Simple Summary
The housefly (Musca domestica) is one of the most common species in urban areas and a mechanical vector of various pathogens affecting human and animal health worldwide. To control this vector, the bacterium Bacillus thuringiensis (Bt) offers a safe alternative to chemical insecticides that is highly specific, environmentally safe, and economically viable. In this study, we evaluated the effectiveness of 12 Bt strains isolated from substrates collected in Macapá, Brazil, on the postembryonic development of housefly. Six strains caused high mortality rates (70–100%) and carried specific cry and vip genes. Scanning electron microscopy revealed diverse crystal morphologies, suggesting bioinsecticidal potential against this and other pest species. These findings demonstrate the promising potential of Brazilian Bt strains for developing bioinsecticides to manage this muscoid dipteran.
Abstract
The high pathogenic activity of Bacillus thuringiensis (Bt) strains against various insect orders has positioned it as the most effective, environmentally safe, and sustainable approach to integrated insect management. We aimed to identify Bt strains capable of effectively controlling the housefly Musca domestica, a species of significant medical, veterinary, and public health concern. Twelve Bt strains from Macapá, Brazil, were tested against housefly larvae. PCR was employed to detect genes encoding Cry and Cyt proteins and Vips. Six strains exhibited 70 to 100% larval mortality, with five containing cry genes, none harboring cyt genes, and four carrying vip genes. Scanning electron microscopy revealed the production of crystal inclusions with distinct morphologies: spherical (TR4J, SOL5DM, SOL6RN), cuboidal and bipyramidal (TRO1TN and TRO2MQ), and spherical and bipyramidal (UNI2MA). The potential presence of genes from the cry1, cry2, vip1, and vip3 families suggests that these strains also exhibit bioinsecticidal activity against other muscoid flies and insect pests across various orders. This study underscores the bioactive potential of these Brazilian Bt strains for developing new bioinsecticides.
1. Introduction
Musca domestica (Linnaeus, 1758) (Diptera: Muscidae), commonly known as the housefly, is a highly synanthropic muscoid with considerable economic, medical, veterinary, and cosmopolitan importance. It is a recognized mechanical vector for over 60 types of foodborne pathogens and serious diseases, including diarrhea, anthrax, typhoid fever, tuberculosis, and cholera [1,2]. Cases of secondary myiasis caused by M. domestica have been documented but remain under-reported in the scientific literature, as noted by Dogra and Mahajan [3] and Salem and Attia [4]. These findings underscore the importance of further research into the bionomics, intrapuparial development, and effective population management of this species [5,6].
Pathogen transmission by M. domestica is facilitated by its sucking–licking mouthparts [7]. During feeding, the housefly regurgitates saliva and small droplets from its digestive tract to dissolve the material it consumes while defecating, contaminating food substrates [8].However, pathogen spread is not restricted to regurgitation and defecation [9]. The mouthparts, wings, and leg extremities of this fly also serve as structures for transporting helminth eggs and larvae, further increasing contamination risks [10]. These combined behaviors play a significant role in facilitating disease transmission [11].
The preference of the housefly for diverse substrates—ranging from human and animal waste to decaying plant matter—for its nutritional and reproductive needs plays a critical role in the dissemination of bacterial strains worldwide. This includes species of Klebsiella (Enterobacterales; Enterobacteriaceae) which are known to harbor antibiotic-resistant genes [12,13]. Such behavior has significantly contributed to the global proliferation of these pathogens, establishing M. domestica as one of the primary pests of medical and veterinary importance [14].
Currently, the control of M. domestica predominantly relies on chemical pesticides, many of which fail to adhere to basic biosafety principles. However, the emergence of insecticide resistance and growing environmental concerns about human and animal health have intensified the demand for safer and more sustainable control strategies within the framework of integrated pest and vector management [15,16]. In this context, microbial biological control has emerged as a self-sustaining and effective technique. It has demonstrated efficacy not only in controlling agricultural pests but also in reducing populations of insect vectors, such as flies and mosquitoes [17]. This method leverages pathogenic microorganisms, offering a highly specific and environmentally benign solution. Its advantages include ease of microorganism multiplication, efficient pathogen dissemination, reduced dependence on chemical pesticides, lower rates of insect resistance, and minimized environmental impact and toxicity to human and animal health [18]. Aligned with this approach, the biological agent Bacillus thuringiensis (Bt) Berliner, 1915 (Caryophanales: Bacillaceae) emerges as a viable alternative for controlling various insect pests and vectors [19].
Bt is an aerobic, Gram-positive bacterium that produces crystalline protein inclusions, known as Cry and Cyt proteins or delta (δ) endotoxins, during the stationary phase. Additionally, Bt strains synthesize other insecticidal proteins, including vegetative insecticidal proteins (Vips) and secreted insecticidal proteins (Sips) [20,21,22]. Cry proteins are active against lepidopterans, coleopterans, hymenopterans, and dipterans, whereas Cyt proteins exhibit specific toxicity to dipterans in vivo [23]. Despite sharing biochemical properties, Cry and Cyt toxins are neither phylogenetically related nor structurally homologous [24]. These proteins are synthesized during the sporulation phase, with Cyt proteins functioning as cytolytic toxins. Cyt proteins are predominantly produced by B. thuringiensis subsp. israelensis, as well as by B. thuringiensis subsp. kyushuensis, B. thuringiensis subsp. jegathesan, and B. thuringiensis subsp. medellin, among others [25,26]. Unlike many Cry proteins, Vips target a broader range of insect species, particularly lepidopterans [27]. The nomenclature of Vips differs from that of Cry proteins, as Vips do not form protein crystals [21]. The genes responsible for Vip synthesis are located on high-molecular-weight plasmids, coexisting with cry genes [28].
The entomopathogenic activity of Bt occurs through the formation of crystals during the sporulation phase, which is driven by protein accumulation. When ingested by specific insects, these crystals release proteins that are activated in their midguts. The activated proteins interact with the epithelial cells, creating pores in the cell membrane. These pores cause disturbances in the insects’ bodies, ultimately leading to their death [29].
Although several Bt-based products are available, including VectoBac, VectoLex, VectoPrime, VectoMax, ReMoa Tri, Bactimos (Valent BioSciences Corporation, Libertyville, IL, USA), Teknar (Syngenta, Basel, Switzerland), Aquabac (Becker Microbial Products, Lake Worth, FL, USA), LarvX SG (Meridian Precision Release Technologies, Dalia, Israel), Culinex (Culinex GmbH, Seattle, WA, USA), Bacticide (Biotech International Ltd., Noida, India), Bactivec (Labiofam, La Habana, Cuba), and Bt-Horus (Bthek Biotecnologia, Brasília, Brazil), much work remains in regard to addressing a broader range of dipteran vectors, particularly muscoids. These vectors, including species commonly found in urban and rural environments, transmit pathogens of significant public health concern [30,31].
As research advances, it is critical to focus on deploying existing registered and commercialized biopesticides and identifying new Bt strains. Particular emphasis should be placed on their molecular characterization rather than relying solely on flagellar antigen H (subspecies) classification [32,33]. This molecular approach is crucial for integrated vector management, as identifying the most effective Bt strains for specific vectors, such as M. domestica, could significantly reduce dependence on chemical pesticides. Additionally, it plays a key role in insect resistance management and strengthens long-term vector control programs [34,35].
Recognizing the importance of this low-impact system for health applications and harnessing Brazil’s vast biodiversity, this study focused on selecting and characterizing Bt strains from Macapá, Amapá State. We assessed their potential for controlling M. domestica by investigating their novel insecticidal properties.
2. Materials and Methods
This study was conducted at the Integrated Laboratory: Simulids and Oncocercosis & Medical and Forensic Entomology (LSOEMF/IOC), in partnership with the Biological Control Laboratory at EMBRAPA Maize and Sorghum, Sete Lagoas, Minas Gerais, Brazil. We followed the methodology proposed by Merdan [36], with modifications.
2.1. M. domestica Colonies
Wild housefly adults were actively collected every two weeks during morning sessions from garbage bins in the Amorim Community (22°52′32″ S; 43°15′02″ W; altitude: 25 m), situated near the entrance to the Oswaldo Cruz Foundation (FIOCRUZ) campus at Rua Sizenando Nabuco, No. 100, Manguinhos, Rio de Janeiro, Brazil. After collection, the flies were transported in an appropriate isothermal box to the LSOEMF/IOC for subsequent identification.
The establishment and maintenance of M. domestica colonies followed the procedures described by Nascimento et al. [37].
