolic activity of B. bassiana. In relation to the B. brongniartii strain, detlamethrin used in each of the tested concentrations significantly affected its viability.
1. Introduction
Biological methods of plant protection against pests in plant production are one of the alternative methods to chemical agents. They are treated as priority actions in IPM (Integrated Pest Management) as well as in the assumptions of the European Green Deal [1,2,3]. However, their implementation requires extensive knowledge of the ecology, biology, and systematics of harmful organisms and their natural enemies, as well as the impact of the synthetic pesticides used [4,5]. One of the most important directions in the development and implementation of biological methods in plant protection is the use of biopreparations based on bacteria, fungi (entomopathogenic fungi EPF) and viruses [1,6,7,8,9].
Entomopathogenic fungi are microorganisms that are used as bioinsecticides in agriculture, especially organic agriculture. Species from the Beauveria and Metarhizium genera, which are characterized by a very wide spectrum of activity, are often used to produce preparations based on EPF [10,11,12]. The above-mentioned types of EPF not only reduce the number of pests but also endophytically penetrate plant tissues, positively influencing their growth and development, and exhibit properties that allow them to be used in biotechnological processes such as the biosynthesis of nanoparticles [6,10,13,14,15,16]. The main place where EPF occurs is the soil environment and, therefore, they are exposed to contact with pesticides commonly used in plant protection, including pyrethroid insecticides. These microorganisms have developed various adaptive mechanisms that allow them to survive in unfavorable conditions [16,17]. There are 12 EPF strains on the list of active substances approved in the EU. Biopreparations containing EPF are currently used mainly to control the population of thrips, beetles, aphids, whiteflies, and mites in fruit and vegetable crops and ornamental plants [18,19].
Pyrethroids are insecticides commonly used to control arthropods in agriculture, forestry, and greenhouses. They are neurotoxic substances that disrupt the nervous system of insects [16,20,21,22,23]. Due to their widespread use by farmers in plant protection, their residues can be found in all elements of the environment. Despite the fact that pyrethroids are less harmful and safer than other insecticides, it has been proven that they have a negative impact on living organisms [16,24,25].
Studies conducted so far have shown that EPF and pyrethroids can not only be used together to control pests but also coexist in the environment. Therefore, it seems important to not only understand the influence of pyrethroids on the growth of EPF but also on their metabolic activity [16,26,27].
The aim of the study was to determine the effect of different doses of tested pyrethroids on the colony growth and metabolic activity of selected EPF from the Beauveria genus.
2. Materials and Methods
2.1. Fungal Isolates
The effect of preparations containing pyrethroides on EPF colony growth was investigated in laboratory conditions. Two selected species of the Beauveria genus were used in the research: the fungus B. bassiana (Bals.-Criv.) Vuill. (B03-UPH) isolated from meadow soil collected in Klimonty (Mazowieckie Voivodeship, Poland) and B. brongniartii (Sacc.) (B04-UPH) isolated from mid-field woodlots in Boćki (Podlaskie Voivodeship, Poland). The cultures of the fungi used in this experiment were deposited in the fungal collection of the Institute of Agriculture and Horticulture, the University of Siedlce, Poland, and stored in an SDA medium at 4 °C (bioMérieux, Craponne, France). They were identified macroscopically using standard keys [28,29], and their systematic affiliation was also confirmed by molecular identification. The ITS marker, proposed as a universal DNA code marker for fungi, was chosen for identification [30]. Molecular identification of isolates was carried out in the mycological laboratory at the Biological and Chemical Research Center of the University of Warsaw, using Qiagen and Blirt tools (DNA isolation kits, PCR kit and cleaning kit). The PCR reaction was carried out according to the procedure provided by Kovač et al. [31]. Sanger sequencing was used with single ITS2, ITS3, ITS4, and ITS5 primers and the BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Waltham, MA, USA), containing fluorescently labeled dideoxynucleotide triphosphates (ddNTPs), deoxynucleotide triphosphates (dNTPs), and Taq-FS polymerase and buffer. After sequencing, product cleaning was conducted by molecular filtration on columns with Sephadex G-50, and the reading of the result was entrusted to Genomed (Warsaw, Poland). The sequences were compared to those available in the NCBI database using the BLASTN 2.2.2 algorithm [32].
