
Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies

 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honeybee Colonies and Treatments
2.2. Assessment of Varroa Mite Infestations and Honeybee Colony Strength
2.3. Honeybee Samples
2.4. RNA Extraction and Reverse Transcription
2.5. Primer Design
2.6. Droplet Digital Polymerase Chain Reaction (ddPCR)
2.7. Statistical Analysis
3. Results
3.1. Honeybee Colony Growth and Development
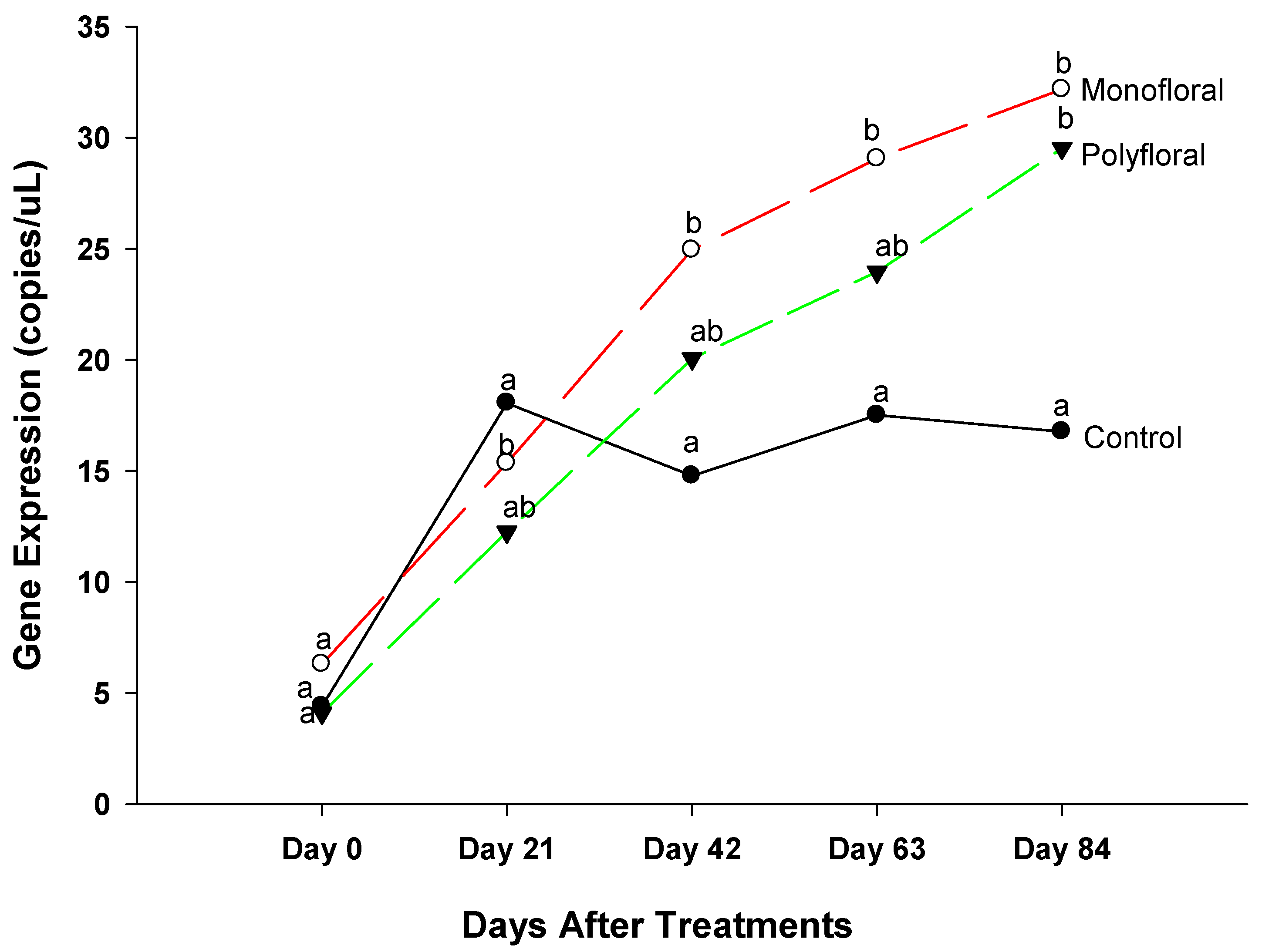
3.2. Expression of Genes Linked to Honeybee Health Fed with Different Diets
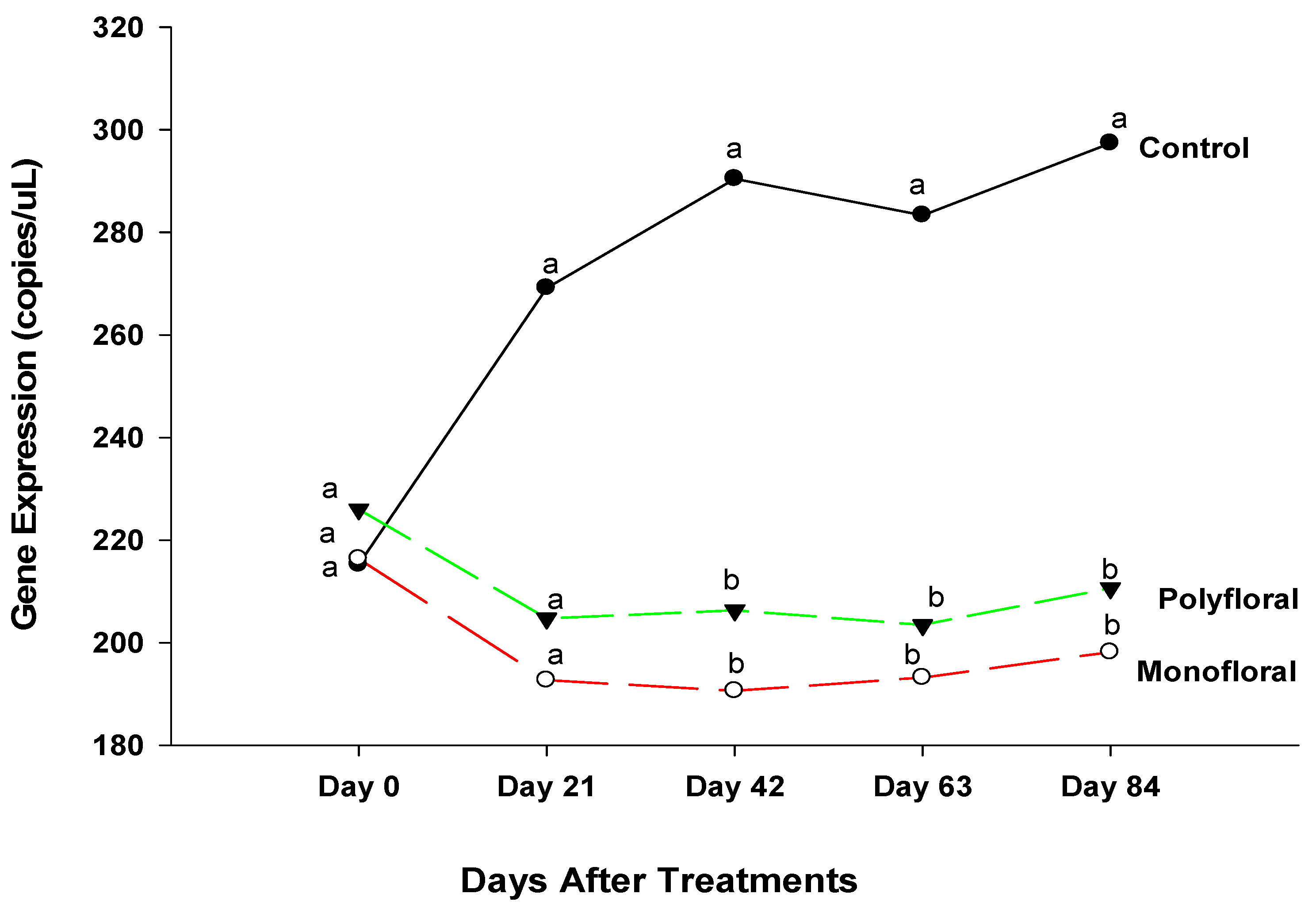
3.2.1. Cactus Gene Expression
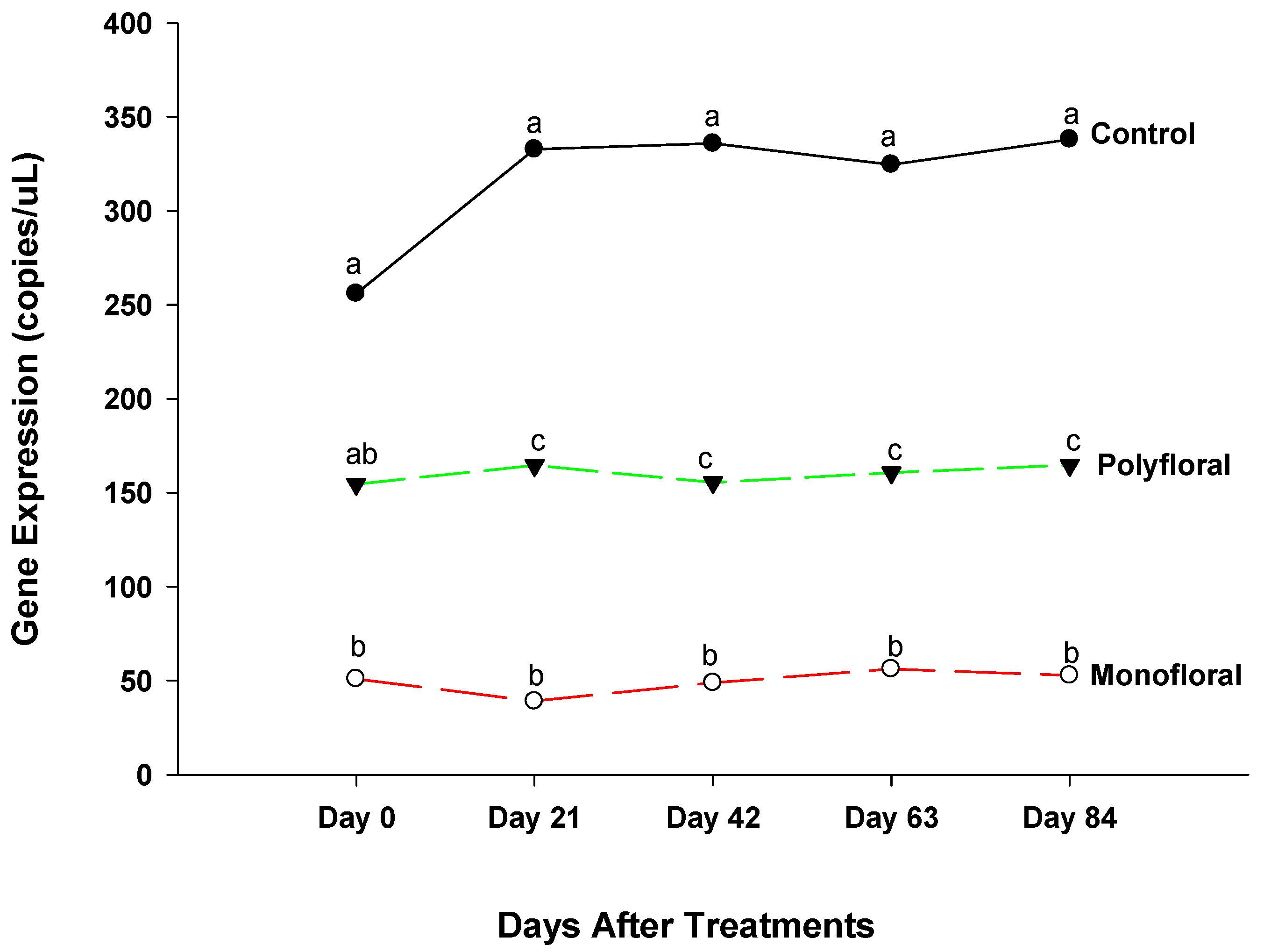
3.2.2. Immune Deficiency Gene Expression
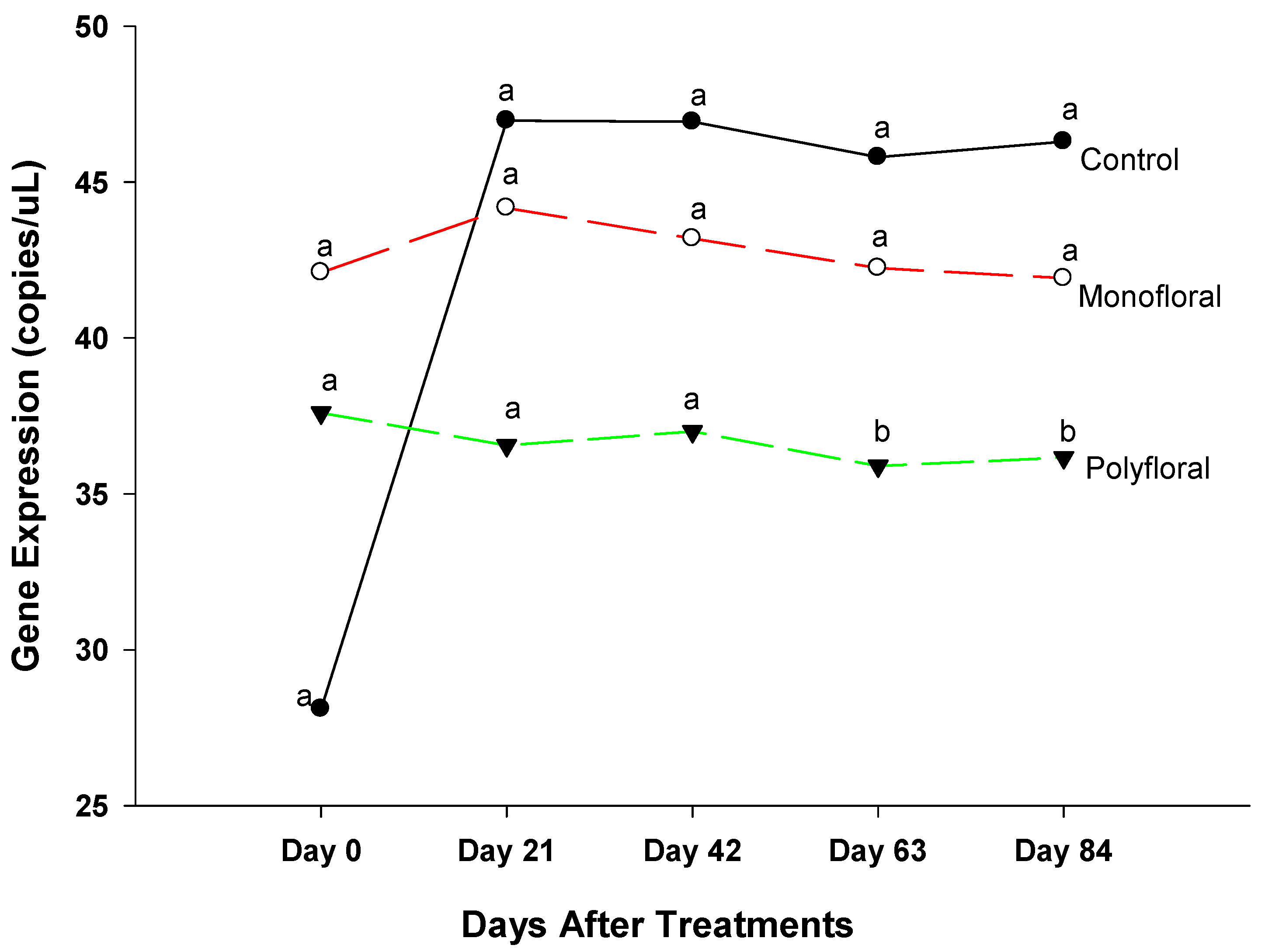
3.2.3. Spaetzle Gene Expression
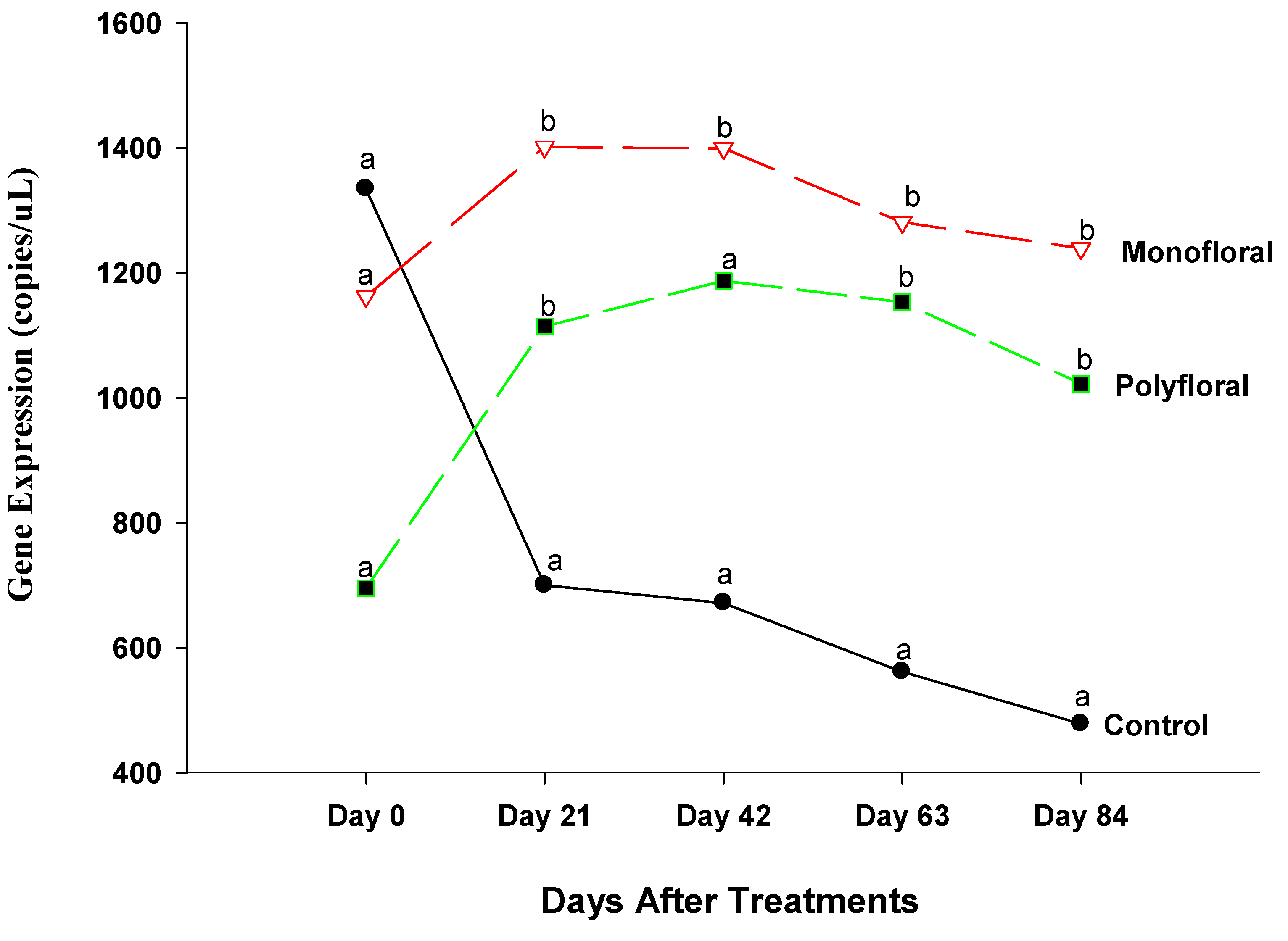
3.2.4. Vitellogenin Gene Expression
3.2.5. Malvolio Gene Expression
3.2.6. Maltase Gene Expression
3.2.7. Single-Minded Homolog 2 Gene
4. Discussion
