
Thought to Be Extinct, but Still Alive Today: The Miocene Genus Primascena Klimaszewsi, 1997 (Hemiptera: Psyllidae) in the Light of Two Extant Species from Brazil

Abstract
Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Psyllid Genus | Number of Species | Host Plants | Distribution |
|---|---|---|---|
| Aphalaroida Crawford [21] | 9 | Fabaceae (8) | North America |
| Baccharopelma Burckhardt, et al. [22] | 6 | Asteraceae (5) | South America |
| Connectopelma Šulc [23] | 6 | Rhamnaceae (6) | South America |
| Ehrendorferiana Burckhardt [9] | 2 | Cupressaceae (2) | South America |
| Freysuila Aleman [24] | 3 | Fabaceae (2), Santalaceae (1) | North and South America |
| Pachyparia Loginova [25] | 1 | Fabaceae (1) | Africa |
| Panisopelma Enderlein [26] | 12 | Zygophyllaceae (10) | South America |
| Primascena Klimaszewski [13] | 3 (including 1 fossil) | Polygonaceae (2) | South America |
| Prosopidopsylla Burckhardt [16] | 5 | Fabaceae (5) | South America |
| Russelliana Tuthill [27] | 43 | Asteraceae (4), Fabaceae (12), Polygonaceae (1), Rosaceae (2), Solanaceae (16), Verbenaceae (5), polyphagous (1) | South America |
| Sphinia Blanchard [28] | 6 | Euphorbiaceae (4), Rosaceae (2) | South America |
| Yangus Fang [29] | 3 | Fabaceae (3) | Asia |
| Zonopelma Burckhardt [16] | 2 | Misodendraceae (2) | South America |
2. Materials and Methods
2.1. Material Examined
2.2. Taxon and Character Selection
2.3. Phylogenetic Analysis
- List of Characters
- Adult
- 0.
- Glandular hairs on body integument: absent (0); present (1).
- 1.
- Vertex: subrectangular (0); trapezoidal (1).
- 2.
- Vertex: flat (0); with flat anterior lobes (1); weakly bulged anteriorly (2); with large anterior tubercles (3).
- 3.
- Genal processes: absent (0); present, shorter than vertex (1); present, longer than vertex, (2).
- 4.
- Preocular sclerite: absent (0); present as small sclerite, narrower than one fifth of the eye width (1); present as large sclerite, wider than one quarter of the eye width (2).
- 5.
- Eyes: hemispherical, adpressed to head (0); weakly stalked, posterior margin of head not indented between eye and lateral ocellus (1); strongly stalked, posterior margin of head indented between eye and lateral ocellus (2).
- 6.
- Sensory plates on antennal segment III: absent (0); present (1).
- 7.
- Antennal rhinaria: without a wreath of spines (0); with a partial wreath of spines on all segments (1); with a complete wreath of spines on all segments (2); on some segments with complete or partial wreath of spines (3).
- 8.
- Rostrum: short, stout or moderately long but only tip of apical segment visible in lateral view (0); long, slender, entire apical segment and part of subapical segment visible in lateral view (1).
- 9.
- Pronotum: flat (0); with two tubercules on either side (1); with a pit on either side (2); with two pits on either side (3).
- 10.
- Propleurites: broader than high, epimeron and episternum subequal (0); higher than broad, epimeron and episternum subequal (1); narrow, episternum much smaller than epimeron (2).
- 11.
- Tubercle on metapostnotum: flat (0); large, strongly produced (1).
- 12.
- Genual spine: absent (0); present (1).
- 13.
- Metatibial and metabasitarsal spurs: strongly sclerotized (0); hardly sclerotized (1).
- 14.
- Metatibia bearing: a posteriorly open crown of < 8 evenly spaced apical spurs (0); a posteriorly open crown of >7 densely spaced apical spurs (1); distinctly grouped apical spurs (2).
- 15.
- Metabasitarsus: with 2 lateral spurs (0); with 1 lateral spur (1); with 1 microscopical lateral spur (2); without lateral spurs (3).
- 16.
- Length of forewing vein R relative to that of M+Cu: shorter (0); as long as (1); longer (2).
- 17.
- Forewing: Rs and M separate (0); Rs and M touching in a point or partially fused (1); with r-m crossvein (2).
