1. Introduction
Ceracris kiangsu Tsai 1929, belonging to Orthoptera Acrididae, is an omnivorous migratory pest [
1], which mainly harms bamboos, including
Thysanolaena maxima,
Trachycarpus fortunei, and
Musa basjoo Siebold, as well as crops such as maize, rice, millet, and sorghum [
2,
3].
C. kiangsu is distributed across India, Thailand, Laos, and southern China with an annual single generation [
3]. It overwinters as eggs deposited in egg pods 1–2 cm beneath the soil surface. The hatching time of overwintering eggs varies geographically, occurring from mid-April in northern regions to early June in southern latitudes. The nymphal stage comprises five instars over approximately two months [
4,
5,
6]. Adults attain mating capability 10 days after emergence, which is followed by a requisite 10-day feeding period prior to oviposition. Gravid females preferentially oviposit in loose, sparsely vegetated soils adjacent to bamboo forests, producing one to six egg pods throughout their lifespan, each containing 20–35 eggs. Females die within 5–7 days after oviposition [
7]. Due to
C. kiangsu being a large feeder, long-lasting pest, it is difficult to control [
8]. Nearly 6000 hectares of farmland in Hunan, China, were infested in 1946, resulting in crop losses valued at approximately CNY 3 million. Since 2015, recurrent outbreaks in Laos have severely impacted bamboo forests and gramineous crops (e.g., rice and maize) [
9]. In 2020, large-scale migrations caused extensive agricultural damage in Laos and Vietnam, followed by incursions into Yunnan, China, where over 10,000 hectares of agroforestry land were infested. Localized population densities reached epidemic levels of 200–800 individuals/m
2 [
10], leading to severe ecological and economic consequences.
Early studies concluded that
C. kiangsu could not migrate over long distances [
11], and its destructive power was limited, so there were few studies on the migration of
C. kiangsu. However, the migratory events in recent years have shown that
C. kiangsu can migrate tens of kilometers or nearly 100 km at a time [
12], and the species can migrate many times. The monitoring data show that
C. kiangsu along the border between China and Laos has a straight-line distance of 235 km from the border line to the northernmost landing point, which is much further than the traditional belief that
C. kiangsu can only migrate 100 km in its lifetime [
13]. Therefore, although the flight distance of
C. kiangsu is not far compared with the flying locust, it also has the ability to migrate a medium—and short—distance.
The long-distance migratory process of insects is the result of the interplay between atmospheric background and motion, and most research on migratory pests draws on atmospheric trajectory analysis methods [
14,
15,
16,
17,
18]. China, situated within the East Asian Monsoon Zone, serves as a recognized critical migration corridor for aerial insect biomass flux in East Asia [
19,
20]. Yunnan is influenced by both the Indian monsoon and the East Asian monsoon, and it is also under the control of the atmospheric circulation system of the Tibetan Plateau [
18]. Yunnan serves as the southern gateway of China. Its unique climate and geographical position render it one of the principal entry points for insect migration into China.
Nilaparvata lugens originating from outside China migrates into the southwestern rice-growing regions of Yunnan via the southwest monsoon from northern Thailand and Myanmar [
21];
Spodoptera frugiperda, a “super pest” native to the Americas, enters China through the Indochina Peninsula and Yunnan, and it can migrate northward as far as the northeast of China [
22]; In addition to these, other migratory pests that enter the Chinese mainland through Yunnan include noctuid moths such as
Spodoptera exigua,
Agrotis ipsilon, and
Mythimna separata. Therefore, the border area of Yunnan plays an important role in the management and control of migratory pests.
So, when did C. kiangsu arrived in Yunnan, where did they come from, and where did they go? These questions are crucial for the prevention and control of C. kiangsu from the source. “Where did they come from” requires clarifying the source region of C. kiangsu, and “where did they go” requires tracking their migratory routes. In order to solve these problems, this study, based on the analysis of the migratory dynamics of C. kiangsu, employs trajectory analysis methods to trace the source regions of cross-border migratory C. kiangsu and explore their migratory corridors so as to provide scientific and accurate prevention and control strategies for the prevention and control of C. kiangsu from the source.