4.1. Cactus Gene Expression
4.2. Immune Deficiency
4.3. Spaetzle
4.4. Vitellogenin
4.5. Malvolio
4.6. Maltase
4.7. Single-Minded Homologue 2
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, E.B. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native pollinators in anthropogenic habitats. Annu. Rev. Ecol. Syst. 2011, 42, 1–22. [Google Scholar]
- Frankie, G.W.; Thorp, R.W.; Coville, R.E.; Ertter, B. California Bees and Blooms: A Guide for Gardeners and Naturalists; Heyday Books: Berkeley, CA, USA, 2014. [Google Scholar]
- Yang, X.; Cox-Foster, D. Effects of parasitization by Varroa destructor on survivorship and physiological traits of Apis mellifera in correlation with viral incidence and microbial challenge. Parasitology 2007, 134, 405–412. [Google Scholar] [CrossRef]
- Ellis, R.D.; McWhorter, T.J.; Maron, M. Integrating landscape ecology and conservation physiology. Landsc. Ecol. 2012, 27, 1–12. [Google Scholar]
- Johnson, M.R.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Acaricide, fungicide and drug interactions in honeybees (Apis mellifera). PLoS ONE 2013, 8, e54092. [Google Scholar] [CrossRef]
- Alaux, C.; Dantec, C.; Parrinello, H.; Conte, Y.L. Nutrigenomics in honeybees: Digital gene expression analysis of pollen’s nutritive effects on healthy and Varroa-parasitized bees. BMC Genom. 2011, 12, 496. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D. A metagenomic survey of microbes in honeybee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar]
- Gregorc, A.; Evans, J.D.; Scharf, M.; Ellis, J.D. Gene expression in honeybee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). J. Insect Physiol. 2012, 58, 1042–1049. [Google Scholar]
- Kanga, L.H.B.; Siebert, S.C.; Sheikh, M.; Legaspi, J.C. Pesticide residues in conventionally and organically managed apiaries in South and North Florida. Curr. Investig. Agric. Curr. Res. 2019, 7, 937–940. [Google Scholar]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defense mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar]
