
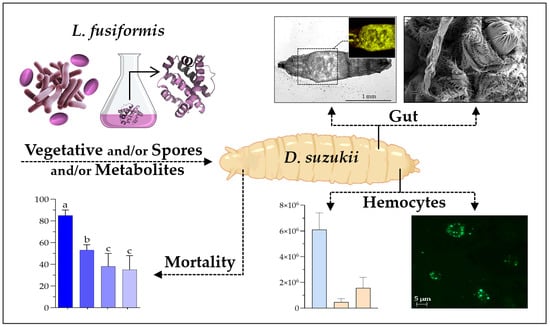
Evaluation of the Insecticidal Potential of Lysinibacillus fusiformis Against Drosophila suzukii Larvae

 ,
,  , , , , ,
, , , , ,  and
and 
Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. Bacteria Isolation
2.3. Target Insect
2.4. L. fusiformis Cultures for Bioassays
2.5. Larvicidal Activity Against D. suzukii
2.6. Electrophoretic Analysis (SDS-PAGE)
2.7. Morphology of Treated D. suzukii Larvae
2.8. L. fusiformis Effect on D. suzukii Hemocytes
2.9. Data Analyses
3. Results
3.1. L. fusiformis Isolation and Culture
3.2. Effects of L. fusiformis Administration to D. suzukii Larvae
3.3. Effects of L. fusiformis on the Gut of D. suzukii Larvae
3.4. Scanning Electron Microscopy of Control and L. fusiformis-Treated D. suzukii Larvae
3.5. Total Hemocytes Count (THC) and Phagocytosis Assay of L. fusiformis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef]
- Kaur, R.; Choudhary, D.; Bali, S.; Bandral, S.S.; Singh, V.; Ahmad, M.A.; Rani, N.; Singh, T.G.; Chandrasekaran, B. Pesticides: An Alarming Detrimental to Health and Environment. Sci. Total Environ. 2024, 915, 170113. [Google Scholar] [CrossRef]
- Thilagam, P.; Sharanappa, C.H.; Roy, S.; Deb, L.; Padhan, S.; Srividhya, S.; Awadhiya, P. A Review on Advances in Biocontrol Techniques for Managing Insect Pests in Sustainable Agriculture. Int. J. Environ. Clim. Change 2023, 13, 2114–2125. [Google Scholar] [CrossRef]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef]
- Tarasco, E.; Fanelli, E.; Salvemini, C.; El-Khoury, Y.; Troccoli, A.; Vovlas, A.; De Luca, F. Entomopathogenic Nematodes and Their Symbiotic Bacteria: From Genes to Field Uses. Front. Insect Sci. 2023, 3, 1195254. [Google Scholar] [CrossRef]
- Sudakin, D.L. Biopesticides. Toxicol. Rev. 2003, 22, 83–90. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H.; et al. Plant Growth-Promoting Microorganisms as Biocontrol Agents of Plant Diseases: Mechanisms, Challenges and Future Perspectives. Front. Plant Sci. 2022, 6, 923880. [Google Scholar] [CrossRef] [PubMed]
- Šunjka, D.; Mechora, Š. An Alternative Source of Biopesticides and Improvement in Their Formulation—Recent Advances. Plants 2022, 20, 3172. [Google Scholar] [CrossRef]
- Sabbahi, R.; Hock, V.; Azzaoui, K.; Saoiabi, S.; Hammouti, B. A Global Perspective of Entomopathogens as Microbial Biocontrol Agents of Insect Pests. J. Agric. Food Res. 2022, 10, 100376. [Google Scholar] [CrossRef]
- Silva-Filha, M.H.N.L.; Romão, T.P.; Rezende, T.M.T.; Carvalho, K.d.S.; de Menezes, H.S.G.; Do Nascimento, N.A.; Soberón, M.; Bravo, A. Bacterial Toxins Active against Mosquitoes: Mode of Action and Resistance. Toxins 2021, 27, 523. [Google Scholar] [CrossRef]
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A Successful Insecticide with New Environmental Features and Tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis Toxins: An Overview of Their Biocidal Activity. Toxins 2014, 11, 3296–3325. [Google Scholar] [CrossRef]
- Heckel, D.G. How Do Toxins from Bacillus thuringiensis Kill Insects? An Evolutionary Perspective. Arch. Insect Biochem. Physiol. 2020, 104, e21673. [Google Scholar] [CrossRef]