2.2. The Insecticides Used in the Experiment
Three insecticides from the pyrethroid group were used in the laboratory experiment: Decis Mega 50 EW (Bayer SAS, Puteaux, France), (recommended field dose for wheat protection-0.1–0.125 L/ha, active substance, and its content-deltamethrin 50 g/L), Karate Zeon 050 CS (Syngenta, Warsaw, Poland), (recommended field dose for wheat protection-0.1 L/ha, active substance, and its content λ-cyhalothrin 50 g/L), and Cyperkil max 500 EC (Arysta LifeScience Benelux SPRL, Brussels, Belgium) (recommended field dose for wheat protection−0.05 L/ha, active substance, and its content α-cypermethrin-500 g/L). The pyrethroids selected for the experiment are among the most frequently used insecticides from this group to control insect pests in agricultural and horticultural crops in Poland.
The tested preparations were used in three doses:
- A—dose 10 times lower than recommended
- B—recommended field dose
- C—dose 10 times higher than recommended
The selection the above does of pyrethroids (especially A and C) was guided by the fact that the dose of pesticide that actually reaches the insect or soil in the field conditions, may differ from the recommend one. Similar doses of insecticides were tested in laboratory conditions in the works concerning EPF conducted by other authors [16,20,33,34].
In the experiment, Sabouraud dextrose agar (SDA) produced by bioMérieux was used as a culture medium, with casein enzymatic hydrolyzate-5.0 g, hydrolyzed animal tissues-5.0 g, glucose-40 g, and agar-15.0 g. They were all sterilized using a steam-pressure autoclave at 121 °C under a pressure of 1 atmosphere. The tested insecticides were added to the prepared culture medium in the tested doses. They were then transferred to sterile plastic Petri dishes with a diameter of 90 mm.
2.3. Effect on Colony Growth
In the first part of the experiment, the tested fungal isolates were grown on the SDA (bioMérieux, France) medium at 21 ± 1 °C. Before starting the experiment, conidial viability was assessed using the plate count technique on SDA, and viability was >96% and >94% for tested isolates of B. bassiana and B. brongniartii, respectively. A fragment of mycelium from 10-day-old cultures was sampled with a preparation needle, to be inoculated centrally into the solid SDA medium on Petri dishes (90 mm, Noex, Komorniki, Poland) with the determined dose on pyrethroids. The plates with the inoculated isolates were placed in incubators, protected from light, at 21 ± 1 °C. Colony growth observations were carried out every 5 days until day 20, by measuring the colony diameter in mm. The experiment was performed in four repetitions. The control consisted of cultures grown on substrates without the pyrethroids. The results are presented as the size of the colony diameter expressed in mm and in the case of the 20th day of the culture as a percentage in relation to the control.
2.4. Metabolic Activity of Beauveria Strains
Each slant containing the fungal strain was rinsed with 5 mL of SDA (bioMérieux, France) medium and the resulting spore suspension was filtered through a funnel lined with glass wool. The filtrate containing the spores was collected and diluted to a density of 1 × 10⁶ spores/mL. The insecticides were diluted to obtain the following concentrations: A—10 times higher than the recommended dose (concentration A), B—the recommended dose (concentration B), and C—10 times lower than the recommended dose (concentration C). An amount of 100 μL of SDA medium with the test insecticides at concentrations A, B, and C were added to each well of a microtiter plate. Then, 100 μL of the prepared spore suspension was added to each well. At the same time, wells containing biotic samples were prepared according to the scheme above, where 100 μL of fresh SDA medium was added instead of 100 μL of insecticides. All plates were incubated at room temperature for 72 h in the dark. After incubation, 100 μL of freshly prepared fluorescein diacetate solution (2 mg/mL in 0.1 M phosphate buffer, pH 6.4), (Sigma-Aldrich, Taufkirchen, Germany) was added to each well. The plates were then incubated in the dark for 30 min at room temperature. The experiment was performed in four repetitions. Fluorescence was measured at an excitation wavelength of 485 nm and an emission wavelength of 520 nm using an OMEGA spectrofluorimetric reader (BMG Labtech, Offenburg, Germany). The final results were expressed as percentages, with the fluorescence intensity of the biotic control set to 100%. For each tested sample, the fluorescence intensity was calculated relative to this control. Values below 100% indicated a decrease in the measured parameter, while values above 100% indicated an increase.
2.5. Statistical Analyses
The results obtained were statistically processed using the Statistica program v. 13.3 (TIBCO Software Inc., Palo Alto, CA, USA). The first stage of the analysis was to check with the chi-square λ2 test whether the distribution of the tested trait (colony growth) in the sample followed a normal distribution. Since the data did not have a normal distribution, the transformation y = log (x + 0.5) was applied. Then, on the transformed data, one-way ANOVA analysis was performed for each factor separately according to the following model:
where
yij = m + ai + eij
- -
- m is the population average;
- -
- yij is the value of the examined trait (colony growth);
- -
- ai is the effect of the ith level of factor A (fungal species, dose of preparation, observation time); and
- -
- eij is the random error.