3. Results
3.1. Hotspots Distribution of Migratory C. kiangsu Populations
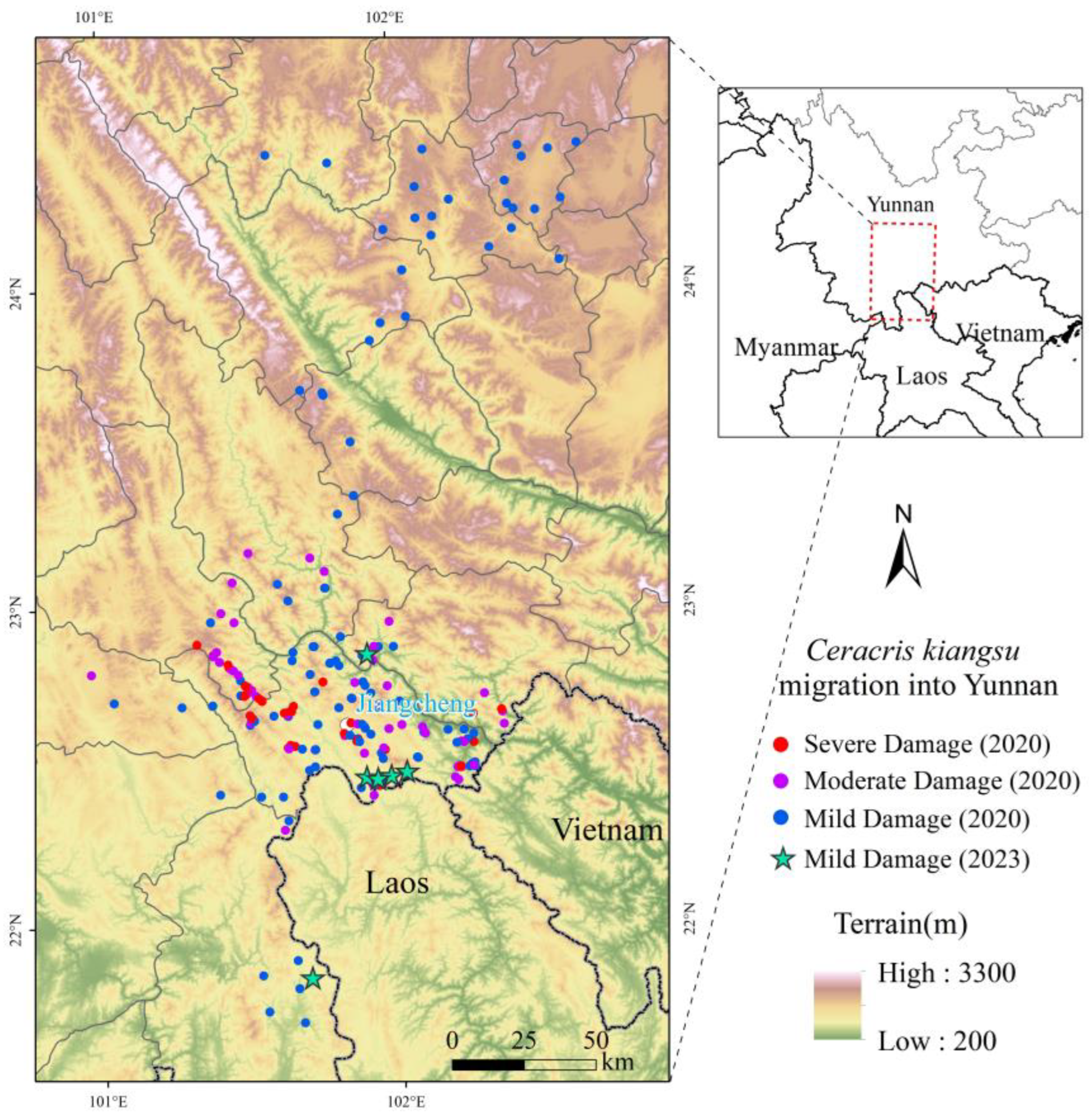
The year 2020 marked the peak of
C. kiangsu migration events and populations abundance since its initial cross-border incursion into China in 2018, which landed in five prefectures and cities in Yunnan Province in batches. A minor migratory population was observed to settle in the China–Laos border area of Jiangcheng County in 2023. Spatially, the migratory populations exhibited distinct distribution patterns (
Figure 2): hotspots were concentrated in the township-level jurisdictions of Jiangcheng County and the adjacent southeastern areas of Lüchun County. Coldspots dominated most of the invaded regions of Yuxi City. Other areas showed no significant clustering or dispersion trends based on the Getis-Ord Gi* spatial autocorrelation analysis.