- Osta, M.A.; Christophides, G.K.; Vlachou, D.; Kafatos, F.C. Innate immunity in the malaria vector Anopheles gambiae: Comparative and functional genomics. J. Exp. Biol. 2004, 207, 2551–2563. [Google Scholar] [CrossRef] [PubMed]
- Boutros, M.; Agaisse, H.; Perrimon, N. Sequential activation of signaling pathways during innate immune responses in Drosophila. Dev. Cell 2002, 3, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Rich, N.; Dres, S.T.; Starks, P.T. The ontogeny of immunity: Development of innate immune strength in the honey bee (Apis mellifera). J. Insect. Physiol. 2008, 54, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Arévalo, S.; Vicente-Rubiano, M.; Puerta, F.; Molero, F.; Sanchez-Vizcaino, J.M. Immune related genes as markers for monitoring health status of honey bee colonies. BMC Vet. Res. 2019, 15, 72. [Google Scholar] [CrossRef]
- Hendriksma, H.P.; Shafir, S. Honey bee foragers balance colony nutritional deficiencies. Behav. Ecol. Sociobiol. 2016, 70, 509–517. [Google Scholar] [CrossRef]
- Zarchin, S.; Dag, A.; Salomon, M.; Hendriksma, H.P.; Shafir, S. Honey bees dance faster for pollen that complements colony essential fatty acid deficiency. Behav. Ecol. Sociobiol. 2017, 71, 172. [Google Scholar] [CrossRef]
- Schmidt, J.O.; Thoenes, S.C.; Levin, M.D. Survival of honey bees, Apis mellifera (Hymenoptera: Apidae), fed various pollen sources. J. Econ. Entomol. 1987, 80, 176–183. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef]
- Gould, J.L.; Gould, C.G. The Honeybee; Scientific American Library: New York, NY, USA, 1988. [Google Scholar]
- USDA Forest Service. Future of America’s Forests and Rangelands: Update to the Forest Service. 2010. Resources Planning Act Assessment. Gen. Tech. Rep. WO-GTR-94. Washington, DC. 2016. Available online: https://www.fs.fed.us/research/publications/gtr/gtr_wo94.pdf (accessed on 12 November 2024).
