
Effects of Colony Breeding System and Nest Architecture on Soil Microbiome and Fertility in the Fungus-Growing Termite Macrotermes barneyi Light
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Soil Physicochemical Analysis
2.3. Metagenome DNA Extraction and Shotgun Sequencing
2.4. Metagenomics Analysis
2.5. Statistical Analysis
3. Results
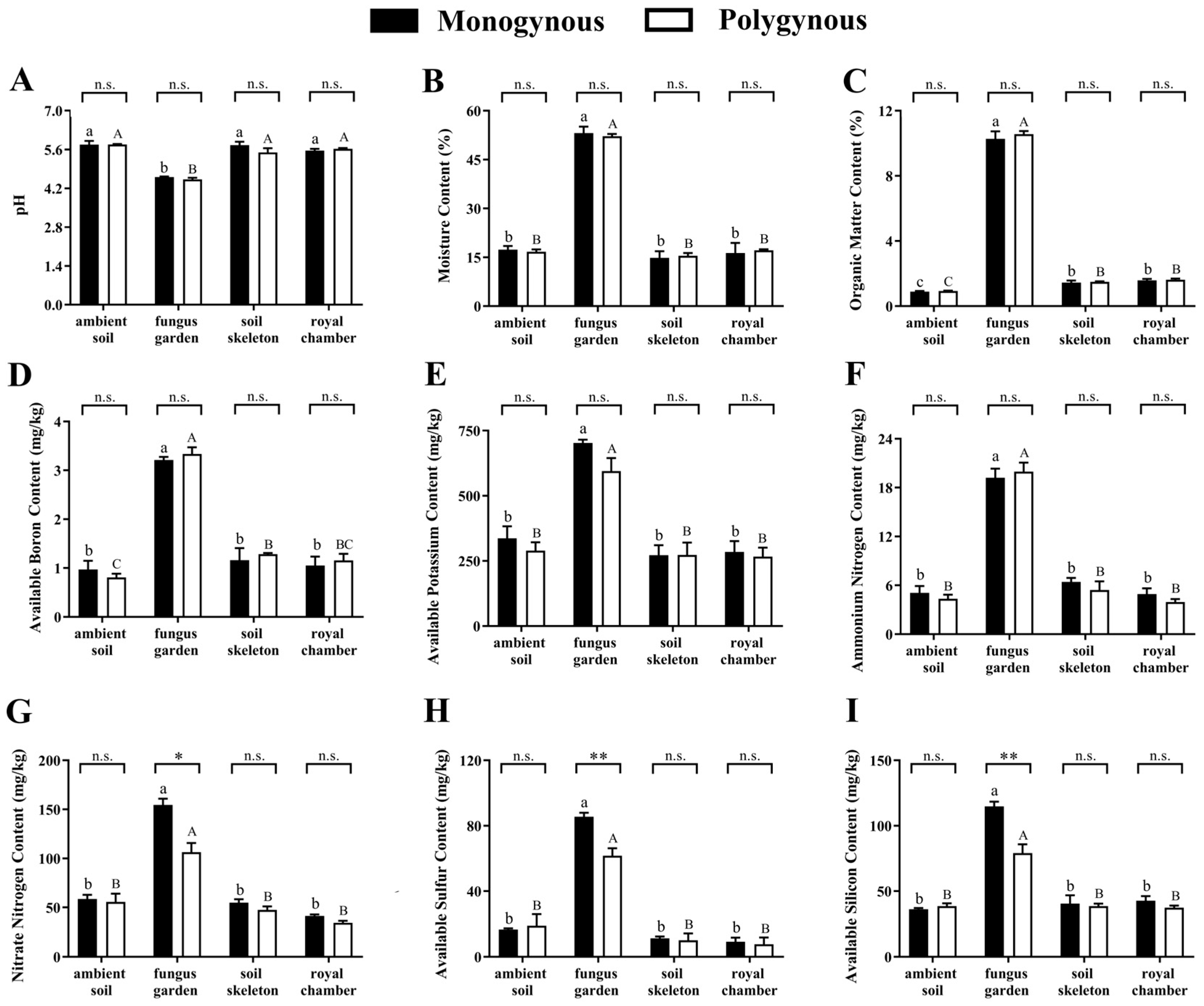
3.1. Soil Physicochemical Characterization
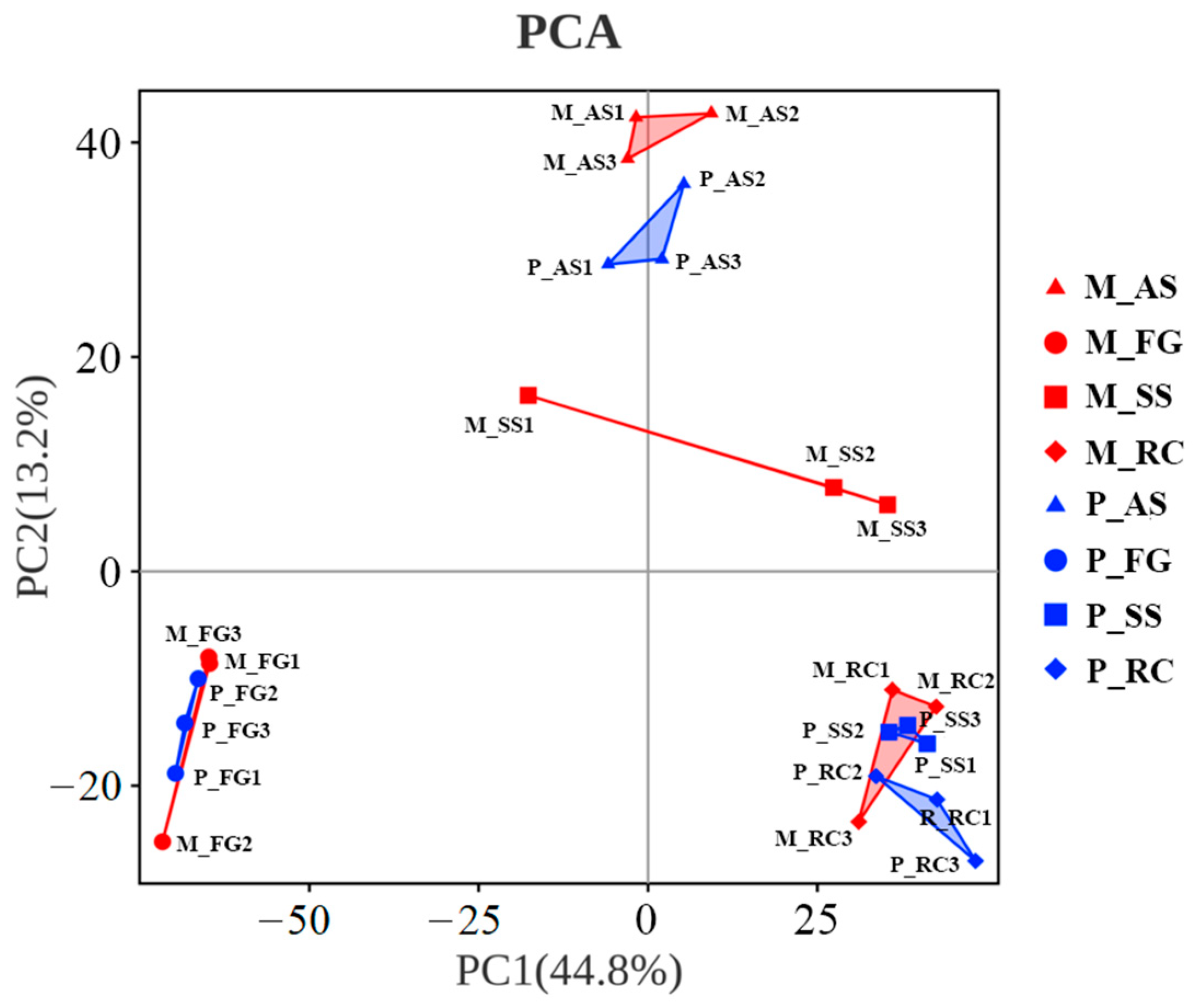
3.2. Microbial Community Composition
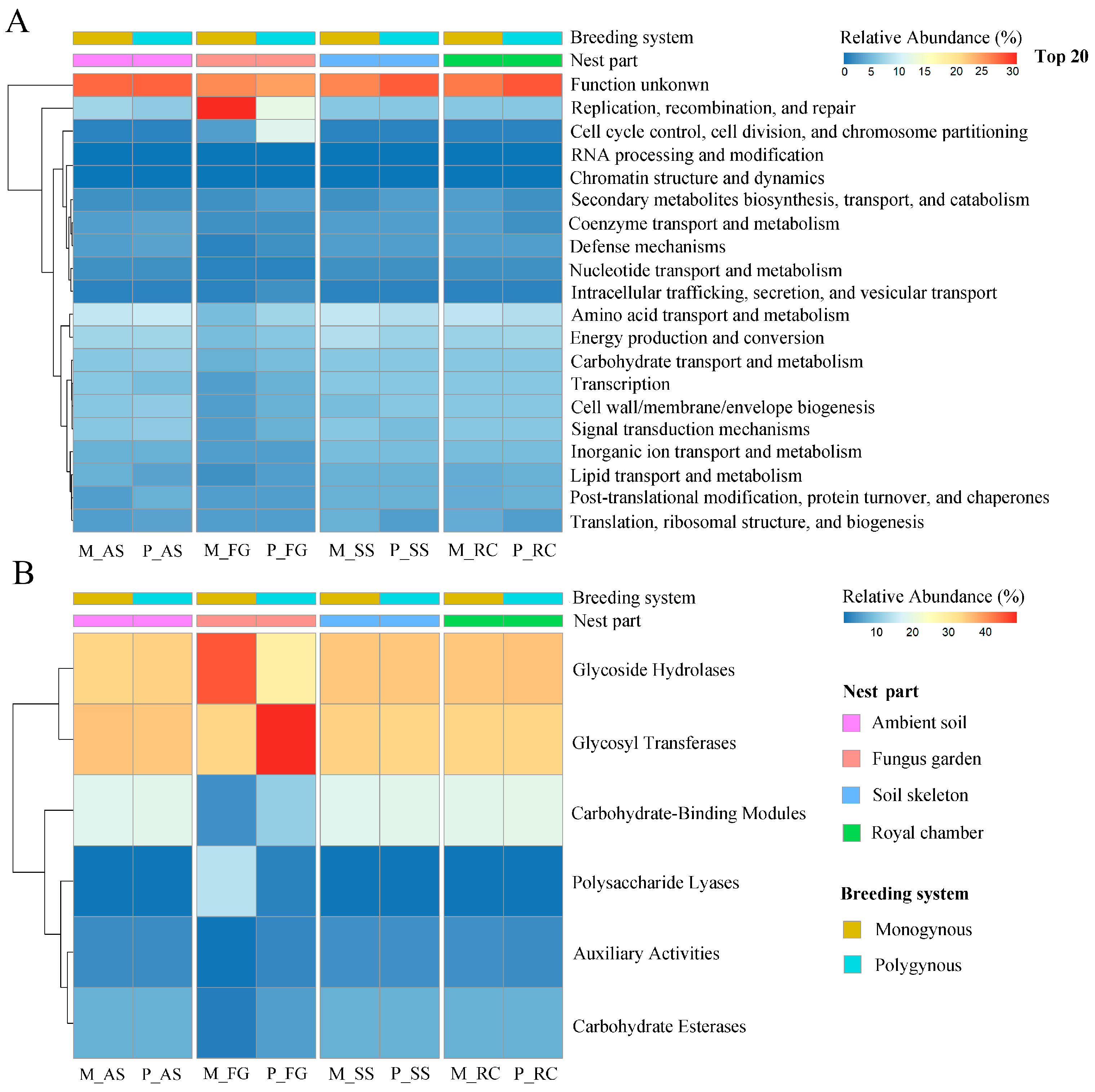
3.3. Functional Annotation of eggNOG
3.4. Functional Annotation of CAZys
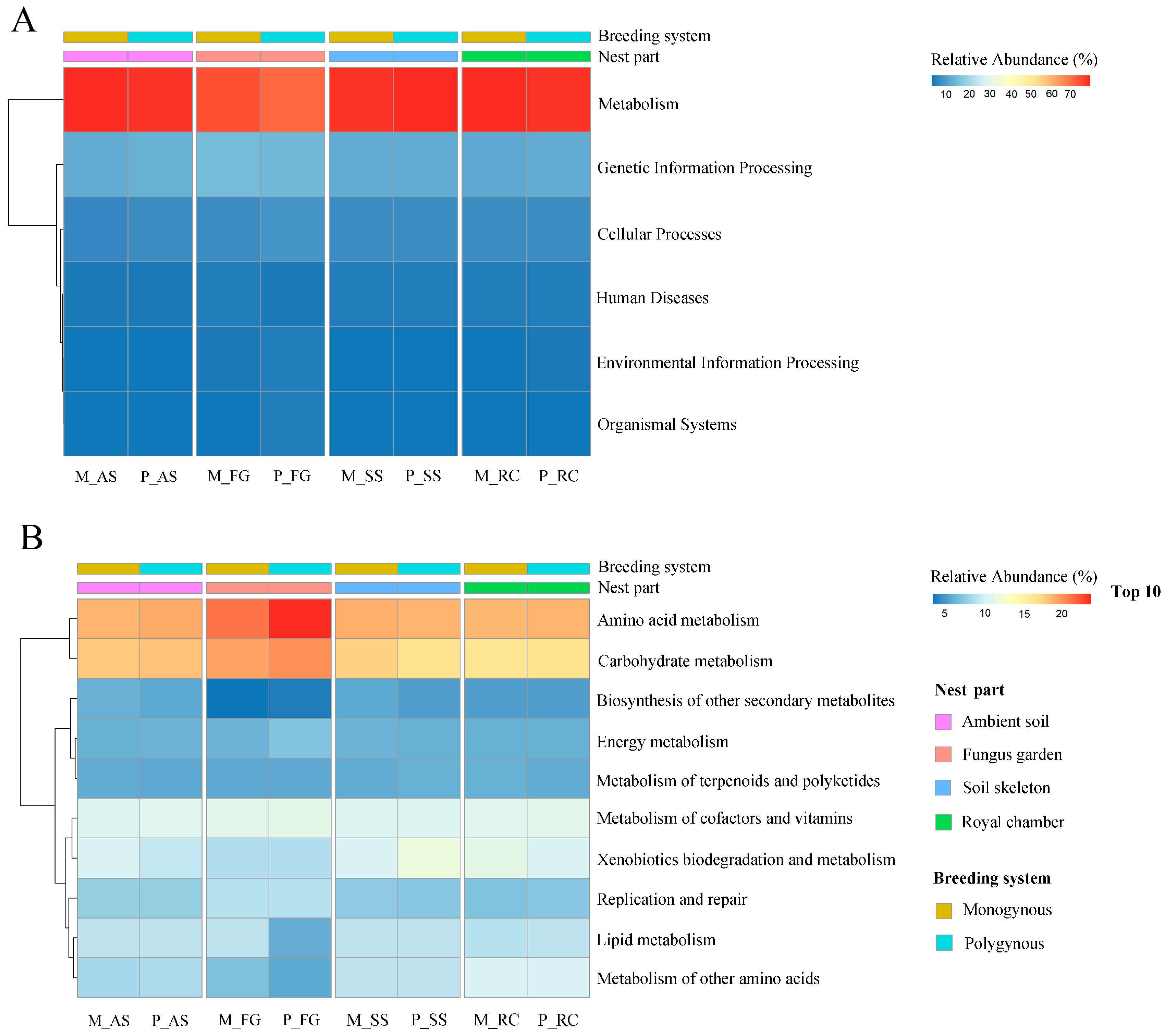
3.5. Functional Annotation of KEGG
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lax, A.R.; La Osbrink, W. United states department of agriculture-agriculture research service research on targeted management of the formosan subterranean termite Coptotermes formosanus Shiraki (Isoptera: Rhinotermitidae). Pest Manag. Sci. 2003, 59, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Su, N.Y. Managing social insects of urban importance. Annu. Rev. Entomol. 2012, 57, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J. Estimation of the damage risk range and activity period of termites (Reticulitermes speratus) in Korean wooden architectural heritage building sites. Forests 2024, 15, 602. [Google Scholar] [CrossRef]
- Wu, D.H.; Seibold, S.; Ellwood, M.D.F.; Chu, C.J. Differential effects of vegetation and climate on termite diversity and damage. J. Appl. Ecol. 2022, 59, 2922–2935. [Google Scholar] [CrossRef]