- Luis Jurat-Fuentes, J.; Heckel, D.G.; Ferré, J. Mechanisms of Resistance to Insecticidal Proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–161. [Google Scholar] [CrossRef] [PubMed]
- Jamal, Q.M.S.; Ahmad, V. Lysinibacilli: A Biological Factories Intended for Bio-Insecticidal, Bio-Control, and Bioremediation Activities. J. Fungi 2022, 8, 1288. [Google Scholar] [CrossRef] [PubMed]
- Naureen, Z.; Ur Rehman, N.; Hussain, H.; Hussain, J.; Gilani, S.A.; Al Housni, S.K.; Mabood, F.; Khan, A.L.; Farooq, S.; Abbas, G.; et al. Exploring the Potentials of Lysinibacillus sphaericus ZA9 for Plant Growth Promotion and Biocontrol Activities against Phytopathogenic Fungi. Front. Microbiol. 2017, 8, 1477. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Santana, A.; Gómez-Garzón, C.; Dussán, J. Lysinibacillus sphaericus. Trends Microbiol. 2022, 30, 705–706. [Google Scholar] [CrossRef]
- McMillan, J.R.; Olson, M.M.; Petruff, T.; Shepard, J.J.; Armstrong, P.M. Impacts of Lysinibacillus sphaericus on Mosquito Larval Community Composition and Larval Competition between Culex Pipiens and Aedes Albopictus. Sci. Rep. 2022, 12, 18013. [Google Scholar] [CrossRef]
- Yu, S.; Liu, Z.; Wang, J.; Zheng, H.; Han, S.; Zheng, F.; Zheng, D.; Zhao, C.; Li, X.; Liu, T.; et al. Lysinibacillus sphaericus Exposure Impedes Anopheles Dirus’s Oviposition via Downregulating Vitellogenin. Parasites Vectors 2025, 18, 111. [Google Scholar] [CrossRef]
- Silva Filha, M.H.N.L.; Berry, C.; Regis, L. Chapter Three—Lysinibacillus sphaericus: Toxins and Mode of Action, Applications for Mosquito Control and Resistance Management. In Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 47, pp. 89–176. ISBN 0065-2806. [Google Scholar]
- Nascimento, N.A.; Torres-Quintero, M.C.; Molina, S.L.; Pacheco, S.; Romão, T.P.; Pereira-Neves, A.; Soberón, M.; Bravo, A.; Neves, M.H.; Silva-Filha, L.; et al. Functional Bacillus thuringiensis Cyt1Aa Is Necessary To Synergize Lysinibacillus sphaericus Binary Toxin (Bin) against Bin-Resistant and-Refractory Mosquito Species. Appl. Environ. Microbiol. 2020, 86, e02770-19. [Google Scholar] [CrossRef]
- Passera, A.; Rossato, M.; Oliver, J.S.; Battelli, G.; Shahzad, G.I.R.; Cosentino, E.; Sage, J.M.; Toffolatti, S.L.; Lopatriello, G.; Davis, J.R.; et al. Characterization of Lysinibacillus fusiformis Strain S4C11: In Vitro, in Planta, and in Silico Analyses Reveal a Plant-Beneficial Microbe. Microbiol. Res. 2021, 244, 126665. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, V.; Muhammad Zafar Iqbal, A.N.; Haseeb, M.; Khan, M.S. Antimicrobial Potential of Bacteriocin Producing Lysinibacillus Jx416856 against Foodborne Bacterial and Fungal Pathogens, Isolated from Fruits and Vegetable Waste. Anaerobe 2014, 27, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Pudova, D.S.; Lutfullin, M.T.; Shagimardanova, E.I.; Hadieva, G.F.; Shigapova, L.; Toymentseva, A.A.; Kabanov, D.A.; Mardanova, A.M.; Vologin, S.G.; Sharipova, M.R. Draft Genome Sequence Data of Lysinibacillus fusiformis Strain GM, Isolated from Potato Phyllosphere as a Potential Probiotic. Data Brief 2018, 21, 2504–2509. [Google Scholar] [CrossRef]