2.2. Bt Strains and Growth Conditions
The Bt strains TRO1TN, TRO2MQ, TA5FV, TA1IC, VG1MD, VG2NN, TOR1KC, TOR2VN, UNI2MA, TRAJ, SOL5DM, and SOL6RN were previously collected from various sources in Macapá, including soil samples, organic plant materials, tree trunks, and spider webs (Table 1). These strains, which do not exhibit β-exotoxin pathogenic activity, have been maintained at the EMBRAPA Maize and Sorghum Microorganism Bank and were obtained by the authors for this research. Prior to this study, they were tested exclusively on agricultural pests, specifically Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae), Helicoverpa armigera (Hübner, 1808) (Lepidoptera: Noctuidae), and Chrysodeixis includens (Walker, 1858) (Lepidoptera: Noctuidae).

Table 1.
Bacillus thuringiensis strains and their respective substrate samples collected in Macapá, Amapá, Brazilian Amazon.
Each strain was cultured on commercial Luria–Bertani (LB) medium supplemented with mineral salts (0.002 g FeSO4, 0.02 g ZnSO4, 0.02 g MnSO4, and 0.3 g MgSO4) in Petri dishes. The cultures were incubated at 29 °C for 72 h in a bacteriological incubator to induce sporulation and crystal release. The bacterial biomass was harvested, suspended in autoclaved deionized water in Falcon tubes, and diluted with a 0.05% Tween-20 emulsifier. Spores were then counted using a Neubauer chamber with a phase-contrast optical microscope, Zeiss Axioskop (Oberkochen, Germany).
2.3. Selective Bioassays on M. domestica
The experimental design was completely randomized, with four replicates per treatment. Forty neolarvae were placed into 300 mL glass containers (canning jars) containing 80 g of a diet comprising meat meal and wheat bran in a 3:1 ratio. To minimize contamination risk, the diet was exposed to UV radiation in a biological safety cabinet for 20 min prior to use in the bioassays. Subsequently, 4 mL of bacterial solution (108 spore–crystal/mL) from the strains TRO1TN, TRO2MQ, TA5FV, TA1IC, VG1MD, VG2NN, TOR1KC, TOR2VN, UNI2MA, TRAJ, SOL5DM, and SOL6RN was added to each treatment. The control group received 4 mL of autoclaved distilled water. Bioassays were performed in a B.O.D. incubator set to 27 ± 1 °C, with relative humidity being maintained at 70% and a 12 h photoperiod. Mortality rates were assessed 72 h after the initiation of the bioassay, focusing on the postembryonic development of M. domestica.
2.4. Data Analysis
The data underwent an analysis of variance (ANOVA; p ≤ 0.05) followed by factorial analysis using the Scott–Knott test (p > 0.05), conducted using the statistical software Sisvar Version 5.6 [38].
2.5. DNA Extraction and Molecular Characterization
The Bt strains were cultured in an LB medium at 29 °C for 16 h. Subsequently, genomic DNA was extracted using the Wizard® Genomic DNA Purification Kit (Promega Corp., Madison, WI, USA), according to the manufacturer’s instructions. The cry, cyt, and vip genes were detected through PCR reactions using specific primers described in the literature [39,40,41,42,43,44,45] (Table 2).
Table 2.
Primers used for genomic DNA amplification of Bacillus thuringiensis strains.
Each PCR reaction contained 10 ng of DNA, 0.5 µM of each primer, 5 µM of each dNTP, 1× buffer solution, 2 mM MgCl2, and 2U of Taq polymerase (KAPA Biosystems, Wilmington, MA, USA) in a total volume of 10 μL. The amplification was performed in a Veriti® 96-Well thermal cycler under the following conditions: initial denaturation at 94 °C for 5 min, 35 cycles of denaturation at 95 °C for one minute, annealing at primer-specific temperatures for one minute, extension at 72 °C for one minute, and final extension at 72 °C for 10 min. The PCR products were analyzed by electrophoresis on a 1% agarose gel using the molecular weight marker 1kb plus (Invitrogen, Carlsbad, CA, USA) for band comparison.
2.6. Scanning Electron Microscopy
Scanning electron microscopy (SEM) analysis was performed to examine the morphology and dimensions of crystals found in Bt strains that caused the highest average mortality rates in M. domestica. The methodology followed the protocols described by Valicente and Souza [46]. The Bt strains were cultured on nutrient agar, adjusted to a final pH of 7.5, and incubated at 30 °C for 72 h to ensure complete sporulation. The presence of the spore–crystal complex was then confirmed using a phase-contrast microscope (Zeiss Axioskop, Oberkochen, Germany) at 1000× magnification.
After incubation, the bacterial content was scraped from Petri dishes, and sporulating cells were isolated by centrifugation at 14,000 rpm for 5 min using a specific microtube rotor. The bacterial pellets were then fixed in 2.5% glutaraldehyde in 0.01 M sodium cacodylate buffer for 1 h at room temperature. Subsequently, the samples were washed three times in 0.01 M sodium cacodylate buffer and immersed in 0.5% osmium tetroxide for 1 h. After post-fixation, the samples were dehydrated through a series of ethanol concentrations (25, 50, 75, 90, and 100%), with two immersions at each concentration for 20 min. The dehydrated samples were critical point-dried, mounted on poly-L-lysine-coated coverslips for 48 h, and placed in a tissue culture plate. The samples were then gold sputter-coated and observed using a Jeol JSM-6390LV scanning electron microscope (Garden, UK) on the electron microscopy platform. Micrographs were saved onto CD-RW media for further analysis.
3. Results
3.1. Bt Virulence
The selective bioassay showed that all 12 Bt strains tested exhibited pathogenic activity against M. domestica larvae. Notably, strains TR4J, SOL5DM, and SOL6RN achieved a mortality rate of 100%, followed by TRO1TN (98%), UNI2MA (97%), and TRO2MQ (93%). The remaining strains displayed mortality rates ranging from 33% to 54%. The control group recorded a mortality rate of 10% (Figure 1). Figure 2 illustrates the average pupal weight values for M. domestica across the different experimental groups. Larvae treated with the UNI2MA, TRO1TN, and TROMQ strains exhibited the lowest average pupal weight gains, with values of 0.7 mg, 1.22 mg, and 1.25 mg, respectively. Insects treated with the TA5FV, VG2NN, TA1IC, VG1MD, TOR2VN, and TOR1KC strains also demonstrated a biologically significant reduction in pupal weight, with averages below 14 mg. The control group had an average pupal biomass of 20 mg.
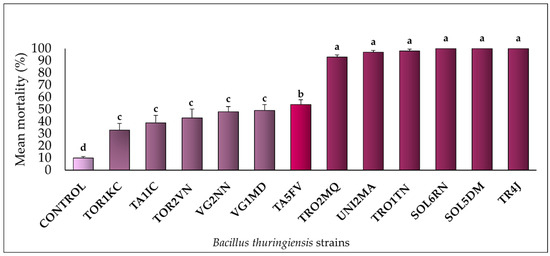
Figure 1.
Mortality rates (%) of Musca domestica (Diptera: Muscidae) larvae in a selective bioassay with Bt strains at a concentration of 108 spores/mL, added to a diet of putrefied ground beef, along with the negative control (autoclaved distilled water). For each treatment, averages marked with the same letter and corresponding to columns of the same color did not differ significantly according to the Scott–Knott test (p ≥ 0.05).
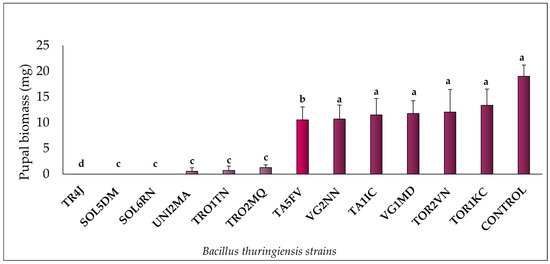
Figure 2.
Pupal biomass rates (%) of Musca domestica (Diptera: Muscidae) larvae in a selective bioassay with Bt strains at a concentration of 108 spores/mL, added to a diet of putrefied ground beef, along with the negative control (autoclaved distilled water). For each treatment, averages marked with the same letter and corresponding to columns of the same color did not differ significantly according to the Scott–Knott test (p ≥ 0.05).