- Xu, X.X.; Zeng, Q.S.; Li, D.; Wu, J.; Wu, X.G.; Shen, J.Y. GPR detection of several common subsurface voids inside dikes and dams. Eng. Geol. 2010, 111, 31–42. [Google Scholar] [CrossRef]
- Holt, J.A.; Lepage, M. Termites and soil properties. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 389–407. [Google Scholar]
- Jouquet, P.; Traore, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Bignell, D.E.; Eggleton, P. Termites in ecosystems. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 363–387. [Google Scholar]
- Joseph, G.S.; Seymour, C.L.; Cumming, G.S.; Cumming, D.H.M.; Mahlangu, Z. Termite mounds increase functional diversity of woody plants in african savannas. Ecosystems 2014, 17, 808–819. [Google Scholar] [CrossRef]
- Jouquet, P.; Guilleux, N.; Shanbhag, R.R.; Subramanian, S. Influence of soil type on the properties of termite mound nests in Southern India. Appl. Soil Ecol. 2015, 96, 282–287. [Google Scholar] [CrossRef]
- Juergens, N. The biological underpinnings of namib desert fairy circles. Science 2013, 339, 1618–1621. [Google Scholar] [CrossRef]
- Jurgensen, M.F.; Miller, C.A.; Trettin, C.T.; Page-Dumroese, D.S. Bedding of wetland soil: Effects of bed height and termite activity on wood decomposition. Soil Sci. Soc. Am. J. 2019, 83, S218–S227. [Google Scholar] [CrossRef]
- Li, H.; Greening, C. Termite-engineered microbial communities of termite nest structures: A new dimension to the extended phenotype. FEMS Microbiol. Rev. 2022, 46, fuac034. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Vargo, E.L.; Kawatsu, K.; Labadie, P.E.; Nakano, H.; Yashiro, T.; Tsuji, K. Queen succession through asexual reproduction in termites. Science 2009, 323, 1687. [Google Scholar] [CrossRef]