- Qiu, X.; Qian, Y.; Zhou, R.; Zhou, Y.; Wang, Y.; Tan, Y. Isolation and Algicidal Effect of Algicidal Bacterium Strain N25. J. Shanghai Jiaotong Univ. (Med. Sci.) 2011, 31, 1375. [Google Scholar] [CrossRef]
- Trivedi, P.; Spann, T.; Wang, N. Isolation and Characterization of Beneficial Bacteria Associated with Citrus Roots in Florida. Microb. Ecol. 2011, 62, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Rahmoune, B.; Morsli, A.; Khelifi-Slaoui, M.; Khelifi, L.; Strueh, A.; Erban, A.; Kopka, J.; Prell, J.; Van Dongen, J.T. Isolation and Characterization of Three New PGPR and Their Effects on the Growth of Arabidopsis and Datura Plants. J. Plant Interact. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- De Mandal, S.; Singh, S.S.; Kumar, N.S. Analyzing Plant Growth Promoting Bacillus Sp. and Related Genera in Mizoram, Indo-Burma Biodiversity Hotspot. Biocatal. Agric. Biotechnol. 2018, 15, 370–376. [Google Scholar] [CrossRef]
- Loulou, A.; Guerfali, M.M.; Muller, A.; Bhat, A.H.; Abolafia, J.; Machado, R.A.R.; Kallel, S. Potential of Oscheius tipulae Nematodes as Biological Control Agents against Ceratitis capitata. PLoS ONE 2022, 17, e0269106. [Google Scholar] [CrossRef]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive Pest of Ripening Soft Fruit Expanding Its Geographic Range and Damage Potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Loulou, A.; Mastore, M.; Caramella, S.; Bhat, A.H.; Brivio, M.F.; Machado, R.A.R.; Kallel, S. Entomopathogenic Potential of Bacteria Associated with Soil-Borne Nematodes and Insect Immune Responses to Their Infection. PLoS ONE 2023, 18, e0280675. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Bhat, A.H.; Abolafia, J.; Muller, A.; Bruno, P.; Fallet, P.; Arce, C.C.M.; Turlings, T.C.J.; Bernal, J.S.; Kajuga, J.; et al. Multi-Locus Phylogenetic Analyses Uncover Species Boundaries and Reveal the Occurrence of Two New Entomopathogenic Nematode Species, Heterorhabditis ruandica n. sp. and Heterorhabditis zacatecana n. sp. J. Nematol. 2021, 53, e2021-89. [Google Scholar] [CrossRef]
- Garriga, A.; Mastore, M.; Morton, A.; Del Pino, F.G.; Brivio, M.F. Immune Response of Drosophila suzukii Larvae to Infection with the Nematobacterial Complex Steinernema carpocapsae–Xenorhabdus nematophila. Insects 2020, 11, 210. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.N.; Lucardi, R.D.; Marsico, T.D. Successful Invasions and Failed Biocontrol: The Role of Antagonistic Species Interactions. Bioscience 2019, 69, 711–724. [Google Scholar] [CrossRef]
- Lv, C.; Huang, Y.-Z.; Luan, J.-B. Insect-microbe Symbiosis-Based Strategies Offer a New Avenue for the Management of Insect Pests and Their Transmitted Pathogens. Crop Health 2024, 2, 18. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents against Plant Diseases: Relevance beyond Efficacy. Front. Plant Sci. 2019, 19, 845. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, R.; Bankar, D.; Sahane, P.; Pushpalatha, M. Biological Control of Insects; NIPA: Houghton Le Spring, UK, 2024; pp. 61–76. ISBN 978-93-58879-78-0. [Google Scholar]