- 18.
- Forewing vein M: longer than its branches (0); shorter than its branches (1).
- 19.
- Forewing cell cu1: short, about 1.5 times as high as long (0); longer than high (1); about twice as high as long (2).
- 20.
- Hindwing costal setae: not grouped (0); grouped (1).
- 21.
- Hindwing vein R+M+Cu: trifurcating into R, M and Cu (0); bifurcating into R and M+Cu (1); bifurcating into R+M and Cu (2).
- Terminalia
- 22.
- Male subgenital plate: subglobular (0); distinctly elongate (1).
- 23.
- Paramere: lamellar with antero-apical or apical sclerotized tooth (0); slender, digitiform (1); lamellar, with forward-directed tooth, and inner tooth at apical margin (2); base robust, apex slender, backwards directed (3); complex, sickle-shaped (4); short, with forward-directed hook (5); short, rectangular, sometimes with antero-apical process (6); lamellar/spiniform with inner sclerotized process/tooth in basal half (7); oval or rectangular with inner process (8).
- 24.
- Distal segment of aedeagus: without lateral lobes (0); with lateral lobes (1).
- 25.
- Distal segment of aedeagus: with small, oval apical inflation (0); with large, irregularly triangular apical inflation (1); with large apical inflation divided into ventral and dorsal parts (2); with moderately sized to large apical inflation incised dorso-basally (3); tubular with appendages (4).
- 26.
- Sclerotized end tube of ductus ejaculatorius: small, shorter than 5 times its diameter (0); medium sized, 5–10 times as long as its diameter and less than a fifth of the length of the distal segment of aedeagus (1); large, a quarter of the length of the distal segment of aedeagus (2); very large, longer than a third of the length of the distal segment of aedeagus (3).
- 27.
- Female proctiger with dorsal margin: straight or curved (0); with a slight depression distal to circumanal ring (1); strongly bent down distal to circumanal ring (2).
- 28.
- Female subgenial plate: long, in lateral view longer than high, lacking glandular setae (0); short, in lateral view as long as high, lacking glandular setae (1); short, in lateral view as long as high, with glandular setae (2).
- 29.
- Brush-like combs of fringing hairs on distal abdominal sternites in female: absent (0); present (1).
- Fifth instar immature
- 30.
- Antenna: 7-segmented (0); 5-segmented (1); 3-segmented (2); 8-segmented (3).
- 31.
- Humeral lobes: absent (0); present (1).
- 32.
- Thoracic tergites: small, covering less than half of the surface of meso and metanotum (0); medium-sized, covering more than half of the surface of meso and metanotum but leaving membranous areas (1); large, almost completely covering meso and metanotum (2).
- 33.
- Body margin and dorsum of immature: with long capitate setae (0); with short or no marginal setae (1); with diamond-shaped setae on margin of wing pads (2); with a few marginal capitate setae on abdomen, otherwise without setae (3); with bristle-like setae (4); with marginal rod setae on caudal plate, without dorsal setae (5); with capitate and short funnel-shaped setae (6); with simple setae (7); with club-shaped setae (8).
- 34.
- Immature forewing pads: with no or few long dorsal or marginal simple setae (0); with moderately long simple setae (1); with long capitate setae (2); with marginal long rod setae (3); with long marginal and funnel-shaped dorsal setae (4); with diamond or club-shaped setae (5)
- 35.
- Pedicel of tarsal arolium: as long as claws (0); longer than claws (1); shorter than claws (2).
- 36.
- Anus: ventral, near hind margin, distance from anus to hind margin of caudal plate less than maximum length of circumanal ring (0); ventral, distant from hind margin, distance from anus to hind margin of caudal plate more than maximum length of circumanal ring (1); terminal (2).
- 37.
- Circumanal ring: unicellular (0); multicellular (1).
- 38.
- Circumanal ring: small, narrower than a third width of terminal ventrite (0); large, wider than three quarters width of terminal ventrite (1).
- 39.
- Pores of the circumanal ring: narrow, elongate (0); circular (1).