- Adams, E.S.; Atkinson, L. Queen fecundity and reproductive skew in the termite Nasutitermes corniger. Insect Soc. 2008, 55, 28–36. [Google Scholar] [CrossRef]
- Reber, A.; Castella, G.; Christe, P.; Chapuisat, M. Experimentally increased group diversity improves disease resistance in an ant species. Ecol. Lett. 2008, 11, 682–689. [Google Scholar] [CrossRef]
- Da Silva, L.H.B.; Jost, C.; Vargo, E.L.; Costa-Leonardo, A.M.; Haifig, I. Incipient colonies of the neotropical termite Cornitermes cumulans (Isoptera: Termitidae): Comparing monogamy and polygamy as reproductive strategies. Insect. Soc. 2022, 69, 99–104. [Google Scholar] [CrossRef]
- Sun, P.; Li, G.; Jian, J.; Liu, L.; Chen, J.; Yu, S.; Xu, H.; Lei, C.; Zhou, X.; Huang, Q. Transcriptomic and functional analyses of phenotypic plasticity in a higher termite, Macrotermes barneyi Light. Front. Genet. 2019, 10, 964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lin, Z.J.; Huang, Q.H.; Shen, Y.L.; Ni, J.F. First record of gregarine protists (Apicomplexa: Sporozoa) in Asian fungus-growing termite Macrotermes barneyi (Blattaria: Termitidae). Sci. Rep. 2021, 11, 989. [Google Scholar] [CrossRef]
- Zhong, J.H.; Liu, L.L. Termite fauna in China and their economic importance. Sociobiology 2002, 40, 25–32. [Google Scholar]
- Chen, F.Y. Colony fusion during natural succession in Macrotermes barneyi. Chin. Bull. Entomol. 1988, 25, 356–359. (In Chinese) [Google Scholar]