3.2. Trajectory and Source Regions in Early Migratory Stage
Based on the collated insect data, the initial observed locations and time of the migration of
C. kiangsu across various regions in Yunnan Province were determined (
Table 2). Our analysis revealed that
C. kiangsu typically crosses the border into Yunnan in late June with large-scale populations being observed in the counties of Pu’er City during early July. Concurrently, infestations were detected in Mengla County, Xishuangbanna Prefecture in early July. By mid-July, the locusts had migrated to Lüchun County, Honghe Prefecture and multiple counties in Yuxi City. The pests then reached Shuangbai County, Chuxiong Prefecture in early August, exhibiting an overall expansion pattern progressing from south to north movement followed by east-to-west movement. Representative locations from
Table 2 were selected as simulation starting points for the backward trajectory analysis to reconstruct the early migration pathways of
C. kiangsu.
The earliest cross-border populations of
C. kiangsu in the China–Laos border area of Pu’er City were identified through backward trajectory simulation analysis. This analysis revealed that locusts invading in late June primarily originated from Phongsaly Province, Laos. In early July,
C. kiangsu continued migrating northward. The backward trajectories of initial sightings in Ning’er and Mojiang Counties both traced back to Jiangcheng County, indicating that
C. kiangsu in these two regions migrated from the China–Laos border area. Meanwhile,
C. kiangsu in Simao likely originated from Xishuangbanna (
Figure 3A). The trajectory simulations showed that most locusts in Mengla County, Xishuangbanna, were locally sourced with a minority coming from Laos (
Figure 3B).
In mid-July, locusts were detected in Lüchun County with backward trajectories tracing their origin to Jiangcheng County (
Figure 3C). Initial sightings in Yuxi persisted from mid-July to early August. Locusts found in Yuanjiang during mid-July were traced back to Mojiang and Jiangcheng counties, while those in Xinping County in late July primarily originated from Yuanjiang (
Figure 3D). In Shuangbai County, the backward trajectories (
Figure 3E) indicated that the locusts mainly came from Eshan with a small portion from Hongta. The trajectory simulation results exhibit strong concordance with empirical field observations. Specifically, the backward-traced endpoints for locust swarms documented in Ning’er and Mojiang County during early July were geolocated in Jiangcheng County, which was temporally consistent with the confirmed large-scale influx of
C. kiangsu into this area in late June. Furthermore, the backward trajectory analyses for infestations recorded in Yuanjiang and Xinping County during mid-to-late July revealed Mojiang and Jiangcheng County as probable origins, which was preceded by a distinct migratory peak that had infiltrated multiple districts of Pu’er City. The spatiotemporal distribution of the trajectory endpoints aligns with the peak migration period of
C. kiangsu. This analysis demonstrates that the early-stage locust populations in Yunnan Province primarily originated from Phongsaly, Laos. After entering Pu’er, some locusts migrated further north and west, reaching areas such as Yuxi and Chuxiong.
3.3. Trajectory and Source Regions in Peak Migratory Period
According to the results of the immigration hotspot analysis of
C. kiangsu populations and the migration scale monitored by insect radar, a small number of
C. kiangsu crossed the border from Laos and landed near the border in 2023. Therefore, the study of migration trajectories focused on 2020. During 2020, the peak immigration period of
C. kiangsu from overseas was divided into four intervals, corresponding to four migration peaks: late June (28–30 June), early July (6–8 July), mid-July (14–17 July), and late July (22–25 July). Backward trajectory simulations of the
C. kiangsu were conducted. By integrating the Chinese vegetation dataset with Google Earth satellite imagery, we identified valid trajectory landing points during the migration peak period, which correspond to the species’ source population locations. The simulation results and source population distribution are shown in
Figure 4.
In late June, the backward trajectories of
C. kiangsu migrating to Menglie Township, Jiangcheng County, at different altitudes consistently converged in Phongsaly, Laos (
Figure 4A). In early July, the hotspot distribution areas of
C. kiangsu covered most of Jiangcheng County, southern Ning’er County, and parts of Mengla County. Representative sites were selected for backward trajectory analysis. The trajectories from southern Ning’er predominantly terminated in Zhengdong Township, Jiangcheng County with a minority extending to Xishuangbanna. However, no
C. kiangsu were detected in Xishuangbanna during early July, suggesting that the local populations likely originated from dispersal near Jiangcheng County. The backward trajectories in Mengla County remained within its boundaries. Given the established populations of
C. kiangsu in Mengla County [
12], it is hypothesized that the populations there derived partially from Phongsaly, Laos, and partially from local sources. In Jiangcheng County, most trajectories terminated in Phongsaly, Laos, while the western Zhengdong Township received partial sources from Mengla County. In the eastern Qushui Township, the trajectories at 200 m altitude partially terminated in Dien Bien Province, Vietnam, whereas trajectories at 400 m and 600 m altitudes crossed the Vietnam–Laos border into Phongsaly, Laos (
Figure 4B).