When the factor’s effect was significant, the Tukey test was used at α = 0.05 to compare the means (posthoc analysis).
The metabolic activity was analyzed for normality using the Shapiro–Wilk test, and significant differences between treatment groups were determined using a one-way ANOVA followed by Dunnett’s test. The standard deviation was calculated (SD).
3. Results
3.1. Effect on Colony Growth of EPF
The pyrethroid insecticides used in the experiment showed various toxic effects towards the tested EPF of the genus Beauveria (Table 1 and Table 2). Deltamethrin and α-cypermethrin in a dose 10 times higher than recommended field dose (C) on the 5th day of the observation completely inhibited the growth of the B. bassiana fungus (Table 1). On days 15 and 20 of observation, α-cypermethrin used in dose C statistically significantly (p < 0.012) limited the growth of B. bassiana colonies. Among the tested pyrethroids, λ-cyhalothrin used in the recommended field dose (B) at each of the observation dates limited the growth of fungal colonies to the least extent compared to the control, and this difference was statistically significant (p < 0.013). On the 20th day of the observation, at a dose 10 times lower than the recommended field dose (A), deltamethrin had the weakest effect on the growth of B. bassiana among the tested pyrethroids. This difference was statistically significant (p < 0.011) in relation to the control and the other tested doses of the preparation. Karate Zeon 050 CS, containing α-cypermethrin, at dose C showed statistically the strongest (p < 0.021) inhibitory effect on the growth of colonies of the tested EPF species. Among the tested pyrethroids, the use of λ-cyhalothrin at the recommended field dose (B) limited the growth of B. brongniartii colonies the least (Table 2).

Table 1.
Diameters of B. bassiana fungus colonies during cultivation on media with the addition of selected insecticides (expressed in mm).
Table 2.
Diameters of B. brongniartii fungus colonies during cultivation on media with the addition of selected pyrethroids (expressed in mm).
The use of pyrethroids at a dose 10 times lower than the recommended field dose (A) limited the growth of B. brongniartii colonies the least, and in the case of λ-cyhalothrin, a slight effect of stimulating colony growth was observed.
Comparing the growth of the tested EPFs from the Beauveria genus on the 20th day of observation, at the recommended field dose of pyrethroids (B), it was found that deltamethrin had a stronger inhibitory effect on the fungus B. brongniartii than on B. bassiana (Figure 1). The B. bassiana fungus showed statistically significant (p < 0.013) greater sensitivity to the presence of λ-cyhalothrin and α-cypermethrin in the culture medium than B. brongniartii.
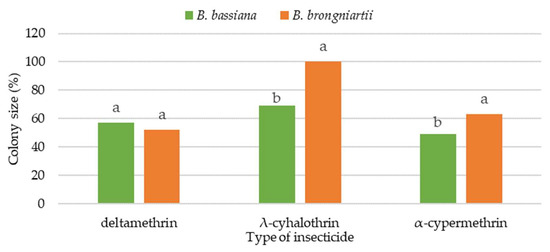
Figure 1.
Colony size B. bassiana and B. brongniartii on the 20th day of growth on media with recommended field dose (B) of tested pyrethroids (% in relation to the control); a,b—means within columns with the same lowercase letters are not significant at α = 0.05.
The λ-cyhalothrin used in the recommended field dose did not show any inhibitory effect on the fungus B. brongniartii. In relation to both tested EPFs, the pyrethroid λ-cyhalothrin had the least toxic effect, and the fungal colonies reached the largest sizes.
3.2. Insecticides Effects on Metabolic Activity of EPF
The metabolic activity of B. bassiana varied depending on the insecticide and concentration tested (Figure 2). Deltamethrin, at both the recommended field dose and 10 times that concentration, inhibited metabolic activity. The α-cypermethrin exhibited the strongest inhibitory effect, almost completely suppressing metabolic activity. Conversely, λ-cyhalothrin demonstrated a more complex effect. At the recommended field dose and 10 times lower than the recommended dose, λ-cyhalothrin increased B. bassiana metabolic activity by approximately 60%. However, a tenfold increase above the recommended dose significantly inhibited metabolic activity. Only a dose 10 times lower than the recommended field dose of deltamethrin resulted in no significant statistical difference (p < 0.068) in the metabolic activity of the tested strain compared to the control.