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M.H. The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honeybees (Apis mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Toth, A.L. Feedbacks between nutrition and disease in honey bee health. Curr. Opin. Insect Sci. 2018, 26, 114–119. [Google Scholar] [CrossRef]
- Williams, N.M. Use of novel pollen species by specialist and generalist solitary bees (Hymenoptera: Megachilidae). Oecologia 2003, 134, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Bryś, M.S.; Skowronek, P.; Strachecka, A. Pollen diet-properties and impact on a bee colony. Insects 2021, 12, 798. [Google Scholar] [CrossRef] [PubMed]
- Landis, A.D.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar]
- Duda, M.M.; Matei, C.F.; Varban, D.I. Agastache foeniculum (Pursh) Kuntze-a new plant in the landscape of our country. Hop Med. Plants 2013, 19, 63–65. [Google Scholar] [CrossRef]
- Saeedfar, S.; Negahban, M.; Soorestani, M.M. The effect of drought stress on the essential oil content and some of the biochemical characteristics of anise hyssop (Agastache foeniculum [Pursh] Kuntze). Eur. J. Mol. Biotechnol. 2015, 8, 103–114. [Google Scholar]
- Abou-Shaara, H. Potential Honey Bee Plants of Egypt. Agron. Res. Mold. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Browning, A.; Smitley, D.; Studyvin, J.; Runkle, E.S.; Huang, Z.Y.; Hotchkiss, E. Variation in pollinator visitation among garden cultivars of marigold, portulaca, and bidens. J. Econ. Entomol. 2023, 116, 872–881. [Google Scholar]
- Leponiemi, M.; Freitak, D.; Moreno-Torres, M.; Eva-Maria, P.W.; Becker-Scarpitta, A.; Tiusanen, M.; Vesterinen, E.J.; Wirta, H. Honeybees’ foraging choices for nectar and pollen revealed by DNA metabarcoding. Sci. Rep. 2023, 13, 14753. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Van Der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar]
- McGregor, S.E.; Rowe, J.B. Honeybee colony quality for alfalfa pollination. Am. Bee J. 1979, 119, 700–703. [Google Scholar]
- Kanga, L.H.B.; Jones, W.A.; James, R.R. Field trials using the fungal pathogen, Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) to control the ectoparasitic mite, Varroa destructor (Acari: Varroidae) in honeybee, Apis mellifera (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2003, 96, 1091–1099. [Google Scholar] [PubMed]
- Viljakainen, L.; Evans, D.J.; Hasselmann, M.; Rueppell, O.; Tingek, S.; Pamilo, P. Rapid evolution of immune proteins in social insects. J. Mol. Biol. Evol. 2009, 26, 1791–1801. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, W.M.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar]
- Lourenço, A.P.; Florecki, M.M.; Simões, Z.L.P.; Evans, J.D. Silencing of Apis mellifera dorsal genes reveals their role in expression of the antimicrobial peptide defensin-1. Insect Mol. Biol. 2018, 27, 577–589. [Google Scholar] [PubMed]
- Scharlaken, B.; de Graaf, D.C.; Goossens, K.; Brunain, M.; Peelman, L.J.; Jacobs, F.J. Reference gene selection for insect expression studies using quantitative real-time PCR: The head of the honeybee, Apis mellifera, after a bacterial challenge. J. Insect Sci. 2008, 8, 33. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT Software: Factors Analysis and Structural Equation Modeling, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Aronstein, K.; Pankiw, T.; Saldivar, E. SID-I is implicated in systemic gene silencing in the honeybee. J. Apic. Res. 2006, 45, 20–24. [Google Scholar]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E., Jr.; Amdam, G.V. The gene Vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, e62. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.H.; Rämet, M. The Drosophila Toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [PubMed]