Table 3 shows the significant effect on the duration of postembryonic development of M. domestica across different treatments. The larval stage lasted an average of 1.5 to 3.0 days in larvae treated with the 1641, TRO1TN, and UNI2MA strains, indicating a reduction in development time compared to other treatments, where the larval stage averaged six to seven days. A similar trend was observed in the pupal stage, with pupae from larvae fed with Bt spore/crystal suspensions of the TRO1TN and UNI2MA strains exhibiting shorter periods than those in other treatments. In contrast, although significant, the deformity rate among emerging adults was below 5%.
Table 3.
Time (in days) of postembryonic development period of Musca domestica (Diptera: Muscidae) larvae fed a substrate based on meat and bone meal and wheat bran, incorporated with Bacillus thuringiensis strains at 108 spores/mL, in comparison with the control group under laboratory conditions.
We also observed that surviving larvae, which developed on substrates containing Bt spores/crystals (108 spores/mL) from the tested strains, took less time to develop and varied in male-to-female ratio rates. The results show a trend toward reduced average postembryonic development time in specimens fed with meat containing Bt spore/crystal suspensions of the TRO1TN and UNI2MA strains. Additionally, macroscopic morphological changes were observed in M. domestica larvae treated with Bt, contrasting with the control group. These changes included variations in coloration, with a darker tone, and alterations in larval tissue texture, which appeared softer and more flaccid compared to the control group (Figure 3). Furthermore, approximately 12 to 24 h after treatment, most infected larvae ceased feeding and gradually abandoned their diet (Figure 4).
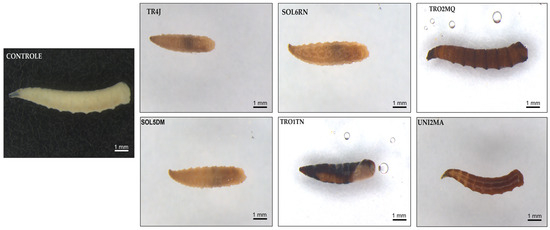
Figure 3.
External symptoms of mortality in Musca domestica larvae infected by Bacillus thuringiensis (108 spores/mL). Control: healthy larvae, with a cream/whitish body color. TR4J, SOL5DM, and SOL6RN: larvae exhibiting partially curved bodies, reduced sizes, darkened abdomens (black coloration), and flaccid body textures. TRO1TN: larva displaying color changes (darkening) over the entire integument, with a necrotic appearance and reduced, curved bodies. TRO2MQ and UNI2MA: larvae showing body darkening (brown and black) and flaccid body textures.

Figure 4.
Pathogenicity of Bacillus thuringiensis strains in Musca domestica larvae. White arrow: M. domestica immatures from the control group feeding on the diet. Purple circle: dead M. domestica larvae treated with the SOL5DM strain, positioned at the side of the container, indicating abandonment of the diet, with dark-brown coloration on the teguments.
3.2. Detection of cry, cyt, and vip Genes
After thoroughly analyzing six Bt strains known for their effectiveness against M. domestica (SOL5DM, SOL6RN, TRO2MQ TRO1MQ, TRO1TN, TR4J), we found that SOL5DM and SOL6RN amplified cry2Aa. TRO2MQ exclusively carried cry2Ac, while TRO1MQ and SOL5DM contained cry2Ad. Additionally, TRO1TN and TRO1MQ showed the presence of cry1B, and TR4J specifically amplified cry9A. However, cry1Aa, cry1Ab, cry1C, cry1G, cry1Ea, cry9B, cyt1, cyt2, and vip2 were not detected in any of the analyzed strains.
The vip1 gene was amplified in three strains: TRO1TN, TRO2MQ, and UNI2MA. Conversely, a larger number of strains, including TRO1TN, TRO2MQ, UNI2MA, and TR4J, was amplified for vip3. Table 4 summarizes the results, detailing the presence or absence of these amplifications.
Table 4.
Molecular characterization of Bacillus thuringiensis strains that are effective against Musca domestica (Diptera: Muscidae) for the presence of selected cry, cyt, and vip genes.
Molecular analysis revealed that vip3 was amplified in 67% of the six strains evaluated, followed by vip1, which was detected in 50%. The cry1B, cry2Aa, and cry2Ad genes were amplified in 33% of the strains, while cry9A and cry2Ac were detected in 17%. Notably, cry1Aa, cry1Ab, cry1C, cry1G, cry1Ea, cry9B, cyt1, cyt2, and vip2 were not identified in any of the strains analyzed.
3.3. SEM Analysis of Crystalline Inclusions of Bt Strains
SEM analysis revealed that the TR4J, SOL5DM, and SOL6RN strains exclusively produced spherical crystals. In contrast, TRO1TN and TRO2MQ produced a variety of crystal shapes, including spherical, cuboidal, and bipyramidal forms. UNI2MA, on the other hand, exhibited both spherical and bipyramidal crystals. Figure 5 presents the SEM images of crystal morphologies of the different Bt strains.

Figure 5.
Scanning electron microscopy of the spore–crystal complex of the Bacillus thuringiensis strains that showed an average mortality rate of over 70% in Musca domestica (Diptera: Muscidae) larvae. ep: spore; cb: bipyramidal crystal; ce: spherical crystal; cc: cuboid crystal (5000× magnification).
4. Discussion
Bt is a bacterium commonly found in diverse habitats, including soil, water, litter, grain, deceased or diseased insects, spider webs, and other debris [47,48]. The diversity of cry, cyt, and vip genes in the Bt genome may be influenced by the environmental factors present at the collection sites and substrates, as suggested by Valicente and Barreto [49] and Djenane et al. [50]. The strains analyzed in this study were isolated from various substrates, including soil, tree trunks, and spider webs.
The TR4J strain was isolated from a spider web near the Matapí River, while TRO1TN and TRO2MQ were isolated from tree trunks in the Matapí River region. UNI2MA was isolated from soil on the campus of the Federal University of Amapá. Previous studies [50,51,52,53] have shown that analyzing Bt strains from diverse ecological and geographic sources results in a weak correlation between their genetic content and the mortality of target insects. This finding highlights the need for further investigation to better understand the broad entomocidal activity of Bt strains [54].
The most frequent amplicons were the vip3 genes, present in four out of six strains (67%), followed by vip1 in 50% and the cry1B, cry2Aa, and cry2Ad genes, which were amplified in 33% of the strains. These findings are consistent with those of Seifinejad et al. [55], who, in their molecular characterization of 57 strains, identified the presence of vip genes in 82.6% of them.
In a survey of insecticidal toxins in 125 Bt strains, Espinasse et al. [56] observed the presence of vip1 and vip2 in approximately 34.5% of the strains and vip3 in 52.8%. Similarly, Nascimento et al. [57], in a study focused on the selection and characterization of Bt strains against lepidopteran pests, recorded a higher frequency of vip1 (75%), vip2 (75%), and vip3 (67%) in the selected strains.
Considering the higher frequency of vip genes in the strains of our study, the efficacy of the Bt crystal protein complex may result from the combination of cry genes and the insecticidal synergy between Cry proteins and Vips [58]. The coexistence of vip with cry genes was observed in the TR4J, TRO1TN, and TRO2MQ strains (Table 3), which is consistent with previous studies [59]. The correlation between the occurrence of cry1, cry2, and vip was previously reported by Hernandez et al. [42] during the identification and classification of cry and vip genes in a collection of 507 Bt strains from Spain and Bolivia.
Previous studies, such as those by Wang et al. [60], investigated the interaction between Bt toxins Cry9A and Vip3Aa, revealing a strong affinity between these proteins and highlighting their insecticidal synergy, which contributed to high activity against the Asian rice borer. In contrast, Wang et al. [61] found no evidence of synergy between three Cry proteins and Vip3Aa. However, they observed that Vip3Aa had toxic effects on the dipteran Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae), challenging the earlier assumption that Vip3Aa’s insecticidal activity was limited to lepidopteran noctuids.
The high mortality rates observed for M. domestica in this study reinforce these findings and highlight the potential of Vips as effective alternatives for controlling pest insects and disease vectors. Moreover, since Vips do not share any sequence or structural homology with Cry proteins, they represent a promising tool for preventing and/or delaying the development of insecticide resistance [62]. Consequently, these findings encourage further investigation into the mechanisms of action of vip genes, which remain poorly understood, and emphasize the potential of Vips to expand the action spectrum of Bt-based products in controlling economically and health-threatening pests worldwide [63].