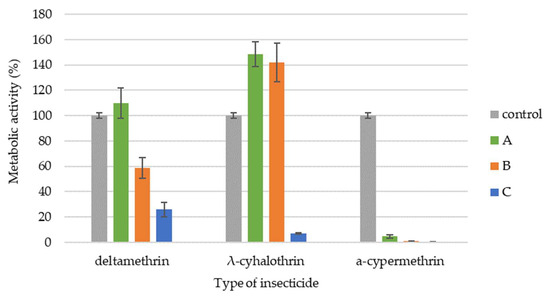
Figure 2.
Metabolic activity of B. bassiana exposed to different doses of the tested pyrethroids (A—10-fold lower than the recommended field dose; B—recommended field dose; C—10-fold higher than the recommended field dose). Statistically significant differences within each treatment group, compared to the control, were determined using one-way ANOVA followed by Dunnett’s test.
The pyrethroids exhibited varying effects on the metabolism of B. brongniarti (Figure 3). Deltamethrin significantly increased metabolic activity across all concentrations. The λ-cyhalothrin only enhanced metabolic activity at the recommended field dose while other tested concentrations of the insecticides had no statistically significant (p < 0.071) effect on the viability of B. brongniartii strain. The α-cypermethrin had a contrasting effect on B. brongniartii metabolism. While the recommended field dose and a ten-fold higher concentration inhibited metabolic activity, the concentration 10 times lower than recommended enhanced it.
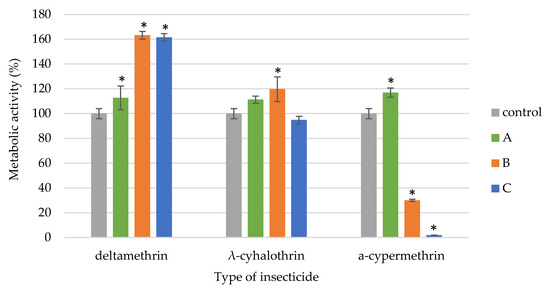
Figure 3.
Metabolic activity of B. brongniartii exposed to different doses of the tested pyrethroids (A—10-fold lower than the recommended field dose; B—recommended field dose; C—10-fold higher than the recommended field dose). Statistically significant differences within each treatment group, compared to the control, were determined using one-way ANOVA followed by Dunnett’s test and are indicated by * (p < 0.05).
4. Discussion
One of the main factors limiting the use of biopesticides containing entomopathogenic fungi in plant protection is the impact of chemical pesticides on their pathogenicity. Most of laboratory research indicates a negative effect of pesticides on EPF [6,15,35,36,37,38,39]. Pyrethroids are pyrethrin analogs that affect the nervous system of insects by interfering with the activity of chloride and sodium channels [25,40]. They are used in agriculture, forestry, and urban areas and show higher efficacy and lower toxicity compared to other insecticides [24].
Based on the conducted research, it was found that pyrethroids added to the culture medium had a different impact on the growth of the tested EPF of the Beauveria genus. The effect of pyrethroid insecticides on the development of colonies of the tested fungi depended on the type of active substance used, the concentration of pyrethroid in the culture medium, and the fungus species. In vitro studies conducted so far have shown that individual fungal species, and sometimes even individual strains within a species, are characterized by varying sensitivity to plant protection products [6,16,27,41,42]. Studies show that entomopathogens introduced into the environment (interchangeably with synthetic insecticides) as part of IPM can maintain their activity in the soil environment for many years, effectively reducing the pest population [43].
When interpreting the results obtained, it should be taken into account that tests were conducted in laboratory conditions which do not always correspond to the situation in the field both in direct contact with the sprayed insects and in the soil environment. The overall actual effect of the pesticides tested on the populations and performance of insect pathogenic fungi in the field is difficult to evaluate. It is certain that most of the time the concentrations of the chemicals that reach the fungi in the soil are smaller than those employed in our experiment. Locally, however, the concentrations may be much higher and exceed the normal ones. This may be especially true for pesticides that are applied directly into the soil. On the other hand, fungi growing on agar in Petri dishes are grown under optimum conditions, which may make them more tolerant to the chemicals than in nature where suboptimal growing conditions, many different antagonists and adverse weather conditions prevail. In such situations even a small additional stress factor may significantly lower their fitness and performance. On the other hand, pesticides are affected and degraded by different environmental factors on the plant and in the soil [41,44].