By mid-July, the hotspot areas of migrating
C. kiangsu expanded northward, covering all of Jiangcheng and Ning’er, southern Mojiang, and southwestern Lüchun counties. The backward trajectories from southern Ning’er County indicated origins in Kangping and Zhengdong Township, Jiangcheng. The trajectories from southern Mojiang County clustered in Jiangcheng County and the China–Laos border area. Trajectories from western Lüchun County terminated in Jiahe Township, Jiangcheng, while southern trajectories originated from Qushui Township, Jiangcheng. The trajectories in Jiangcheng County revealed that most sources originated from Phongsaly, Laos with partial sources in eastern Qushui Township from Dien Bien, Vietnam. At 400 m and 600 m altitudes in western Kangping and Zhengdong Township, the trajectories suggested potential origins in Xishuangbanna. Thus, the mid-July migration peak predominantly originated from Phongsaly, Laos, with minimal contributions from Xishuangbanna and Vietnam (
Figure 4C).
In late July, the final migration peak of
C. kiangsu occurred with hotspots concentrated in Jiangcheng County and sporadic influxes into Mojiang and Yuanjiang counties. The backward trajectories for sporadic influxes in these two areas primarily terminated in Jiangcheng County. In Jiangcheng, most trajectories ended in Phongsaly, Laos, except for isolated western trajectories terminating in Xishuangbanna and eastern Qushui Township trajectories in Dien Bien, Vietnam (
Figure 4D).
In summary, the source distribution of C. kiangsu invading Yunnan exhibited pronounced spatial aggregation. During late June and early July, the primary sources originated from Laos with minor contributions from Xishuangbanna. From mid- to late July, populations in areas north of Jiangcheng County derived from the dispersal of previously migrated locusts, while the majority of Jiangcheng County’s sources remained concentrated in Laos, which was supplemented by limited inputs from Vietnam.
3.4. Forward Trajectory of Remigration C. kiangsu in Yunnan
A spatiotemporal analysis of C. kiangsu migration peaks and source region clustering identified mid-to-late July as the mass migration phase with primary landing clusters concentrated in Jiangcheng and Mojiang counties. Since Mojiang County populations derived from either Jiangcheng County or transboundary zones along the China–Laos border, the representative infestation points in Mojiang County were selected as simulation origins for remigration trajectory modeling. Simulations were initiated two days following the termination of the third migratory surge.