Our findings align with those of Uribe et al. [52], who characterized a collection of Bt strains from various agricultural and wild ecosystems in Colombia. The authors found that 73% of the analyzed strains reacted with universal primers for the cry1 genes, highlighting cry1B as one of the most common. Similarly, Valicente et al. [43] identified cry1B as one of the most frequent genes among the 165 characterized Bt strains. The detection of cry1B in the TRO1TN and TRO2MQ strains, which demonstrated potential efficacy against M. domestica, supports previous evidence that, like other Bt proteins, Cry1B is toxic to lepidopterans, dipterans, and coleopterans. It is worth noting that this specific toxicity may be influenced by variations in the insect midgut environment and the interactions between the toxin and its receptors [64].
It is important to note that, while the Cry1 family is generally effective against lepidopteran pests, several of its proteins also exhibit pathogenicity against dipterans [65]. Previous studies [66,67,68] have demonstrated that proteins like Cry1Ab are effective against Ae. aegypti larvae; Cry1Ac targets adult Glossina morsitans Westwood, 1851 (Diptera: Glossinidae); Cry1Ba shows larvicidal activity against M. domestica, Chrysomya albiceps (Wiedemann, 1819) (Diptera: Calliphoridae), and Lucilia cuprina (Wiedemann, 1830) (Diptera: Calliphoridae); and Cry1Ca is toxic to larvae of various mosquito species. The toxicity bioassays and molecular characterization of Bt strains conducted in this study revealed the production of some of these Cry proteins with toxic activity against dipterans. These findings highlight the potential of these strains in controlling muscoids, especially considering the limited availability of effective pesticidal proteins and commercial products for fly management.
The profile of the cry2A gene from the Bt strains analyzed in this study revealed that cry2Aa and cry2Ad were the most prevalent genes, occurring in 33% of the analyzed strains, while cry2Ac was detected in only one strain (17%). This distribution of cry2 genes aligns with the findings of Mendoza et al. [69]. They characterized 28 Bt strains isolated from the Tijuana–Ensenada region in northwestern Mexico and found a predominance of cry2Aa (71%) and cry2Ac (14%).
Van Frankenhuyzen [70] and Alzahrani and Crickmore [71] found that the Cry2Aa protein exhibits activity against lepidopterans, dipterans, and hemipterans, while Cry2Ac is effective against lepidopterans and dipterans. In contrast, Cry2Ad is specific to lepidopterans. Considering the action spectrum of Cry2A proteins and their binding site specificity relative to Cry1 family proteins [72], the toxicity assays and cry2 distribution in the Bt strains that we analyzed provide valuable insights into the lethal effects of these proteins on M. domestica.
Most PCR reactions yielded the anticipated products, except for those using specific primers targeting cry1Aa, cry1Ab, cry1C, cry1G, cry1Ea, cry9B, cyt1 cyt2, and vip2, which failed to amplify. This suggests that these genes are not responsible for the toxicity of the strains against M. domestica. These findings align with those of Van Frankenhuyzen [70] and Polanczyk et al. [73], which showed that, out of 72 proteins tested against Diptera, only 42 demonstrated activity, indicating that pure proteins encoded by some of these genes did not induce significant mortality in M. domestica during bioassays. Additionally, gene frequency variation may be influenced by environmental factors, such as the different locations and substrates from which the strains were collected, which could impact the distribution of cry, cyt, and vip genes [74].
Collections of Bt strains encompass a variety of cry, cyt, and vip genes arranged in diverse combinations within their genomes [55,75]. However, it is crucial to emphasize that the mere presence of a specific gene does not necessarily indicate toxic activity [76]. This discrepancy arises because gene presence does ensure bacterial expression [77]; some genes may remain inactive or may be expressed at low levels, reducing their entomopathogenic potential. Nonetheless, identifying the types of genes within the strains is essential for establishing correlations between their genetic profiles and bioinsecticidal efficacy [41].
The pathogenicity of Cry proteins is directly associated with the formation of protein inclusions during Bt sporulation, which is triggered by adverse environmental conditions [29,78]. The composition of proteins like Cry and Cyt dictates the morphology of the resulting crystals, demonstrating a strong correlation between crystal structure and protein composition.
For example, bipyramidal crystals are often linked to the presence of Cry1 and/or Cry9 proteins [79,80]. In contrast, cuboidal and spherical inclusions are commonly associated with Cry2 proteins and typically coexist with bipyramidal crystals [81]. These structural characteristics are crucial for the entomopathogenic effectiveness of Bt strains, as crystal morphology can influence their interaction with insect gut cells, directly impacting their virulence [82].
Palma et al. [83] and Nair et al. [79] reported that many Bt strains feature parasporal crystal proteins that are pivotal to their insecticidal action, with their morphology varying based on the type of the δ-endotoxins and their corresponding genes. In our investigation, only spherical Bt crystals were identified in the TR4J, SOL5DM, and SOL6RN strains, all exhibiting 100% mortality against M. domestica. This observation aligns with the findings of Loutfi et al. [84], who also identified spherical crystals in Bt strains that were effective against dipterans.
When assessing pathogenic activity and characterizing Brazilian Bt strains against lepidopteran and dipteran pests, Gitahy et al. [85] and Arsov et al. [86] identified efficient strains that also produced spherical, bipyramidal, and cuboidal crystals along with genes from the cry1 and cry2 families that were similar to those observed in the Bt strains analyzed in our study. These findings highlight that Bt strains with these characteristics exhibit entomopathogenic activity across diverse insect orders, underscoring their potential for developing new biopesticide products.
The housefly’s rapid development, high reproductive capacity, mobility, ability to exploit diverse substrates for reproduction, adaptability to different environments, and propensity to develop resistances to new insecticides pose significant challenges to its control [87,88].
Our findings revealed the genetic diversity among the analyzed Bt strains, offering key perspectives on the variability of their entomopathogenic activity and their potential for managing M. domestica. Soares-da-Silva et al. [89] also identified Brazilian Bt strains pathogenic to Ae. aegypti, the vector for several arboviruses. This emphasizes the importance of exploring new strains that may provide novel toxin combinations for the biological control of insect vectors, as demonstrated in our study on houseflies.
The results indicate a trend toward a reduced average time of postembryonic development in housefly specimens that feed on meat containing spore/crystal suspensions of the Bt strains TRO1TN and UNI2MA. This reduction may be attributed to the entomopathogenic action of Bt on susceptible M. domestica larvae. The larvae exhibited symptoms such as loss of appetite and abandonment of their diet, likely due to discomfort or intestinal paralysis. This paralysis may result from the hydrolysis of the crystals in the midgut, which releases active toxins (crystals). These toxins can affect the intestinal epithelium and potentially spread to other body cavities, affecting various organs and systems in the insect.
According to Sebesta et al. [90], the larval stage is always more susceptible to Bt than adults, with sublethal doses causing anomalies, deformities, and teratological changes during critical metamorphosis stages. Similar observations were made in the present study, where preliminary laboratory tests suggested that M. domestica is susceptible to the tested Bt suspensions.
However, further investigations into the external symptoms and histological changes in specimens infected by this pathogen are necessary. A more comprehensive approach will enable a detailed characterization of the effects of these Bt strains on M. domestica and may help guide more precise and sustainable control strategies for this fly.
Indeed, most screening studies on Bt strains prioritize lethal action, focusing primarily on its immediate impact on insects. However, it is equally important to assess sublethal effects, which, although not causing obvious mortality, can significantly influence the dynamics of the target insect [91]. Considering sublethal effects is crucial for integrated pest management programs, as not all field applications result in target-insect mortality. Even products with limited efficacy can affect fundamental biological parameters, such as fecundity, longevity, development rate, and sex ratio [92]. In this context, the present study highlights the variation in the insecticidal potential of bacterial suspensions of this pathogen when applied against M. domestica.
Body mass estimation is a crucial parameter in evaluating biological agents as candidates for controlling pests and vector insects. According to Hanski [93], Davidowitz et al. [94], and Chapman et al. [95], the relative success of each insect species largely depends on the larvae’s ability to reach the minimum body weight required for viable pupation. These studies suggest that forming viable pupae with comparatively lower body masses is a strategy for mitigating the adverse effects of competition and/or food restrictions.
Conversely, other researchers, such as Ullyett [96] and Goodbrod and Goff [97], have argued that some muscoids are better adapted to pupate even when their final body mass falls below pre-established standard values compared to other species. Furthermore, both a significant reduction and increase in body mass can be detrimental to an insect’s developmental, sexual, and reproductive performance, depending on the context. In this regard, our study suggests that the sublethal effects of Bt exposure led to a reduction in body mass in M. domestica—a significant biological change that could considerably impact the species’ viability.