- Pylak, M.; Oszust, K.; Panek, J.; Siegieda, D.; Cybulska, J.; Zdunek, A.; Orzeł, A.; Frąc, M. Impact of Microbial-Based Biopreparations on Soil Quality, Plant Health, and Fruit Chemistry in Raspberry Cultivation. Food Chem. 2025, 462, 140943. [Google Scholar] [CrossRef] [PubMed]
- Pylak, M.; Oszust, K.; Frąc, M. Review Report on the Role of Bioproducts, Biopreparations, Biostimulants and Microbial Inoculants in Organic Production of Fruit. Rev. Environ. Sci. Biotechnol. 2019, 18, 597–616. [Google Scholar] [CrossRef]
- Pylak, M.; Oszust, K.; Frąc, M. Optimization of Growing Medium and Preservation Methods for Plant Beneficial Bacteria and Formulating a Microbial Biopreparation for Raspberry Naturalization. Agronomy 2021, 11, 2521. [Google Scholar] [CrossRef]
- Khoso, M.A.; Wagan, S.; Alam, I.; Hussain, A.; Ali, Q.; Saha, S.; Poudel, T.R.; Manghwar, H.; Liu, F. Impact of Plant Growth-Promoting Rhizobacteria (PGPR) on Plant Nutrition and Root Characteristics: Current Perspective. Plant Stress 2024, 11, 100341. [Google Scholar] [CrossRef]
- Vendan, R.T.; Yu, Y.J.; Lee, S.H.; Rhee, Y.H. Diversity of Endophytic Bacteria in Ginseng and Their Potential for Plant Growth Promotion. J. Microbiol. 2010, 48, 559–565. [Google Scholar] [CrossRef]
- Pantoja-Guerra, M.; Burkett-Cadena, M.; Cadena, J.; Dunlap, C.A.; Ramírez, C.A. Lysinibacillus spp.: An IAA-Producing Endospore Forming-Bacteria That Promotes Plant Growth. Antonie Van Leeuwenhoek Int. J. General. Mol. Microbiol. 2023, 116, 615–630. [Google Scholar] [CrossRef]
- Lekakarn, H.; Promdonkoy, B.; Boonserm, P. Interaction of Lysinibacillus sphaericus Binary Toxin with Mosquito Larval Gut Cells: Binding and Internalization. J. Invertebr. Pathol. 2015, 132, 125–131. [Google Scholar] [CrossRef]
- Dahmana, H.; Raoult, D.; Fenollar, F.; Mediannikov, O. Insecticidal Activity of Bacteria from Larvae Breeding Site with Natural Larvae Mortality: Screening of Separated Supernatant and Pellet Fractions. Pathogens 2020, 9, 486. [Google Scholar] [CrossRef]
- Aneha, K.; Bora, B.; Hemaladkshmi, P.; Manikandan, S.; Lukose, J.; Abhisubesh, V.; Gangmei, K.; Mandodan, S.; Mathivanan, A.; Vijayalakshmi, K.; et al. Lysinibacillus fusiformis: A Novel Mosquitocidal Bacterium Isolated from Western Ghats, Kerala, India. Entomon 2023, 48, 605–610. [Google Scholar] [CrossRef]
- Cossentine, J.; Robertson, M.; Xu, D. Biological Activity of Bacillus thuringiensis in Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Mastore, M.; Caramella, S.; Quadroni, S.; Brivio, M.F. Drosophila suzukii Susceptibility to the Oral Administration of Bacillus thuringiensis, Xenorhabdus nematophila and Its Secondary Metabolites. Insects 2021, 12, 635. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and Its Pesticidal Crystal Proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef]