- Holdo, R.M.; McDowell, L.R. Termite mounds as nutrient-rich food patches for elephants. Biotropica 2004, 36, 231–239. [Google Scholar] [CrossRef]
- Wood, T.G. Termites and the soil environment. Biol. Fertil. Soils. 1988, 6, 228–236. [Google Scholar] [CrossRef]
- Baker, C.C.; Castillo Vardaro, J.A.; Doak, D.F.; Pansu, J.; Puissant, J.; Pringle, R.M.; Tarnita, C.E. Spatial patterning of soil microbial communities created by fungus-farming termites. Mol. Ecol. 2020, 29, 4487–4501. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Steinegger, M.; Söding, J. MMseqs2 enables sensitive protein sequence searching for the analysis of massive data sets. Nat. Biotechnol. 2017, 35, 1026–1028. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.H.; Lomsadze, A.; Borodovsky, M. Ab initio gene identification in metagenomic sequences. Nucleic Acids Res. 2010, 38, e132. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Kingsford, C. Salmon: Accurate, Versatile and Ultrafast Quantification from RNA-seq Data using Lightweight-Alignment. bioRxiv 2015, 10, 021592. [Google Scholar]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Tilahun, A.; Kebede, F.; Yamoah, C.; Erens, H.; Mujinya, B.B.; Verdoodt, A.; Van Ranst, E. Quantifying the masses of Macrotermes subhyalinus mounds and evaluating their use as a soil amendment. Agric. Ecosyst. Environ. 2012, 157, 54–59. [Google Scholar] [CrossRef]
- Gong, M.F.; Guan, Q.L. Growth conditions of Termitomyces albuminosus under artificial cultivation conditions. Int. J. Agric. Biol. 2020, 23, 135–141. [Google Scholar]
- Chiu, C.I.; Neoh, K.B.; Li, H.F. Colony-founding success of pleometrosis in a fungus-growing termite Odontotermes formosanus. Behav. Ecol. Sociobiol. 2018, 72, 13. [Google Scholar] [CrossRef]