Panizzi and Parra [98] emphasized the importance of examining various aspects during the pupal stage, including the duration of the pupal period, the weight of pupae at a specific age, pupal viability, sex ratio, and the occurrence of body deformities, emphasizing the close correlation between pupal weight and reproductive capacity. They also noted that pupae lose water over time, which justifies the need for weighing them at fixed intervals, such as 24 or 48 h after pupation.
Based on these guidelines, the present study suggests that the ingestion and utilization of the substrate by M. domestica larvae exposed to Bt spores/crystals were compromised. The variation in the effectiveness of the tested strains can be attributed to a range of factors that are both related and unrelated to the Bt mode of action. These factors include the dissolution of the crystal, the activation of the protein, and the binding of the activated protein to receptors in the intestinal epithelium. Consequently, both the amount of food consumed by the larvae and the intestinal dysfunction caused by Bt may have interfered with the development time and pupal biomass of the target dipteran.
Miranda et al. [99] evaluated the large-scale larval performance of M. domestica using 4000 larvae and 1 kg of different diets based on swine, dairy cattle, and poultry manure, along with a control group fed the Gainesville diet, which consisted of wheat bran, alfalfa meal, and cornmeal. They found that peak larval weight was reached four days after inoculating the larvae with the diet, with no significant differences being seen in the development time to first pupation across the diets. However, the survival rate to pupation varied considerably, with the highest survival rates being seen in the Gainesville (74%), swine (73%), and poultry (67%) diets and the lowest rates being seen in the dairy cattle manure diet (50%). Larval weight also varied between the diets, with larvae fed with swine, dairy cattle, and poultry manure averaging 21 mg, 24 mg, and 25 mg, respectively, while the control group had an average weight of approximately 27 mg.
When comparing these results with those of the present study, which was conducted on a smaller scale (40 larvae fed with 80 g of meat meal and wheat flour per treatment), we noted that the average pupal weight in the control group (19 mg) was adequate for the development of M. domestica, maintaining the specimens’ viability throughout the evaluation period. However, it was evident that the larval feeding response after ingesting a diet containing Bt negatively impacted their development, leading to a significant reduction in pupal biomass rates for this fly, as shown in Table 2.
Fisher [100] and Norris [101] observed that a stable population typically had a male-to-female ratio of 1:1 or slightly more females than males, which helped ensure the species’ perpetuation. Building on these observations, it is advisable to assess other biological parameters, such as the pre-oviposition period, daily mortality rates for both sexes, and daily egg-laying capacity. These parameters could offer further insights into the impact of Bt exposure on the biology and population dynamics of this fly.
Despite the growing interest in prospecting new Bt strains, a significant gap remains in the literature regarding their bioactive potential against insect pests and vectors, as well as the characterization of Bt strains from the state of Amapá in the Brazilian Amazon, particularly in regard to the content of the cry and vip genes. In this study, we provide important information on Bt strains isolated from samples collected in the city of Macapá, emphasizing the need to perform additional sampling of this species, particularly across different Brazilian states.
Macapá houses four terrestrial ecosystems—forest, cerrado, floodable fields, and mangroves—along with a rich aquatic and estuarine biota. This vast biodiversity offers a unique opportunity for isolating Bt strains with promising biological properties [102]. Therefore, exploring these strains is crucial for developing biological pest control strategies, identifying specific entomopathogenic activities, and discovering new approaches to managing local insect pests. The efficacy of the strains TR4J, SOL5DM, SOL6RN, TRO1TN, TRO2MQ, and UNI2MA against M. domestica underscores the importance of investigating their potential against other species of flies and pests. Further assessments are needed, as the proteins present in these Bt strains may pose risks to insects of medical relevance in both sanitary and veterinary scenarios.
Author Contributions
Conceptualization, T.A.N. and M.M.d.C.Q.; methodology, T.A.N. and M.J.P.; software, T.A.N.; validation, M.M.d.C.Q. and F.H.V.; formal analysis, T.A.N. and M.J.P.; investigation, T.A.N., M.J.P. and M.M.d.C.Q.; data curation, T.A.N. and M.J.P.; writing—original draft preparation, T.A.N.; writing—review, T.A.N., M.J.P., F.H.V. and M.M.d.C.Q.; supervision, M.M.d.C.Q. and F.H.V.; project administration, M.M.d.C.Q., funding acquisition, M.M.d.C.Q. This work was carried out in collaboration between all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funds from PAEF-IOC/FIOTEC (Strategic Actions for Development and Strengthening Accredited Laboratories and Research Support Areas) from The Oswaldo Cruz Foundation (FIOCRUZ) (Process Identification: IOC-023-FIO-18-2-30), Coordination for the Improvement of Higher Education Personnel (CAPES) under Financial Code 001, The National Council of Research and Technological Development—CNPq (Process number: 316.254/2021-5), and The Carlos Chagas Filho Research Support Foundation of the State of Rio de Janeiro—FAPERJ (Process Identification: E-26/210.228/2018; E-26/210.982/2021; E-26/200.471/2023), and VPPCB/Fundação Oswaldo Cruz.
Data Availability Statement
The data that support the findings of this study are openly available in Mendeley Data at https://data.mendeley.com/datasets/rcyt27txr9/1, accessed on 20 June 2024. DOI: 10.17632/rcyt27txr9.1.
Acknowledgments
The authors gratefully acknowledge the financial support provided by the funding agencies and colleagues from laboratories the for their support. Finally, they acknowledge the collaboration of the Rudolf Barth Electron Microscopy Platform at the Oswaldo Cruz Institute, especially Roger Magno, Wendell Girard-Dias and José Lopes.
Conflicts of Interest
The authors declare no conflicts of interest.
Correction Statement
This article has been republished with a minor correction to the Funding statement. This change does not affect the scientific content of the article.
References
- Pitkin, A.; Deen, J.; Otake, S.; Moon, R.; Dee, S. Further assessment of houseflies Musca domestica as vectors for the mechanical transport and transmission of porcine reproductive and respiratory syndrome virus under field conditions. Can. J. Vet. Res. 2009, 73, 91–96. [Google Scholar] [PubMed]
- Baker, S.Z.; Atiyae, Q.M.; Khairallah, M.S. Isolation and Identification of some Species of Bacterial Pathogens from Musca Domestica and Test their Susceptibility Againts Antibiotics. Tikrit J. Pure Sci. 2018, 23, 20–27. [Google Scholar] [CrossRef]
- Dogra, S.S.; Mahajan, V.K. Oral myiasis caused by Musca domestica larvae in a child. Int. J. Pediatr. Otorhinolaryngol. 2010, 5, 105–107. [Google Scholar] [CrossRef]