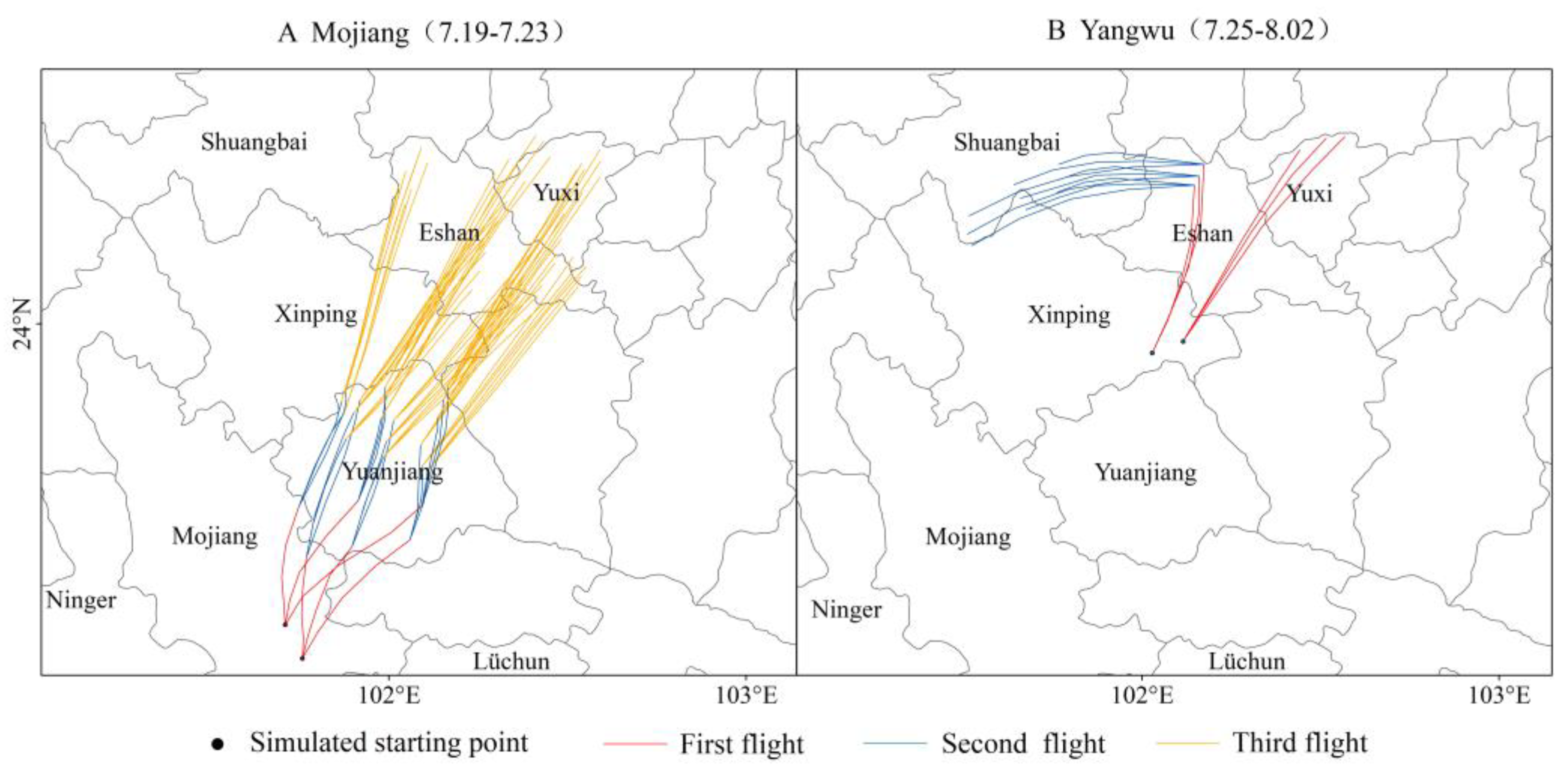
The forward trajectory modeling predicted that
C. kiangsu would reach the northernmost regions of Eshan County and Hongta District after three successive migration events (
Figure 5A). Notably, isolated discrepancies between the simulation outcomes and field observations were identified. Specifically, among the 235 monitoring points collected in 2020, we filtered the migration points for the relevant time periods and found that more points were located in Yangwu Town, Xinping County, rather than in the simulated trajectory endpoint areas. This divergence was driven by precipitation-induced aggregation and partial settlement in Yangwu with remaining populations dispersing northward to Eshan County and Hongta District. Consequently, Yangwu was designated as new starting points for remigration trajectory modeling. The results demonstrated that under conducive meteorological conditions,
C. kiangsu cohorts originating from Yangwu could migrate westward in early August, traversing Eshan County to invade Shuangbai County (
Figure 5B). Subsequent meteorological shifts toward persistent northeasterly airflow patterns suppressed any further northward migration of
C. kiangsu. This finding corresponds with the monitoring data for
C. kiangsu in Shuangbai County during early August, as presented in
Table 2.
The simulation results for Eshan County and Hongta District in
Figure 5B indicated the presence of
C. kiangsu by late July, whereas the field surveys documented in
Table 2 first detected the species in early August. We hypothesized that this temporal mismatch likely arose from constraints (e.g., sampling intervals or spatial coverage limitations) during the field investigations, thereby delaying the detection of the initial migration events.
3.5. Wind Field Analysis During C. kiangsu Migration
The airflow analysis demonstrates that from June to July, a sustained southwesterly airflow has penetrated the 850 hPa level across southwestern China and adjacent regions, persisting with a northward progression trajectory. Under this synoptic configuration, 850 hPa winds over Laos and Yunnan Province exhibited consistent southwesterly orientations in late June. The winds progressively shifted to south–southwesterly directions throughout July. This evolving atmospheric regime established persistent wind corridors, thus facilitating the migration of C. kiangsu populations during their northward migration phase.