- Berry, C. The Bacterium, Lysinibacillus sphaericus, as an Insect Pathogen. J. Invertebr. Pathol. 2012, 109, 1–10. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The Gut Microbiota of Insects—Diversity in Structure and Function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Castro, B.M.d.C.e.; Martinez, L.C.; Barbosa, S.G.; Serrão, J.E.; Wilcken, C.F.; Soares, M.A.; da Silva, A.A.; de Carvalho, A.G.; Zanuncio, J.C. Toxicity and Cytopathology Mediated by Bacillus thuringiensis in the Midgut of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Sci. Rep. 2019, 9, 6667. [Google Scholar] [CrossRef]
- Guo, W.; Kain, W.; Wang, P. Effects of Disruption of the Peritrophic Membrane on Larval Susceptibility to Bt Toxin Cry1Ac in Cabbage Loopers. J. Insect Physiol. 2019, 117, 103897. [Google Scholar] [CrossRef]
- Ramos, J.L.; Goldberg, J.B.; Filloux, A. Pseudomonas: Volume 7: New Aspects of Pseudomonas Biology; Springer: Amsterdam, The Netherlands, 2015; ISBN 9789401795555. [Google Scholar]
- Tetreau, G. Interaction between Insects, Toxins, and Bacteria: Have We Been Wrong so Far? Toxins 2018, 6, 281. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Hsieh, Y.H.; Jacobs, E.; Miranda, N.; Simpson, S.; Kerdahi, K. Identification of Lysinibacillus fusiformis Isolated from Cosmetic Samples Using MALDI-TOF MS and 16S RRNA Sequencing Methods. J. AOAC Int. 2018, 101, 1757–1762. [Google Scholar] [CrossRef] [PubMed]
- Ruffner, B.; Péchy-Tarr, M.; Ryffel, F.; Hoegger, P.; Obrist, C.; Rindlisbacher, A.; Keel, C.; Maurhofer, M. Oral insecticidal activity of plant-associated pseudomonads. Environ. Microbiol. 2013, 15, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Grizanova, E.V.; Dubovskiy, I.M.; Whitten, M.M.A.; Glupov, V.V. Contributions of Cellular and Humoral Immunity of Galleria mellonella Larvae in Defence against Oral Infection by Bacillus thuringiensis. J. Invertebr. Pathol. 2014, 119, 40–46. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastore, M.; Broggio, E.; Banfi, D.; Machado, R.A.R.; Bhat, A.H.; Kallel, S.; Reguzzoni, M.; Quadroni, S.; Brivio, M.F. Evaluation of the Insecticidal Potential of Lysinibacillus fusiformis Against Drosophila suzukii Larvae. Insects 2025, 16, 1090. https://doi.org/10.3390/insects16111090
Mastore M, Broggio E, Banfi D, Machado RAR, Bhat AH, Kallel S, Reguzzoni M, Quadroni S, Brivio MF. Evaluation of the Insecticidal Potential of Lysinibacillus fusiformis Against Drosophila suzukii Larvae. Insects. 2025; 16(11):1090. https://doi.org/10.3390/insects16111090
Chicago/Turabian StyleMastore, Maristella, Elisa Broggio, Davide Banfi, Ricardo A. R. Machado, Aashaq Hussain Bhat, Sadreddine Kallel, Marcella Reguzzoni, Silvia Quadroni, and Maurizio F. Brivio. 2025. "Evaluation of the Insecticidal Potential of Lysinibacillus fusiformis Against Drosophila suzukii Larvae" Insects 16, no. 11: 1090. https://doi.org/10.3390/insects16111090
APA StyleMastore, M., Broggio, E., Banfi, D., Machado, R. A. R., Bhat, A. H., Kallel, S., Reguzzoni, M., Quadroni, S., & Brivio, M. F. (2025). Evaluation of the Insecticidal Potential of Lysinibacillus fusiformis Against Drosophila suzukii Larvae. Insects, 16(11), 1090. https://doi.org/10.3390/insects16111090