- Salem, H.M.; Attia, M.M. Accidental intestinal myiasis caused by Musca domestica L. (Diptera: Muscidae) larvae in broiler chickens: A field study. Int. J. Trop. Insect Sci. 2021, 41, 2549–2554. [Google Scholar] [CrossRef]
- Abellán, M.; Ruano, F.; Rojo, S.; Martínez-Sánchez, A. Effect of two larval diets on the reproductive parameters of the housefly, Musca domestica L. (Diptera, Muscidae). J. Insects Food Feed 2023, 1, 1–14. [Google Scholar] [CrossRef]
- Couto, L.C.M.; Queiroz, M.M.C. Characterization of intrapuparial development of Musca domestica (Diptera: Muscidae), under different temperatures in laboratory conditions. J. Med. Entomol. 2024, 61, 64–73. [Google Scholar] [CrossRef]
- Greenberg, B. Flies and Disease: II. Biology and Disease Transmission; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar] [CrossRef]
- Moon, R.D. Muscid flies (muscidae). In Medical and Veterinary Entomology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–368. [Google Scholar] [CrossRef]
- Sasaki, T.; Kobayashi, M.; Agui, N. Epidemiological potential of excretion and regurgitation by Musca domestica (Diptera: Muscidae) in the dissemination of Escherichia coli O157: H7 to food. J. Med. Entomol. 2020, 37, 945–949. [Google Scholar] [CrossRef]
- Brito, L.G.; Oliveira, M.D.S.; Giglioti, R.; Barbieri, F.D.S.; Silva Neto, F.G.; Chagas, A.D.S.; Celestino, O.D.O. Manual de Identificação, Importância e Manutenção de Colônias Estoque de Dípteras de Interesse Veterinário em Laboratório; Documentos/Embrapa Rondonia; Embrapa Rondônia: Porto Velho, Brazil, 2008; 25p, ISSN 0103-9865. [Google Scholar]
- Nayduch, D.; Neupane, S.; Pickens, V.; Purvis, T.; Olds, C. House flies are underappreciated yet important reservoirs and vectors of microbial threats to animal and human health. Microorganisms 2023, 11, 583. [Google Scholar] [CrossRef]
- Carramaschi, I.N.; Castro, E.A.R.D.; Leite, J.A.; Queiroz, M.M.D.C.; Boas, M.H.S.V.; Rangel, K.; Zahner, V. First report of Raoultella ornithinolytica carrying blaKPC-2 isolated from a dipteran muscoid collected in a garbage from a public hospital in Rio de Janeiro, Brazil. Ver. Inst. Med. Trop. São Paulo 2019, 61, e32. [Google Scholar] [CrossRef]
- Carramaschi, I.N.; Lopes, J.C.O.; Leite, J.A.; Carneiro, M.T.; Barbosa, R.R.; Boas, M.H.V.; Zahner, V. Surveillance of antimicrobial resistant bacteria in flies (Diptera) in Rio de Janeiro city. Acta Trop. 2021, 220, 105962. [Google Scholar] [CrossRef]
- Abbas, M.N.; Sajeel, M.; Kausar, S. House fly (Musca domestica), a challenging pest; biology, management and control strategies. Elixir. Entomol. 2013, 64, 19333–19338. [Google Scholar]
- Young, B.W. The need for a greater understanding in the application of pesticides. Outlook Agric. 1996, 15, 80–87. [Google Scholar] [CrossRef]
- Pathak, V.M.; Verma, V.K.; Sharma, A.; Dewali, S.; Kumari, R.; Mohapatra, A.; Cunill, J.M. Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: A comprehensive review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef] [PubMed]
- Achari, T.S.; Barik, T.K.; Acharya, U.R. Toxins of Bacillus thuringiensis: A Novel Microbial Insecticide for Mosquito Vector Control. In Molecular Identification of Mosquito Vectors and Their Management; Springer: Berlin/Heidelberg, Germany, 2020; pp. 89–116. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef]
- Pinos, D.; Andrés-Garrido, A.; Ferré, J.; Hernández-Martínez, P. Response mechanisms of invertebrates to Bacillus thuringiensis and its pesticidal proteins. Microbiol. Mol. Biol. 2021, 85, 10-1128. [Google Scholar] [CrossRef]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial vegetative insecticidal proteins (Vip) from entomopathogenic bacteria. Microbiol. Mol. Biol. 2016, 80, 329–350. [Google Scholar] [CrossRef]
- Wang, C.; Li, M.; Chen, X.; Fan, S.; Lan, J. Binding Analysis of Sf-SR-C MAM Domain and Sf-FGFR Ectodomain to Vip3Aa. Insects 2024, 15, 428. [Google Scholar] [CrossRef]
- Pacheco, S.; Gómez, I.; Peláez-Aguilar, A.E.; Verduzco-Rosas, L.A.; García-Suárez, R.; do Nascimento, N.A.; Rivera-Nájera, L.Y.; Cantón, P.E.; Soberón, M.; Bravo, A. Structural changes upon membrane insertion of the insecticidal pore-forming toxins produced by Bacillus thuringiensis. Front. Insect Sci. 2023, 3, 1188891. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Mengist, H.M.; Shi, C.; Zhang, C.; Wang, B.; Li, T.; Huang, Y.; Xu, Y.; Jin, T. Structural basis of the pore-forming toxin/membrane interaction. Toxins 2021, 13, 128. [Google Scholar] [CrossRef]
- Chilcott, C.N.; Ellar, D.J. Comparative toxicity of Bacillus thuringiensis var. israelensis crystal proteins in vivo and in vitro. Microbiology 1988, 134, 2551–2558. [Google Scholar] [CrossRef] [PubMed]
- Guerchicoff, A.; Delécluse, A.; Rubinstein, C.P. The Bacillus thuringiensis cyt genes for hemolytic endotoxins constitute a gene family. Appl. Environ. Microbiol. 2001, 67, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, S.; Jin, M.; Wu, C.; Chakraborty, P.; Xiao, Y. Bacillus thuringiensis vegetative insecticidal protein family Vip3A and mode of action against pest Lepidoptera. Pest Manag. Sci. 2020, 76, 1612–1617. [Google Scholar] [CrossRef] [PubMed]
- Şahin, B.; Gomis-Cebolla, J.; Güneş, H.; Ferré, J. Characterization of Bacillus thuringiensis isolates by their insecticidal activity and their production of Cry and Vip3 proteins. PLoS ONE 2018, 13, e0206813. [Google Scholar] [CrossRef]
- Höfte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [CrossRef]
- Golding, N.; Wilson, A.L.; Moyes, C.L.; Cano, J.; Pigott, D.M.; Velayudhan, R.; Brooker, S.J.; Smith, D.L.; Hay, S.I.; Lindsay, S.W. Integrating vector control across diseases. BMC Med. 2015, 13, 249. [Google Scholar] [CrossRef]
- Rajagopal, G.; Ilango, S. Native Bacillus strains from infected insects: A potent bacterial agent for controlling mosquito vectors Aedes aegypti and Culex quinquefasciatus. Int. J. Mosq. Res. 2020, 7, 51–52. [Google Scholar]
- Valicente, F.H. Bacillus thuringiensis. In Natural Enemies of Insect Pests in Neotropical Agroecosystems; Souza, B., Vázquez, L., Marucci, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Queiroz, P.R.; Posso, M.C.; Martins, É.S.; Grynberg, P.; Togawa, R.; Monnerat, R.G. Identification of cry genes in Bacillus thuringiensis by multiplex real-time PCR. J. Microbiol. Methods 2023, 205, 106665. [Google Scholar] [CrossRef]
- Van Den Berg, H.; Takken, W. Viewpoint: A framework for decision-making in integrated vector management to prevent disease. Trop. Med. Int. Health 2007, 12, 1230–1238. [Google Scholar] [CrossRef]
- Li, K.; Chen, M.; Shi, J.; Mao, T. An overview of the production and use of Bacillus thuringiensis toxin. Open Life Sci. 2024, 19, 20220902. [Google Scholar] [CrossRef]
- Merdan, B.A. Bacillus thuringiensis as a feed additive to control Musca domestica associated with poultry houses. J. Basic Appl. Zool. 2012, 65, 83–87. [Google Scholar] [CrossRef]
- Nascimento, T.A.; Santos, M.J.P.; Valicente, F.H.; Dutok-Sánchez, C.M.; Queiroz, M.M.d.C. Desarrollo postembrionario de Musca domestica (Diptera: Muscidae) criada en diferentes dietas a base de harinas, en condiciones de laboratorio. Contrib. Cienc. Soc. 2024, 17, e8447. [Google Scholar] [CrossRef]
- Ferreira, D.F. Sisvar: A computer statistical analysis system. Ciência Agrotecnologia 2011, 35, 1039–1042. [Google Scholar] [CrossRef]
- Ceron, J.; Ortíz, A.; Quintero, R.; Güereca, L.; Bravo, A. Specific PCR primers directed to identify cryI and cryIII genes within a Bacillus thuringiensis strain collection. Appl. Environ. Microbiol. 1995, 61, 3826–3831. [Google Scholar] [CrossRef]