- Valanne, S.; Kallio, J.; Kleino, A.; Rämet, M. Large-scale RNAi screens add both clarity and complexity to Drosophila NF-κB signaling. Dev. Comp. Immunol. 2012, 37, 9–18. [Google Scholar]
- Gilmore, T.D.; Wolenski, F.S. NF-κB: Where did it come from and why? Immunol. Rev. 2012, 246, 14–35. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Hood, W.M. Economic threshold for Varroa jacobsoni Oud. in the southeastern USA. Apidologie 1999, 30, 383–395. [Google Scholar]
- Lindsay, S.A.; Wasserman, S.A. Conventional and non- conventional Drosophila Toll signaling. Dev. Comp. Immunol. 2014, 42, 16–24. [Google Scholar] [PubMed]
- Maruščáková, C.I.; Schusterová, P.; Bielik, B.; Toporccak, J. Effect of Application of Probiotic Pollen Suspension on Immune Response and Gut Microbiota of Honey Bees (Apis mellifera). Probiotics Antimicrob. Proteins 2022, 12, 929–936. [Google Scholar] [CrossRef]
- Ferrandon, D.; Imler, J.L.; Hoffmann, J.A. Sensing infection in Drosophila: Toll and beyond. Semin. Immunol. 2004, 16, 43–53. [Google Scholar]
- Sabin, L.R.; Hanna, S.L.; Cherry, S. Innate antiviral immunity in Drosophila. Curr. Opin. Immunol. 2010, 22, 4–9. [Google Scholar] [PubMed]
- Tanji, T.; Ip, Y.T. Regulators of the toll and IMD pathways in the Drosophila innate immune response. Trends Immunol. 2005, 26, 193–198. [Google Scholar]
- Hoffmann, J. Antifungal defense in Drosophila. Nat. Immunol. 2007, 8, 543–545. [Google Scholar] [CrossRef]
- Ferrandon, D.; Imler, J.L.; Hetru, C.; Hoffmann, J.A. The Drosophila systemic immune response: Sensing and signaling during bacterial and fungal infections. Nat. Rev. Immunol. 2007, 7, 862–874. [Google Scholar]
- Chaimanee, V.; Chantawannakul, P.; Chen, Y.; Evans, J.D.; Pettis, J.S. Differential expression of immune genes of adult honeybee (Apis mellifera) after inoculated by Nosema ceranae. J. Insect Physiol. 2012, 58, 1090–1095. [Google Scholar] [CrossRef]
- Seehuus, S.-C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honeybee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honeybee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef]
- Amdam, G.V.; Simões, Z.L.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, O.; Kirkwood, T.B.; Omholt, S.W. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.; Underwood, R.; Schwarz, R.; Evans, J.D.; Pettis, J. Direct effect of acaricides on pathogen loads and gene expression levels in honeybees Apis mellifera. J. Insect Physiol. 2012, 58, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Orgad, S.; Nelson, H.; Segal, D.; Nelson, N. Metal ions suppress the abnormal taste behavior of the Drosophila mutant malvolio. J. Exp. Biol. 1998, 201, 115–120. [Google Scholar]
- Ben-Shahar, Y.; Dudek, N.L.; Robinson, G.E. Phenotypic deconstruction reveals involvement of manganese transporter malvolio in honeybee division of labor. J. Exp. Biol. 2004, 207, 3281–3288. [Google Scholar] [CrossRef]
- Zayed, A.; Robinson, G.E. Understanding the relationship between brain gene expression and social behavior: Lessons from the honeybee. Annu. Rev. Genet. 2012, 46, 591–615. [Google Scholar]
- Seeley, T.D. Adaptive significance of the age polyethism schedule in honeybee colonies. Behav. Ecol. Sociobiol. 1982, 11, 287–293. [Google Scholar] [CrossRef]
- Page, R.E.; Scheiner, R.; Erber, J.; Amdam, G.V. The development and evolution of division of labor and foraging specialization in a social insect (Apis mellifera L.). Curr. Top. Dev. Biol. 2006, 74, 253–286. [Google Scholar]
- Siebert, S.C.; Kanga, L.H.; Basha, S.M.; Legaspi, J.C. Molecular assessment of genes linked to immune response traits of honeybees in conventional and organically managed apiaries. Insects 2020, 11, 637. [Google Scholar] [CrossRef]