3. Results
3.1. Phylogeny
3.2. Taxonomy
3.2.1. Subfamily: Aphalaroidinae Vondráček, 1963: 277 [18]. Type: genus. Aphalaroida Crawford
3.2.2. Genus: Primascena Klimaszewsi, 1997 [13]
3.2.3. Key to Species of Adult Primascena
- 1.
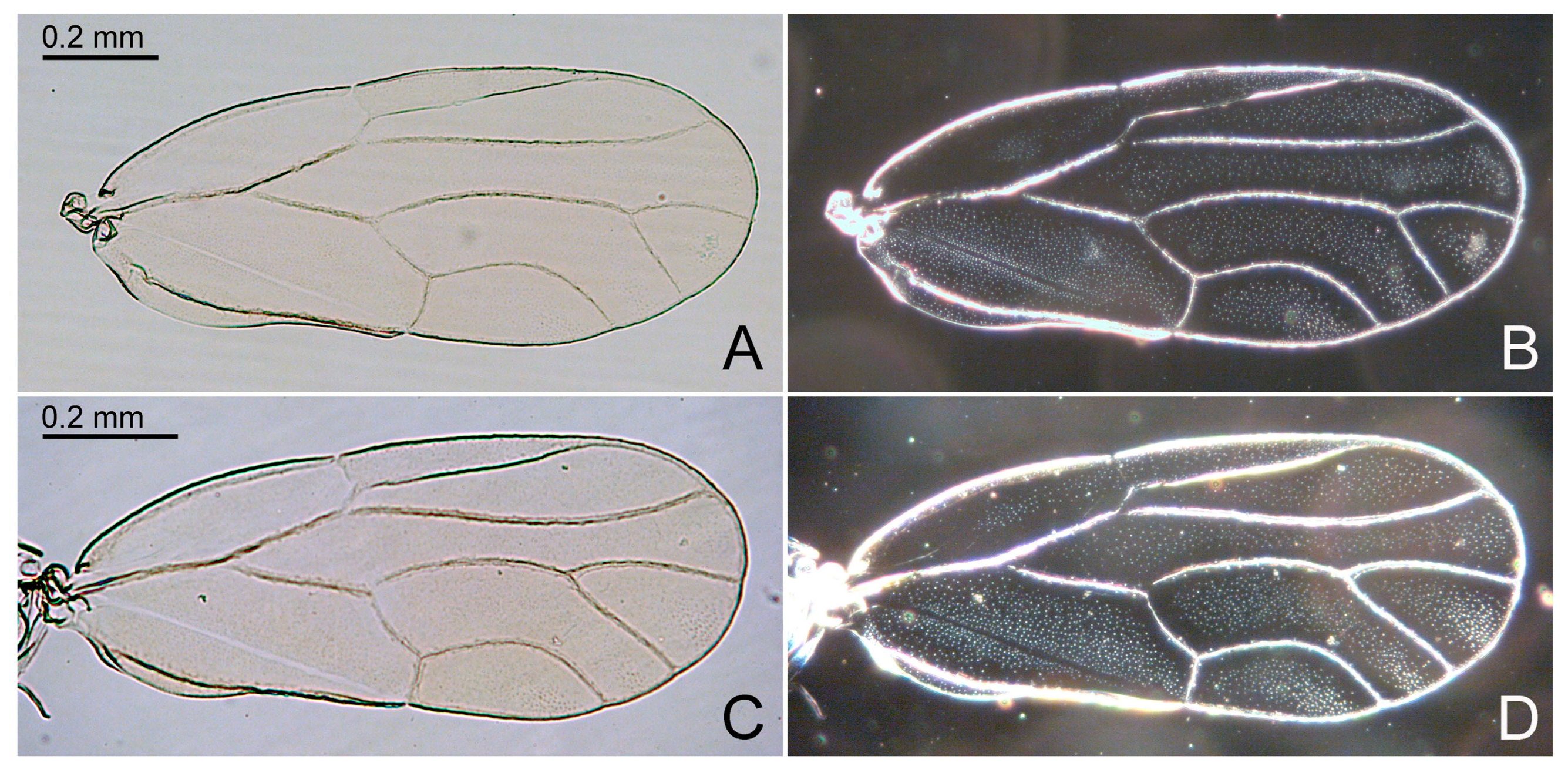
- Antennal segment III more than twice as long as segment IV (Figure 4A). Metabasitarsus with two spurs. Pterostigma of forewing (Figure 4D) narrower than adjacent cell r1, at base. Paramere, in lateral view, rectangular (Figure 4E). Apical inflation of distal aedeagal segment moderately sized (Figure 4E) Primascena subita.