In the literature, the results of studies conducted so far indicate a negative impact of insecticides on EPF used to control crop pests. Amutha et al. [38] in their study showed that profenofos and methyldemeton inhibited the growth of B. bassiana by 93.72% and 95.17% after 14 days of inoculation and by 83.55% and 93.86% after 30 days. The addition of indoxacarb to the culture medium inhibited the growth of B. bassiana colonies by 100% after 14 and 30 days of inoculation. Forlani et al. [26] in their study showed that deltamethrin used at a concentration of 40 lg cm−2 caused a significant decrease in the number of colonies forming units of B. bassiana. Litwin et al. [16] found that pyrethroids used at recommended (and higher) doses strongly inhibited the development of B. bassiana during the initial growth period, reducing the rate of colony formation by more than 40%, whereas at lower doses, these insecticides had minimal inhibitory effect. Bednarek et al. [45] in their study found that the insecticides Karbosulfan inhibited, while Karbofuran stimulated the growth of B. brongniartii fungus colonies. Widyaningsih et al. [39] studied the effect of Profenofos pesticide and λ-cyhalothrin on the growth of EPF isolated from citrus orchard and found that all EPF (Hirsutella sp., Paecilomyces sp., B. bassiana and M. anisopliae) used in this experiment did not show significant growth reduction after treatment with different doses of λ-cyhalothrin pesticide. The use of the tested pyrethroids in higher concentrations can significantly limit the growth of EPF isolates [6,39]. Srinivasulu and Ortiz [46] in their studies showed that with the use of higher concentrations of pesticides, the fungal population gradually decreased and reached a minimum concentration of 10 kg ha−1. This suggests that the concentration of pesticides used in the field or in the substrate must be controlled. In contrast, studies conducted by James and Elzen [47], Bhattacharya et al. [48], and Kumar et al. [49] showed that B. bassiana strains were highly compatible with the insecticides imidacloprid and spinosad, showing no inhibition of growth, sporogenisation, or viability.
Our own studies showed that the metabolic activity of B. bassiana and B. brongniartii varied depending on the insecticide and the tested concentration. Studies conducted so far indicate that insecticides commonly used in agricultural production not only limit growth but also cause abnormalities in cellular metabolic pathways and damage EPF cell structures [16,25,50,51,52,53]. Litwin et al. [16] showed that the tested pyrethroids did not show a negative effect on the metabolic activity of B. bassiana, and even a slight increase in activity was found in samples exposed to the tested substances. In 24 h cultures, a 1.47-fold increase in metabolic activity was noted in the presence of deltamethrin, a 1.36-fold increase in the presence of λ-cyhalothrin, and a 1.32-fold increase in the presence of α-cypermethrin. In 36 h cultures, a 1.1-fold increase in metabolic activity was observed in the presence of λ-cyhalothrin, while in the presence of α-cypermethrin and deltamethrin, the metabolic activity in the samples was comparable to that observed in the control. In 48 h cultures, no differences in metabolic activity were found between cultures exposed to pyrethroids and biotic control. Różalska et al. [50] examined the metabolic activity of Metarhizium robertsii and found that the addition of both 4-n-nonylphenol and technical nonylphenol caused a statistically significant decrease in the metabolic activity of the tested strain. Litwin et al. [25] found that the soil EPF B. bassiana ARSEF 2860 accumulated pyrethroids as early as day 2 of incubation with an average efficiency of 90%. Pyrethroids accumulating in large amounts in the mycelium of B. bassiana induced oxidative stress and interacted differently with enzymes of basic metabolic pathways. This study showed that accumulated pyrethroids reduced phospholipase C activity, which increased the triacylglycerol to diacylglycerol (TAG/DAG) ratio, especially in the mycelium in which α-cypermethrin had accumulated.
5. Conclusions
Entomopathogenic fungi have a broad spectrum of action and can infect a wide range of arthropod species including crop pests. Research shows that in addition to the use of EPF in biocontrol, these fungi can also be used for removing harmful substances from the environment, because they possess enzymes that enable the removal of toxic compounds of anthropogenic origin. The results of our research confirm that the use of pyrethroids affects the colony growth and metabolic activity of the fungi B. bassiana and B. brongniartii. The studies conducted showed that on the 20th day of the observation, λ-cyhalothrin used in the recommended field dose limited the growth of B. bassiana to the least extent in relation to the other tested pyrethroids. However, with respect to the fungus B. brongniartii, no toxic effect of this pyrethroid was found, and even a slight stimulation of its growth was observed at a dose 10 times lower than the recommended one. Based on the results obtained, it was found that λ-cyhalothrin, used in the recommended field dose and 10 times lower than recommended, significantly increased the metabolic activity of B. bassiana. In relation to the B. brongniartii strain, deltamethrin used in each of the tested concentrations significantly affected its viability. Regardless of the pyrethroid used, at a dose 10 times higher than the recommended field dose, it was found that the B. brongniartii fungus showed greater viability compared to the B. bassiana strain. This indicates that the tested B. brongniartii strain can potentially be used interchangeably with synthetic pyrethroids. Data obtained in this study may guide future recommendations of use pyrethroids in IPM programs where B. bassiana and B. brongniartii are intended to be used as a biocontrol agent of insect and mite pests in agricultural and horticultural crops.