- Wheeler, M.; Robinson, G. Diet-dependent gene expression in honeybees: Honey vs. sucrose or high fructose corn syrup. Sci. Rep. 2014, 4, 5726. [Google Scholar] [CrossRef]
- Gmeinbauer, R.; Crailsheim, K. Glucose utilization during flight of honeybee (Apis mellifera) workers, drones and queens. J. Insect Physiol. 1993, 39, 959–967. [Google Scholar]
- Becker, A.; Schlöder, P.; Steele, J.E.; Wegener, G. The regulation of trehalose metabolism in insects. Experientia 1996, 52, 433–439. [Google Scholar] [PubMed]
- Blatt, J.; Roces, F. Haemolymph sugar levels in foraging honeybees (Apis mellifera carnica): Dependence on metabolic rate and in vivo measurement of maximal rates of trehalose synthesis. J. Exp. Biol. 2001, 204, 2709–2716. [Google Scholar]
- Crews, S.T.; Thomas, J.B.; Goodman, C.S. The Drosophila single-minded gene encodes a nuclear protein with sequence similarity to the per gene product. Cell 1988, 52, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.B.; Crews, S.T.; Goodman, C.S. Molecular genetics of the single- minded locus: A gene involved in the development of the Drosophila nervous system. Cell 1988, 52, 133–141. [Google Scholar]
- Pielage, J.; Steffes, G.; Lau, D.C.; Parente, B.A.; Crews, S.T.; Strauss, R.; Klämbt, C. Novel behavioral and developmental defects associated with Drosophila single-minded. Dev. Biol. 2002, 249, 283–299. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | Function |
|---|---|---|---|---|
| Cactus | FJ546095.1 | ACATAGTTCGG GCCACACTG | AGGTGCGGTTGCA GTATTCA | Immune response [35] |
| Immune deficiency (IMD) | NM_001163717.2 | ACCCGCCAAAT GCCAATAGA | AGTCGATGGTGGTA ATGGTACT | Antimicrobial defense [37] |
| Spaetzle | XM_026444398.1 | GTGTCAGTGGC GGTGACTAA | GCACGTGTTGATG TATCCGC | Member of the TOLL-immune signaling pathway against bacteria and fungi [36] |
| Vitellogenin (Vg) | NM_001011578.1 | AACGCTTTTACTG TTCGCGG | TATGCACGTCCGAC AGATCG | Immune function and longevity [36] |
| Maltase | XM_006564751.3 | CGAAAGCAGCAAC GAATGGG | ACAGGTTTATCGC TGTTACCGA | Energy metabolism [36] |
| Malvolio (mvl) | XM_006563052.3 | TCCCCGCCAAGAT CACATTT | ACCACACCAAGTCT TGCACT | Involved in sucrose responsiveness [36] |
| Single-minded homolog 2 | XM_016914389.2 | TGCGATCGGGAGA AAGTGTC | TTTCGCCTCCAACT ACCGAC | Locomotor behavior [36] |
| Treatments | 1 Number of Mites per Hundred Adult Bees (Mean ± SE) | 2 Colony Strength Parameters (Mean ± SE) | ||||
|---|---|---|---|---|---|---|
| Bees | Honey | |||||
| Before | After | Before | After | Before | After | |
| Control | 0.11 ± 0.01 aA | 0.94 ± 0.61 aA | 5.20 ± 0.52 aA | 0.72 ± 0.67 aB | 2.28 ± 0.30 aA | 1.67 ± 1.07 aA |
| Monofloral | 0.35 ± 0.14 aA | 0.41 ± 0.19 aA | 5.04 ± 1.00 aA | 2.92 ± 0.82 aA | 3.08 ± 0.32 aA | 0.32 ± 0.19 aB |
| Polyfloral | 0.01 ± 0.00 aA | 0.20 ± 0.12 aA | 5.54 ± 0.27 aA | 1.12 ± 0.73 aB | 2.14 ± 0.27 aA | 0.01 ± 0.00 aB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diedrick, W.A.; Kanga, L.H.B.; Mallinger, R.; Pescador, M.; Elsharkawy, I.; Zhang, Y. Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies. Insects 2025, 16, 374. https://doi.org/10.3390/insects16040374
Diedrick WA, Kanga LHB, Mallinger R, Pescador M, Elsharkawy I, Zhang Y. Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies. Insects. 2025; 16(4):374. https://doi.org/10.3390/insects16040374
Chicago/Turabian StyleDiedrick, Worrel A., Lambert H. B. Kanga, Rachel Mallinger, Manuel Pescador, Islam Elsharkawy, and Yanping Zhang. 2025. "Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies" Insects 16, no. 4: 374. https://doi.org/10.3390/insects16040374
APA StyleDiedrick, W. A., Kanga, L. H. B., Mallinger, R., Pescador, M., Elsharkawy, I., & Zhang, Y. (2025). Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies. Insects, 16(4), 374. https://doi.org/10.3390/insects16040374