- Kaib, M.; Hacker, M.; Brandl, R. Egg-laying in monogynous and polygynous colonies of the termite Macrotermes michaelseni (Isoptera, Macrotermitidae). Insect. Soc. 2001, 48, 231–237. [Google Scholar] [CrossRef]
- Chiu, C.I.; Ou, J.H.; Chen, C.Y.; Li, H.F. Fungal nutrition allocation enhances mutualism with fungus-growing termite. Fungal Ecol. 2019, 41, 92–100. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, L.; Xue, J.; Cai, Q.; Xu, Y.; Guo, J.; Wei, H.; Huo, Z.; Xu, K.; Zhang, H. Reduced nitrogen application with dense planting achieves high eating quality and stable yield of rice. Foods 2024, 13, 3017. [Google Scholar] [CrossRef]
- Mohamed, I.; El-habbak, A.K.; Abbas, M.H.; Scopa, A.; Drosos, M.; AbdelRahman, M.A.; Bassouny, M.A. Rice straw biochar and NPK minerals for sustainable crop production in arid soils: A case study on maize-wheat cropping system. CABI Agr. Biosci. 2024, 5, 91. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Fischer, M.; Ammer, C.; Boch, S.; Boeddinghaus, R.S.; Bonkowski, M.; Buscot, F.; Fiore-Donno, A.M.; et al. Land-use intensity alters networks between biodiversity, ecosystem functions, and services. Proc. Natl. Acad. Sci. USA 2020, 117, 28140–28149. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Z.; Ma, L.; Zhang, L.; Zhou, X.; Li, H.; Wei, J.; Zheng, F.; Tan, D. Long-term application of pig manure fertiliser affects wheat yield and soil microorganism composition. Plant Soil Environ. 2023, 69, 374–386. [Google Scholar] [CrossRef]