Beginning in August, southeasterly–southerly winds prevailed over Laos and Yunnan. This mesoscale circulation pattern redirected the remigration of
C. kiangsu populations northwestward, thereby providing infestation sources to northern Hongta and Chuxiong (
Figure 6E). Notably in the Sino-Vietnamese border area, the air currents entering Yunnan Province from the Vietnamese sector are fundamentally sourced from Laos. This meteorological evidence suggests that
C. kiangsu populations invading Qushui Township, Jiangcheng County via Vietnam likely originated from Laos.
3.6. Migration Corridors of C. kiangsu
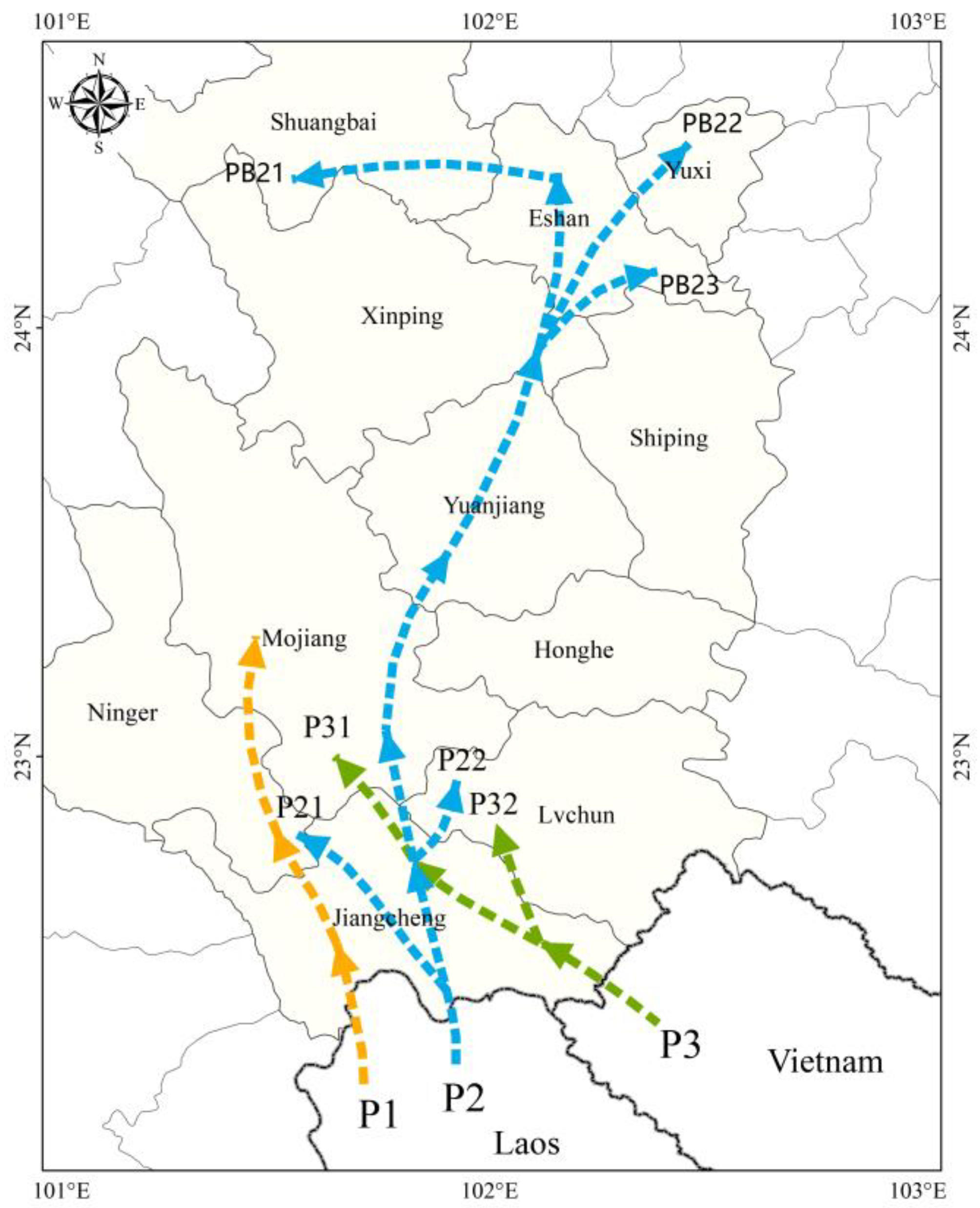
Based on an integrated analysis of
C. kiangsu’s migration trajectories, the meteorological patterns during dispersal periods, and source tracing studies (including the initial migration time and peak migration intensity), three transboundary migration corridors into Yunnan Province were identified (
Figure 7).