- Valicente, F.H.; de Toledo Picoli, E.A.; de Vasconcelos, M.J.V.; Carneiro, N.P.; Carneiro, A.A.; Guimarães, C.T.; Lana, U.G. Molecular characterization and distribution of Bacillus thuringiensis cry1 genes from Brazilian strains effective against the fall armyworm, Spodoptera frugiperda. Biol. Control 2010, 53, 360–366. [Google Scholar] [CrossRef]
- Ceron, J.; Covarrubias, L.; Quintero, R.; Ortiz, A.; Ortiz, M.; Aranda, E.; Bravo, A. PCR Analysis of the cryI insecticidal crystal family genes from Bacillus thuringiensis. Appl. Environ. Microbiol. 1994, 60, 353–356. [Google Scholar] [CrossRef]
- Fagundes, R.B.S. Laboratório de Controle Biológico, Embrapa Milho e Sorgo, Sete Lagoas, Minas Gerais, Brazil. 2019; unpublished work. [Google Scholar]
- Valicente, F.H. Laboratório de Controle Biológico, Embrapa Milho e Sorgo, Sete Lagoas, Minas Gerais, Brazil. 2010; unpublished work. [Google Scholar]
- Ibarra, J.E.; del Rincón, M.C.; Ordúz, S.; Noriega, D.; Benintende, G.; Monnerat, R.; Bravo, A. Diversity of Bacillus thuringiensis strains from Latin America with insecticidal activity against different mosquito species. Appl. Environ. Microbiol. 2003, 69, 5269–5274. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, C.S.; Boets, A.; Van Rie, J.; Ferré, J. Screening and identification of vip genes in Bacillus thuringiensis strains. Appl. Environ. Microbiol. 2009, 107, 219–225. [Google Scholar] [CrossRef]
- Valicente, F.H.; Souza, I.R.P. Cultivo e preparo de Bacillus thuringiensis para macroscopia eletrônica de varredura. In Congresso Nacional de Milho e Sorgo, 25, Proceedings of the Simpósio Brasileiro Sobre a Lagarta-do-Cartucho, Spodoptera frugiperda; Embrapa Milho e Sorgo; Brazilian Association of Corn and Sorghum (ABMS): Sete Lagoas, Brazil; Empaer: Cuiabá, Brazil, 2004. [Google Scholar]
- Lambert, B.; Peferoen, M. Insecticidal promise of Bacillus thuringiensis. BioScience 1992, 42, 112–122. [Google Scholar] [CrossRef]
- Habib, M.E.M.; Andrade, C.F.S. Bactérias entomopatogênicas. Controle Microbiano Insetos 1998, 2, 383–446. [Google Scholar]
- Valicente, F.H.; Barreto, M.R. Bacillus thuringiensis survey in Brazil: Geographical distribution and insecticidal activity against Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Neotrop. Entomol. 2003, 32, 639–644. [Google Scholar] [CrossRef]
- Djenane, Z.; Nateche, F.; Amziane, M.; Gomis-Cebolla, J.; El-Aichar, F.; Khorf, H.; Ferré, J. Assessment of the Antimicrobial Activity and the Entomocidal Potential of Bacillus thuringiensis Isolates from Algeria. Toxins 2017, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Travers, R.S.; Martin, P.A.; Reichelderfer, C.F. Selective process for efficient isolation of soil Bacillus spp. Appl. Environ. Microbiol. 1987, 53, 1263–1266. [Google Scholar] [CrossRef]
- Uribe, D.; Martinez, W.; Ceron, J. Distribution and diversity of cry genes in native strains of Bacillus thuringiensis obtained from different ecosystems from Colombia. J. Invert. Pathol. 2003, 82, 119–127. [Google Scholar] [CrossRef]
- Mukhija, B.; Khanna, V. Isolation, characterization and crystal morphology study of Bacillus thuringiensis isolates from soils of Punjab. J. Pure Appl. Microbiol. 2018, 12, 189–193. [Google Scholar] [CrossRef]
- da Costa Fernandes, G.; de Prado Costa, D.K.; de Oliveira, N.S.; de Sousa, E.C.P.; Machado, D.H.B.; Polanczyk, R.A.; de Siqueira, H.Á.A.; da Silva, M.C. Genetic diversity of Brazilian Bacillus thuringiensis isolates with toxicity against Aedes aegypti (Diptera: Culicidae). Sci. Rep. 2022, 12, 14408. [Google Scholar] [CrossRef]
- Seifinejad, A.; Jouzani, G.S.; Hosseinzadeh, A.; Abdmishani, C. Characterization of Lepidoptera-active cry and vip genes in Iranian Bacillus thuringiensis strain collection. Biol. Control 2008, 44, 216–226. [Google Scholar] [CrossRef]
- Espinasse, S.; Chaufaux, J.; Buisson, C.; Perchat, S.; Gohar, M.; Bourguet, D.; Sanchis, V. Occurrence and Linkage Between Secreted Insecticidal Toxins in Natural Isolates of Bacillus thuringiensis. Curr. Microbiol. 2003, 47, 501–507. [Google Scholar] [CrossRef]
- Nascimento, T.A.; de Carvalho, K.S.; Martins, D.C.; de Oliveira, C.R.; de Carvalho Queiroz, M.M.; Valicente, F.H. Bacillus thuringiensis strains to control Noctuidae pests. J. Appl. Entomol. 2024, 148, 371–381. [Google Scholar] [CrossRef]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and Its Dipteran-Specific. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Hemthanon, T.; Promdonkoy, B.; Boonserm, P. Screening and characterization of Bacillus thuringiensis isolates for high production of Vip3A and Cry proteins and high thermostability to control Spodoptera spp. J. Invertebr. Pathol. 2023, 201, 108020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fang, L.; Zhou, Z.; Pacheco, S.; Gomez, I.; Song, F.; Soberon, M.; Zhang, J.; Bravo, A. Specific binding between Bacillus thuringiensis Cry9Aa and Vip3Aa toxins synergizes their toxicity against Asiatic rice borer (Chilo suppressalis). J. Biol. Chem. 2018, 293, 11447–11458. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, J.; He, J.; Wu, J.; Sun, J.; Wang, R.; Zhang, W. Vip3Aa from Bacillus thuringiensis subsp. kurstaki HD1 is toxic to Aedes aegypti (Diptera: Culicidae). J. Invertebr. Pathol. 2020, 171, 107342. [Google Scholar] [CrossRef] [PubMed]
- Boukedi, H.; Khedher, S.B.; Abdelkefi-Mesrati, L.; Van Rie, J.; Tounsi, S. Comparative analysis of the susceptibility/tolerance of Spodoptera littoralis to Vip3Aa, Vip3Ae, Vip3Ad and Vip3Af toxins of Bacillus thuringiensis. J. Invertebr. Pathol. 2018, 152, 30–34. [Google Scholar] [CrossRef]
- Bernardi, O.; Bernardi, D.; Horikoshi, R.J.; Okuma, D.M.; Miraldo, L.L.; Fatoretto, J.; Medeiros, F.C.; Burd, T.; Omoto, C. Selection and characterization of resistance to the Vip3Aa20 protein from Bacillus thuringiensis in Spodoptera frugiperda. Pest Manag. Sci. 2016, 72, 1794–1802. [Google Scholar] [CrossRef]
- Chen, D.; Moar, W.J.; Jerga, A.; Gowda, A.; Milligan, J.S.; Bretsynder, E.C.; Rydel, T.J.; Baum, J.A.; Semeao, A.; Fu, X.; et al. Bacillus thuringiensis chimeric proteins Cry1A.2 and Cry1B.2 to control soybean lepidopteran pests: New domain combinations enhance insecticidal spectrum of activity and novel receptor contributions. PLoS ONE 2021, 16, e0249150. [Google Scholar] [CrossRef]
- Federici, B.A.; Park, H.-W.; Sakano, Y. Insecticidal protein crystals of Bacillus thuringiensis. In Inclusions in Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 195–236. [Google Scholar] [CrossRef]
- Johnson, C.; Bishop, A.H.; Turner, C.L. Isolation and Activity of Strains of Bacillus thuringiensis Toxic to Larvae of the Housefly (Diptera: Muscidae) and Tropical Blowflies (Diptera: Calliphoridae). J. Invertebr. Pathol. 1998, 71, 138–144. [Google Scholar] [CrossRef]
- Abdul-Rauf, M.; Ellar, D.J. Toxicity and Receptor Binding Properties of aBacillus thuringiensis CryIC Toxin Active against Both Lepidoptera and Diptera. J. Invertebr. Pathol. 1999, 73, 52–58. [Google Scholar] [CrossRef]
- Valtierra-de-Luis, D.; Villanueva, M.; Berry, C.; Caballero, P. Potential for Bacillus thuringiensis and other bacterial toxins as biological control agents to combat dipteran pests of medical and agronomic importance. Toxins 2020, 12, 773. [Google Scholar] [CrossRef]