Author Contributions
A.M.-S. conceived and designed the experiments. A.M.-S., C.T., S.R. and M.N. conducted the experiments. A.M.-S., C.T. and S.R. analyzed the data. A.M.-S., C.T. and S.R. contributed to writing and editing of the manuscript; writing—original draft, A.M.-S. and S.R.; writing—review and editing, A.M.-S. and C.T.; supervision, A.M.-S. All authors have read and agreed to the published version of the manuscript.
Funding
The results of the research carried out under research theme No. 159/23/B were financed by a science grant from the Ministry of Science and Higher Education.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sosnowska, D. The contribution of conservation biological control method to integrated plant protection and organic farming. Prog. Plant Prot. 2018, 58, 288–293. [Google Scholar] [CrossRef]
- Ginter, A. Small Farms Facing the Challenges of Sustainability and the Green Deal; Scientific Dissertation; UPH: Siedlce, Poland, 2021; ISBN 978-83-66541-76-4. [Google Scholar]
- Majchrowska-Safaryan, A.; Tkaczuk, C.; Wrzosek, M. The effect of humic substances on the colony growth and conidial germination of entomopathogenic fungi from the genus Metarhizium. Sustainability 2024, 16, 3616. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Chukwuneme, C.F.; Samuel, O.; Olanrewaju, M.A.; Adegboyega, T.T.; Babalola, O.O. Contribution of microbial inoculants in sustainable maintenance of human health, including test methods and evaluation of safety of microbial pesticide microorganisms. In Botanicals and Microorganisms for Improving Agriculture and Human Health; Logos Verlag Berlin GmbH: Berlin, Germany, 2021; Volume 1, pp. 207–240. [Google Scholar]
- Ayilara, M.S.; Adeleke, B.S.; Babalola, O.O. Bioprospecting and challenges of plant microbiome research for sustainable agriculture, a review on soybean endophytic bacteria. Microb. Ecol. 2022, 85, 1113–1135. [Google Scholar] [CrossRef]
- Tkaczuk, C. Occurrence and Infective Potential of Entomopathigenic Fungi in Soils of Agrocenoses and Seminatural Habitats in the Agricultural Landscape; Scientific Dissertation No. 94; AP: Siedlce, Poland, 2008. [Google Scholar]
- Tomalak, M. Market for biological control agents and their legal regulation. Prog. Plant Prot. 2010, 50, 1053–1063. [Google Scholar]
- Fenibo, E.O.; Grace, N.I.; Weiz, N.; Tonderayi, M. The potential and green chemistry attributes of biopesticides for sustainable agriculture. Sustainability 2022, 14, 14417. [Google Scholar] [CrossRef]
- Šunjka, D.; Špela, M. An Alternative Source of Biopesticides and Improvement in Their Formulation—Recent Advances. Plants 2022, 11, 3172. [Google Scholar] [CrossRef] [PubMed]
- Rıos-Moreno, A.; Garrido-Jurado, I.; Resquın-Romero, G.; Arroyo-Manzanares, N.; Arce, L.; Quesada-Morenga, E. Destruxin A production by Metarhizium brunneum strains during transient endophytic colonisation of Solanum tuberosum. Biocontrol Sci. Technol. 2016, 26, 1574–1585. [Google Scholar] [CrossRef]
- Gao, B.J.; Mou, Y.N.; Tong, S.M. Subtilisin-like Pr1 proteases marking the evolution of pathogenicity in a wide-spectrum insect-pathogenic fungus. Virulence 2020, 11, 365–380. [Google Scholar] [CrossRef]
- Holka, M.; Kowalska, J. The potential of adjuvants used with microbiological control of insect pests with emphasis on organic farming. Agriculture 2023, 13, 1659. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 117. [Google Scholar] [CrossRef]
- Różalska, S.; Soliwoda, K.; Długoński, J. Synthesis of silver nanoparticles from Metarhizium robertsii waste biomass extract after nonylphenol degradation, and their antimicrobial and catalytic potential. RSC Adv. 2016, 6, 21475–21485. [Google Scholar] [CrossRef]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Litwin, A.; Bernat, P.; Nowak, M.; Słaba, M.; Różalska, S. Lipidomic response of the entomopathogenic fungus Beauveria bassiana to pyrethroids. Sci. Rep. 2021, 11, 21319. [Google Scholar] [CrossRef]