Corridor 1 (Laos–Mengkang Port): During early to mid-July, southwest airflow-mediated transport facilitated locust migration from Laos through Mengkang Port → Jiangcheng → Ning’er → the convergence zone southeast of the Ailao Mountains.
Corridor 2 (Laos–Niuluohe): From late June to early August, the main migration axis followed southerly airflow trajectories: Niuluohe Port → Jiangcheng → Mojiang → Yuanjiang → Xinping → Eshan/Hongta → Shuangbai. Branch 1: Northward dispersal from Jiangcheng to Ning’er County; Branch 2: Mid-July southwest airflow-mediated transport to Lüchun.
Corridor 3 (Vietnam–Qushui): From early to late July, southwest–southerly airflow transport enabled migratory populations from Vietnam → southeastern Jiangcheng → partial dispersal to Lüchun County. The Laos–Niuluohe border corridor, exhibiting prolonged migration duration and peak flux density, constituted the principal migration route.
4. Discussion
Since 2018,
C. kiangsu has successively migrated across the China–Laos border into Yunnan, primarily concentrating in Jiangcheng County with smaller distributions being observed in Mengla and Mojiang counties. Bordering the Indochinese Peninsula, Jiangcheng County serves as the first stopover for the northward monsoon-driven migration of these locusts. This pattern highlights exceptionally close insect source exchanges between southern Yunnan and northern Laos [
32], as the Indochinese Peninsula—characterized by a tropical climate, diverse plant species, and complex agricultural systems—functions as a year-round breeding ground for many critical agricultural and forestry pests, earning it the designation of a “perennial insect reservoir”. Furthermore, these low-elevation regions with higher temperatures support extensive growth of bamboo, corn, and rice—key host plants for
C. kiangsu. The favorable habitat provides ample food resources and oviposition sites, meaning that large populations of cross-border migratory locusts typically undertake secondary migrations only when optimal weather conditions coincide.
Research has identified the Niuluohe corridor as the primary migratory route for
C. kiangsu. Monitoring data from consecutive years reveal that both the early and peak migration phases of
C. kiangsu involve cross-border entry into China through this corridor. A critical question arises: Why does the Niuluohe corridor dominate the entire migration cycle of
C. kiangsu? Is this due to its valley terrain facilitating locust movement? Studies on
S. frugiperda indicate that river valleys often act as migration corridors for its populations while also exhibiting barrier and convergence effects on their dispersal [
33]. This raises further questions: Does valley terrain exert similar or analogous influences on
C. kiangsu? If so, how do such topographic features shape insects’ decision-making during migration? These mechanisms represent key priorities for future research.
We assumed that
C. kiangsu migrates passively with wind currents without accounting for its intrinsic flight speed or responses to varying atmospheric conditions. However, insects with autonomous flight capabilities are not entirely passive in aerial displacement; they exhibit active behavioral modulation [
18]. Medium-to-large insects, such as locusts, dragonflies, and noctuid moths, demonstrate collective orientation and stratified aggregation behaviors. These strategies enable rapid and efficient migration, enhance migration success rates, and represent adaptive ecological strategies [
34,
35,
36]. Notably, few studies have scientifically measured the flight speed of
C. kiangsu or clarified its in-flight behavioral strategies. Furthermore, the HYSPLIT model does not incorporate biological flight behaviors (e.g., orientation and airspeed) when simulating insect migration trajectories [
37]. This limitation may compromise the accuracy of the simulated migratory pathways. In the present study, the synergistic effects of wind direction, wind speed, and the self-powered flight speed of
C. kiangsu could not be fully characterized, owing to both the absence of empirical flight speed data and inherent model constraints. Consequently, actual migration distances are likely to exceed simulated estimates. Future research will prioritize the quantification of the species’ flight kinematics to refine trajectory simulation precision.