- Mendoza, G.; Portillo, A. New combinations of cry genes from Bacillus thuringiensis strains isolated from northwestern Mexico. Int. Microbiol. 2012, 15, 209–216. [Google Scholar] [CrossRef]
- Van Frankenhuyzen, K. Insecticidal activity of Bacillus thuringiensis crystal proteins. J. Invertebr. Pathol. 2009, 101, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, F.A.; Crickmore, N. N-terminal proteolysis determines the differential activity of Bacillus thuringiensis Cry2A toxins towards Aedes aegypti. J. Invertebr. Pathol. 2024, 204, 108100. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.-Z.; Xu, L.; Liu, B.; Chen, Q.-X.; Zhu, Y.-J. Key residues of Bacillus thuringiensis Cry2Ab for oligomerization and pore-formation activity. AMB Express 2011, 11, 112. [Google Scholar] [CrossRef]
- Polanczyk, R.A.; van Frankenhuyzen, K.; Pauli, G. The American Bacillus thuringiensis based biopesticides market. In Bacillus thuringiensis and Lysinibacillus sphaericus: Characterization and Use in the Field of Biocontrole; Fiuza, L., Polanczyk, R., Crickmore, N., Eds.; Springer: Cham, Switzerland, 2017; pp. 173–184. [Google Scholar] [CrossRef]
- Raymond, B.; Johnston, P.R.; Nielsen-LeRoux, C.; Lereclus, D.; Crickmore, N. Bacillus thuringiensis: An impotent pathogen? Trends Microbiol. 2010, 18, 189–194. [Google Scholar] [CrossRef]
- Costa, M.L.; Lana, U.G.; Barros, E.C.; Paiva, L.V.; Valicente, F.H. Molecular characterization of Bacillus thuringiensis cyt genes efficient against fall armyworm, Spodoptera frugiperda. J. Agric. Sci. 2014, 6, 1–10. [Google Scholar] [CrossRef][Green Version]
- Vilas-Bôas, G.T.; Lemos, M.V.F. Diversity of cry genes and genetic characterization of Bacillus thuringiensis isolated from Brazil. Can. J. Microbiol. 2004, 50, 605–613. [Google Scholar] [CrossRef]
- Porcar, M.; Juárez-Pérez, V. PCR-based identification of Bacillus thuringiensis pesticidal crystal genes. FEMS Microbiol. Rev. 2003, 26, 419–432. [Google Scholar] [CrossRef]
- Agaisse, H.; Lereclus, D. How does Bacillus thuringiensis produce so much insecticidal crystal protein? J. Bacteriol. 1995, 177, 6027–6032. [Google Scholar] [CrossRef]
- Nair, K.; Al-Thani, R.; Ahmed, T.; Jaoua, S. Diversity of Bacillus thuringiensis strains from Qatar as shown by crystal morphology, δ-endotoxins and cry gene content. Front. Microbiol. 2018, 9, 339313. [Google Scholar] [CrossRef]
- Djenane, Z.; Lázaro-Berenguer, M.; Nateche, F.; Ferré, J. Evaluation of the toxicity of supernatant cultures and spore–crystal mixtures of Bacillus thuringiensis strains isolated from Algeria. Curr. Microbiol. 2020, 77, 2904–2914. [Google Scholar] [CrossRef]
- Yamamoto, T.; McLaughlin, R.E. Isolation of a protein from the parasporal crystal of Bacillus thuringiensis var kurstaki toxic to the mosquito larva, Aedes taeniorhynchus. Biochem. Biophys. Res. Commun. 1981, 103, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Sarabia, S.; Lopez, L.; Ontiveros, H.; Abarca, C.; Ortiz, A.; Ortiz, M.; Lina, L.; Villalobos, F.J.; Peña, G.; et al. Characterization of cry genes in a Mexican Bacillus thuringiensis strain collection. Appl. Environ. Microbiol. 1998, 64, 4965–4972. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef]
- Loutfi, H.; Fayad, N.; Pellen, F.; Le Jeune, B.; Chakroun, M.; Benfarhat, D.; Lteif, R.; Kallassy, M.; Le Brun, G.; Abboud, M. Morphological study of Bacillus thuringiensis crystals and spores. Appl. Sci. 2020, 11, 155. [Google Scholar] [CrossRef]
- Gitahy, P.D.M.; Souza, M.T.D.; Monnerat, R.G.; Arrigoni, E.D.B.; Baldani, J.I. A Brazilian Bacillus thuringiensis strain highly active to sugarcane borer Diatraea saccharalis (Lepidoptera: Crambidae). Braz. J. Microbiol. 2007, 38, 531–537. [Google Scholar] [CrossRef]
- Arsov, A.; Gerginova, M.; Paunova-Krasteva, T.; Petrov, K.; Petrova, P. Multiple cry genes in Bacillus thuringiensis strain BTG suggest a broad-spectrum insecticidal activity. Int. J. Mol. Sci. 2023, 24, 11137. [Google Scholar] [CrossRef]
- Scott, J.G.; Warren, W.C.; Beukeboom, L.W.; Bopp, D.; Clark, A.G.; Giers, S.D.; Hediger, M.; Jones, A.K.; Kasai, S.; Leichter, C.A.; et al. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol. 2014, 15, 466. [Google Scholar] [CrossRef]
- Geden, C.J.; Nayduch, D.; Scott, J.G.; Burgess, E.R., IV; Gerry, A.C.; Kaufman, P.E.; Machtinger, E.T. House fly (Diptera: Muscidae): Biology, pest status, current management prospects, and research needs. J. Integr. Pest Manag. 2021, 12, 39. [Google Scholar] [CrossRef]
- Soares-da-Silva, J.; Pinheiro, V.C.S.; Litaiff-Abreu, E.; Polanczyk, R.A.; Tadei, W.P. Isolation of Bacillus thuringiensis from the state of Amazonas, in Brazil, and screening against Aedes aegypti (Diptera, Culicidae). Rev. Bras. Entomol. 2015, 59, 1–6. [Google Scholar] [CrossRef]
- Šebesta, K.; Horska, K.; Vaňková, J. Inhibition of de novo RNA synthesis by the insecticidal exotoxin of Bacillus thuringiensis var. gelechiae. Collect. Czechoslov. Chem. Commun. 1969, 34, 1786–1791. [Google Scholar] [CrossRef]
- Gowelo, S.; Chirombo, J.; Spitzen, J.; Koenraadt, C.J.; Mzilahowa, T.; Van Den Berg, H.; Takken, W.; McCann, R. Effects of larval exposure to sublethal doses of Bacillus thuringiensis var. israelensis on body size, oviposition and survival of adult Anopheles coluzzii mosquitoes. Parasites Vectors 2020, 13, 259. [Google Scholar] [CrossRef]
- Iqbal, H.; Fatima, A.; Khan, H.A.A. ZnO nanoparticles produced in the culture supernatant of Bacillus thuringiensis ser. israelensis affect the demographic parameters of Musca domestica using the age-stage, two-sex life table. Pest Manag. 2022, 78, 1640–1648. [Google Scholar] [CrossRef] [PubMed]
- Hanski, I. Carrion fly community dynamics: Patchiness, seasonality and coexistence. Ecol. Entomol. 1987, 12, 257–266. [Google Scholar] [CrossRef]
- Davidowitz, G.; D’Amico, L.J.; Nijhout, H.F. Critical weight in the development of insect body size. Evol. Dev. 2003, 5, 188–197. [Google Scholar] [CrossRef]
- Chapman, R.F.; Simpson, S.J.; Douglas, A.E. The Insects: Structure and Function, 5th ed.; Cambridge University Press: New York, NY, USA, 2013; ISBN 978-0-521-11389-2. [Google Scholar]
- Ullyett, G. Pupation habits of sheep blowflies in relation to parasitism by Mormoniella vitripennis, Wlk.(Hym., Pteromalid.). Bull. Entomol. Res. 1950, 40, 533–537. [Google Scholar] [CrossRef]
- Goodbrod, J.R.; Goff, M.L. Effects of larval population density on rates of development and interactions between two species of Chrysomya (Diptera: Calliphoridae) in laboratory culture. J. Med. Entomol. 1990, 27, 338–343. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Parra, J.R.P.P. Insect Bioecology and nutrition for integrated pest management. Fla. Entomol. 2013, 96, 298–299. [Google Scholar]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Large-scale production of house fly, Musca domestica (Diptera: Muscidae), larvae fed 3 manure types. J. Econ. Entomol. 2023, 116, 1102–1109. [Google Scholar] [CrossRef]
- Fisher, R. The evolution of dominance in certain polymorphic species. Am. Nat. 1930, 64, 385–406. [Google Scholar] [CrossRef]
- Norris, K. The bionomics of blow flies. Annu. Rev. Entomol. 1965, 10, 47–68. [Google Scholar] [CrossRef]
- Oliveira, E.M.D.; Sales, V.H.G.; Andrade, M.S.; Zilli, J.É.; Borges, W.L.; Souza, T.M.D. Isolation and Characterization of Biosurfactant-Producing Bacteria from Amapaense Amazon Soils. Intern. J. Microbiol. 2021, 2021, 9959550. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).