- Shi, Y.; Ziadi, N.; Hamel, C.; Bélanger, G.; Abdi, D.; Lajeunesse, J.; Lafond, J.; Lalande, R.; Shang, J. Soil microbial biomass, activity and community structure as affected by mineral phosphorus fertilization in grasslands. Appl. Soil Ecol. 2020, 146, 103391. [Google Scholar] [CrossRef]
- Chouvenc, T.; Sobotník, J.; Engel, M.S.; Bourguignon, T. Termite evolution: Mutualistic associations, key innovations, and the rise of Termitidae. Cell. Mol. Life Sci. 2021, 78, 2749–2769. [Google Scholar] [CrossRef]
- Tharavecharak, S.; D’Alessandro-Gabazza, C.N.; Toda, M.; Yasuma, T.; Tsuyama, T.; Kamei, I.; Gabazza, E.C. Culture conditions for mycelial growth and anti-cancer properties of Termitomyces. Mycobiology 2023, 51, 94–108. [Google Scholar] [CrossRef]
- Yaguchi, K.; Saito, D.; Menon, T.; Matsura, A.; Hosono, M.; Mizutani, T.; Kotani, T.; Nair, S.; Uehara, R. Haploidy-linked cell proliferation defects limit larval growth in zebrafish. Open Biol. 2024, 14, 240126. [Google Scholar] [CrossRef]
- Martin, F.; Aerts, A.; Ahrén, D.; Brun, A.; Danchin, E.G.J.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef]
- Martinez, D.; Challacombe, J.; Morgenstern, I.; Hibbett, D.; Schmoll, M.; Kubicek, C.P.; Ferreira, P.; Ruiz-Duenas, F.J.; Martinez, A.T.; Kersten, P.; et al. Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proc. Natl. Acad. Sci. USA 2009, 106, 1954–1959. [Google Scholar] [CrossRef]
- Yang, J.J.; Li, W.J.; Teng, D.X.; Yang, X.D.; Zhang, Y.J.; Li, Y. Metagenomic insights into microbial community structure, function, and salt adaptation in saline soils of arid land, China. Microorganisms 2022, 10, 2183. [Google Scholar] [CrossRef]