Insect flight initiation is influenced by multiple factors, particularly the temperature [
38]. Migratory insects typically initiate mass takeoffs under thermally suitable conditions, and airborne swarms actively select altitudes with temperatures meeting their flight threshold [
39,
40]. When ambient temperatures fall below this threshold, swarms either adjust their flight altitude or descend [
41]. For example,
Locusta migratoria are forced to land at temperatures below 19 °C [
42], while
Oedaleus asiaticus exhibits optimal survival at 20–25 °C and demonstrates enhanced flight capacity between 16 and 28 °C [
43]. Although
C. kiangsu belongs to the same taxonomic group, its flight threshold temperature remains uncharacterized. Laboratory experiments indicate that
C. kiangsu achieves peak survival rates at 30 °C, suggesting that its thermal adaptation exceeds that of
O. asiaticus [
44]. Do these findings imply that
C. kiangsu possesses a higher flight threshold temperature compared with other locust species? To address these questions, we will conduct systematic experiments to quantify the intrinsic flight speed, thermal flight range, and abiotic drivers of migration in
C. kiangsu. The further research aims to elucidate the behavioral patterns and environmental triggers underlying its migratory behavior, thereby advancing a precise mechanistic understanding of its flight ecology.
As a natural disaster, locust plagues exhibit certain cyclical patterns in their formation and occurrence. For instance,
Locusta migratoria manilensis shows outbreak intervals ranging from 9–11 years at the longest to 4–5 years at the shortest [
45]. Since the 1990s, four major outbreaks of this subspecies have been recorded in Hebei Province, China [
46]. However, the periodicity of locust plagues is not isolated; it is closely linked to factors such as drought and the overexploitation of land, which create intrinsic causal relationships [
11]. These factors also contribute to the synchronized, large-scale emergence of locust swarms. Yunnan experienced its first migration of
C. kiangsu from neighboring countries in 2018. Subsequent sporadic border crossings occurred in 2019, 2021, and 2022 with a massive migratory influx in 2020 causing significant agricultural damage. Small-scale populations were detected in 2023, but no incursions have been observed as of 2022. Meteorological analysis suggests that the early spring drought and high temperatures in Laos during 2020 accelerated egg hatching, improved survival rates, and advanced migration timelines, creating optimal conditions for the early and large-scale migration. Beyond climatic drought, habitat alterations due to land-use changes also contributed to these outbreaks.
Global climate change is becoming increasingly frequent with a rise in extreme weather events. Studies indicate that under the context of global warming, particularly the rising winter temperatures, the overwintering eggs of locusts are more likely to survive, providing a “seed bank” for outbreaks of locust infestations in the following year. Additionally, the compounding effects of intensified droughts, pasture degradation, and other factors create suitable breeding grounds for locusts to lay eggs, coupled with their strong adaptability to arid conditions. Consequently, due to expanding habitats, phenological changes, and migratory behaviors, the frequency and scale of
C. kiangsu outbreaks may escalate significantly in the future [
47]. Therefore, clarifying the migration timing and pathways of
C. kiangsu holds critical importance for implementing targeted control measures across regions and timeframes, thereby enhancing pest management efficiency.
5. Conclusions
Our findings demonstrate that Laos constitutes the principal source region for the transboundary migrations of
C. kiangsu. Populations initially dispersed from Laos are hypothesized to traverse Vietnamese airspace via atmospheric currents before infiltrating Yunnan Province, underscoring the China–Laos border as a critical focal area for
C. kiangsu control. Following the 2018 migration, small established populations were observed in Mengla County [
12], providing localized sources for subsequent domestic migration. In addition to the three transboundary migration corridors, a fourth domestic route was identified. Furthermore, airflow analysis suggests that Mengla County may become a potential high risk deposition zone for cross-border
C. kiangsu migrations in August.
Meteorological forcing emerges as the principal extrinsic factor governing C. kiangsu dispersal dynamics. In July, prevailing southwestern winds over southern Yunnan facilitate the sustained northward migration of populations from the Indochina Peninsula and southern Yunnan, enabling locusts to reach Yuxi City. However, the shift to southeastern winds in early August partially disrupts these northward movements. Precipitation forces C. kiangsu to land, with locusts preferentially selecting habitats dominated by preferred host plants, such as bamboo and maize. Trajectory analysis of C. kiangsu cohorts departing Mojiang County revealed an initial dispersal toward Eshan County and the Hongta District northern periphery. However, mid-flight interception by precipitation systems resulted in a forced deposition at Yangwu Township, Xinping County. The subsequent resumption of westerly migration toward Chuxiong Prefecture under improved meteorological conditions was temporally congruent with the field surveillance data, empirically validating precipitation as a critical modulator of depositional phase transitions in C. kiangsu migratory cycles.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}