- Nowak, M.; Soboń, A.; Litwin, A.; Rożalska, S. 4-nnonylphenol degradation by the genus Metarhizium with cytochrome P450 involvement. Chemosphere 2019, 220, 324–334. [Google Scholar] [CrossRef]
- Dara, S.K. Entomopathogenic Microorganisms: Modes of Action and Role in IPM. Strawberries and Vegetables; ANR Blogs: Davis, CA, USA, 2017; pp. 1–7. [Google Scholar]
- Arthurs, S.; Dara, S.K. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Tkaczuk, C.; Krzyczkowski, T.; Głuszczak, B.; Król, A. The influence of selected pesticides on the colony growth and conidial germination of the entomopathogenic fungus Beauveria bassiana (Bals.) Vuill. Prog. Plant Prot. 2012, 52, 969–974. [Google Scholar]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-degrading microorganisms and their potential for the bioremediation of contaminated soils: A review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef]
- Tkaczuk, C.; Majchrowska-Safaryan, A.; Głuszczak, B. The effect of selected pesticides approved for use in organic farming on the growth of entomopathogenic fungi. Prog. Plant Prot. 2016, 56, 265–271. [Google Scholar]
- Skolarczyk, J.; Pekar, J.; Nieradko-Iwanicka, B. Immune disorders induced by exposure to pyrethroid insecticides. Postep. Hig. Med. Dosw. 2017, 71, 446–453. [Google Scholar] [CrossRef]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid pesticide residues in the global environment: An overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef]
- Litwin, A.; Mironenka, J.; Bernat, P.; Soboń, A.; Różalska, S. Accumulation of pyrethroids induces changes in metabolism of the entomopathogenic fungus Beauveria bassiana—Proteomic and lipidomic background. Ecotoxicol. Environ. Saf. 2023, 249, 114418. [Google Scholar] [CrossRef] [PubMed]
- Forlani, L.; Juarez, M.P.; Lavarías, S.; Pedrini, N. Toxicological and biochemical response of the entomopathogenic fungus Beauveria bassiana after exposure to deltamethrin. Pest Manag. Sci. 2014, 70, 751–756. [Google Scholar] [CrossRef]
- Mayerhofer, J.; Enkerli, J.; Zelger, R.; Strasser, H. Biological control of the European cockchafer: Persistence of Beauveria brongniartii after long-term applications in the Euroregion Tyrol. BioControl 2015, 60, 617–629. [Google Scholar] [CrossRef]
- Samson, R.A.; Evans, H.C. Two new Beauveria spp. from South America. J. Invertebr. Pathol. 1982, 39, 93–97. [Google Scholar] [CrossRef]
- Humber, R.A. Manual of Techniques in Invertebrate Pathology. In Identification of Entomopathogenic Fung; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 151–187. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of entomopathogenic fungi as naturally occurring enemies of the invasive oak lace bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new 432 generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Miętkiewski, R.T.; Pell, J.; Clark, S.J. Influence of pesticides use on the natural occurrence of etomopathogenic fungi in arable soils in the UK: Field and laboratory comparisons. Biocontrol Sci. Technol. 1997, 7, 565–575. [Google Scholar] [CrossRef]
- Rashid, M.; Baghdadi, A.; Sheikhi, A.; Pourian, H.R.; Gacavi, M. Compatability of Metarhizium anisopilae (Ascomycota: Hypocreales) with several insceticides. J. Plant Prot. Res. 2010, 50, 22–27. [Google Scholar] [CrossRef]
- Miętkiewski, R.; Miętkiewska, Z.; Sapieha, A.; Badowska-Czubik, T. Wpływ herbicydów na patogeniczność grzybów entomopatogennych. Zesz. Naukowe. Ser. Rolnictwo. Wyższa Szk. Rol. Pedagog. Siedlcach 1995, 37, 179–184. [Google Scholar]
- Miętkiewski, R.; Ignatowicz, S.; Górski, R. Porównanie wpływu BioNEMTM, insektycydu pochodzenia roślinnego i wybranych insektycydów syntetycznych na wzrost grzybni grzybów owadobójczych. Pestycydy 1996, 1, 15–27. [Google Scholar]