- Yazhini, G.; Thiyageshwari, S.; Manikandan, A.; Saravanan, V.S.; Selvi, D.; Chithra, L.; Anandham, R. Low and moderate rice-sodic soils affect bacterial diversity and their functions. J. Soil Sci. Plant Nutr. 2024, 24, 855–869. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Qin, W.; Zeng, Y.; Huang, X.; Yuan, J.; Yin, Y.; Xu, P.; Fan, X.; Zhang, R.; Li, G.; et al. Effects of Colony Breeding System and Nest Architecture on Soil Microbiome and Fertility in the Fungus-Growing Termite Macrotermes barneyi Light. Insects 2025, 16, 470. https://doi.org/10.3390/insects16050470
Zhou J, Qin W, Zeng Y, Huang X, Yuan J, Yin Y, Xu P, Fan X, Zhang R, Li G, et al. Effects of Colony Breeding System and Nest Architecture on Soil Microbiome and Fertility in the Fungus-Growing Termite Macrotermes barneyi Light. Insects. 2025; 16(5):470. https://doi.org/10.3390/insects16050470
Chicago/Turabian StyleZhou, Jiachang, Wenquan Qin, Yang Zeng, Xin Huang, Jing Yuan, Yuting Yin, Paike Xu, Xiaohong Fan, Runfeng Zhang, Ganghua Li, and et al. 2025. "Effects of Colony Breeding System and Nest Architecture on Soil Microbiome and Fertility in the Fungus-Growing Termite Macrotermes barneyi Light" Insects 16, no. 5: 470. https://doi.org/10.3390/insects16050470
APA StyleZhou, J., Qin, W., Zeng, Y., Huang, X., Yuan, J., Yin, Y., Xu, P., Fan, X., Zhang, R., Li, G., & Zhang, Y. (2025). Effects of Colony Breeding System and Nest Architecture on Soil Microbiome and Fertility in the Fungus-Growing Termite Macrotermes barneyi Light. Insects, 16(5), 470. https://doi.org/10.3390/insects16050470