- Tkaczuk, C. Wpływ wybranych pestycydów stosowanych w ochronie sadów na wzrost grzybów owadobójczych. Biul. Nauk. 2001, 12, 375–383. (In Polish) [Google Scholar]
- Amutha, M.; Banu, G.J.; Surulivelu, T.; Gopalakrishnan, N. Effect of commonly used insecticides on the growth of white Muscardine fungus, Beauveria bassiana under laboratory conditions. J. Biopest. 2010, 3, 143–146. [Google Scholar]
- Widyaningsih, S.; Triasih, H.U.; Agustina, D. Effect pesticides to entomopathogen fungi from citrus orchard in vitro. IOP Conf. Ser. Earth Environ. Sci. 2021, 803, 012021. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhan, H.; Chen, S. Insight into microbial applications for the biodegradation of pyrethroid insecticides. Front. Microbiol. 2019, 10, 1778. [Google Scholar] [CrossRef]
- Vänninen, I.; Hokkanen, H. Effect of pesticides on four species of entomopathogenic fungi in vitro. Ann. Agric. Fenn. 1998, 27, 345–353. [Google Scholar]
- Bajan, C.; Kmitowa, K. Thirty year studies on entomopathogenic fungi in the Institute of Ecology, PAS. Pol. Ecol. Stud. 1997, 23, 133–154. [Google Scholar]
- Mayerhofer, J.; Lutz, A.; Franco Widmer, F.; Rehner, S.A.; Leuchtmann, A.; Enkerli, J. Multiplexed microsatellite markers for seven Metarhizium species. J. Invertebr. Pathol. 2015, 132, 132–134. [Google Scholar] [CrossRef]
- Clark, R.A.; Casagrande, R.A.; Wallace, D.B. Influence of pesticides on Beauveria bassiana, a pathogen of the Colorado potato beetle. Environ. Entomol. 1982, 11, 67–70. [Google Scholar] [CrossRef]
- Bednarek, A.; Popowska-Nowak, E.; Pezowicz, E.; Kamionek, M. Integrated methods in pest control: Effect of insecticides on entomopathogenic fungi [Beauveria bassiana [Bals] Vuill., B. brongniartii [Sacc.]] and nematodes [Heterorhabditis megidis Poinar, Jackson, Klein, Steinernema feltiae Filipjev, S.Glaseri Steiner]. Pol. J. Ecol. 2004, 52, 223–228. [Google Scholar]
- Srinivasulu, M.; Ortiz, D.R. Effect of pesticides on bacterial and fungal populations in ecuadorian tomato cultivated soils. Environ. Process. 2017, 4, 93–105. [Google Scholar] [CrossRef]
- James, R.R.; Elzen, G.W. Anatgonism between Beauveria bassiana and imidachloprid when combined for Bemisia argentifolia (Homoptera: Aleuyrodidae) control. J. Econ. Entomol. 2001, 94, 357–361. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Dutta, S.; Dhar, T. In vitro compatibility of different entomopathogens to pesticides, plant growth regulators and micronutrients. Ann. Plant Prot. Sci. 2004, 12, 199–202. [Google Scholar]
- Kumar, S.S.; Monga, D.; Kumar, R.; Nagrale, D.T.; Hiremani, N.S.; Kranthi, S. Compatibility of entomopathogenic fungi with insecticides and their efficacy for IPM of Bemisia tabaci in cotton. J. Pestic. Sci. 2019, 44, 97–105. [Google Scholar] [CrossRef]
- Różalska, S.; Glińska, S.; Długoński, J. Metarhizium robertsii morphological fexibility during nonylphenol removal. Int. Biodeterior. Biodegrad. 2014, 95, 285–293. [Google Scholar] [CrossRef]
- Szewczyk, R.; Soboń, A.; Słaba, M.; Długoński, J. Mechanism study of alachlor biodegradation by Paecilomyces marquandii with proteomic and metabolomic methods. J. Hazard. Mater. 2015, 291, 52–64. [Google Scholar] [CrossRef]
- Stolarek, P.; Różalska, S.; Bernat, P. Lipidomic adaptations of the Metarhizium robertsii strain in response to the presence of butyltin compounds. Biochim. Biophys. Acta Biomembr. 2019, 1861, 316–326. [Google Scholar] [CrossRef]
- Subbanna, A.R.N.S.; Chandrashekara, C.; Stanley, J.; Mishra, K.K.; Mishra, P.K.; Pattanayak, A. Bio-efficacy of chitinolytic Bacillus thuringiensis isolates native to northwestern Indian Himalayas and their synergistic toxicity with selected insecticides. Pestic. Biochem. Physiol. 2019, 158, 166–174. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).