- -
- 2.
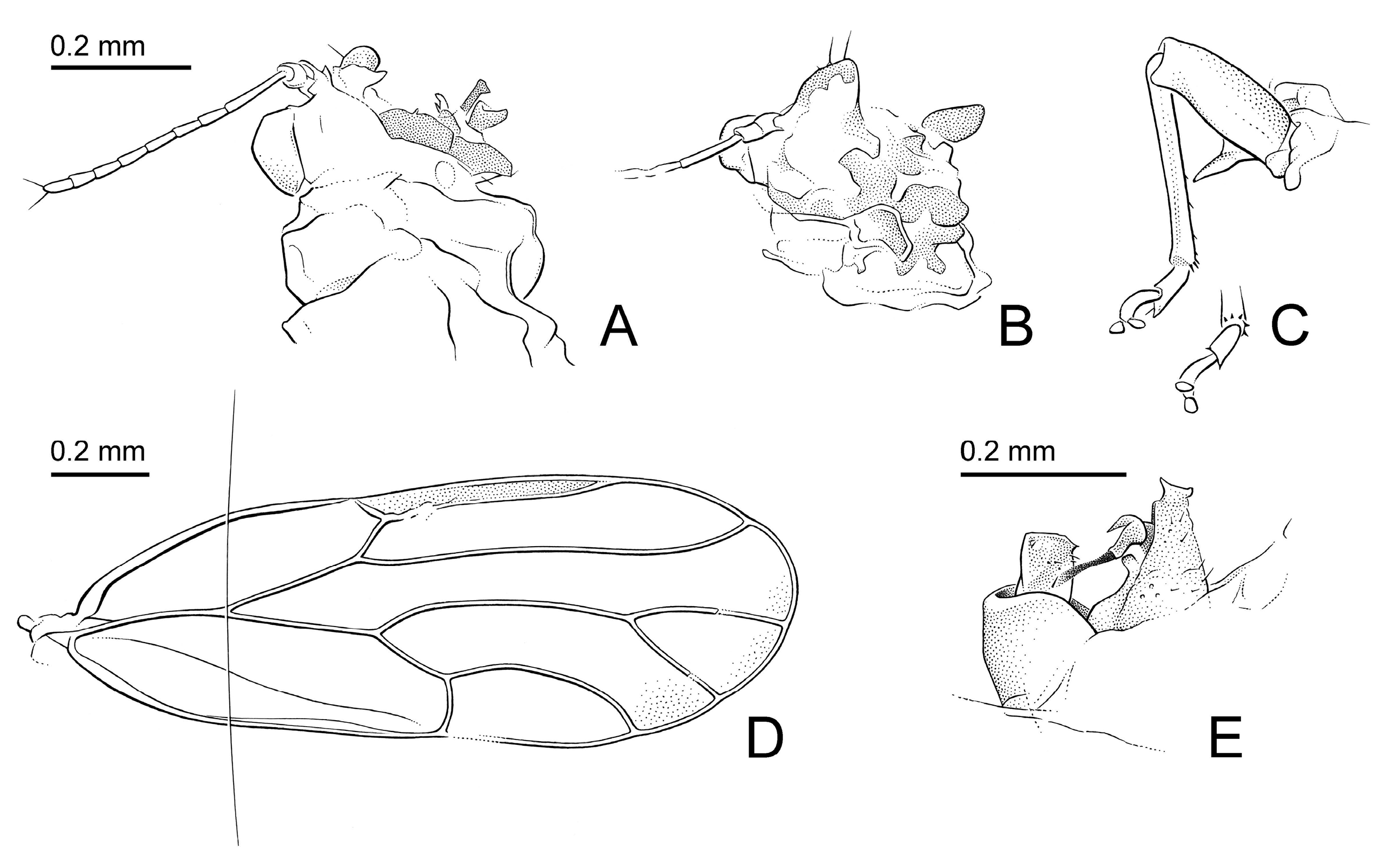
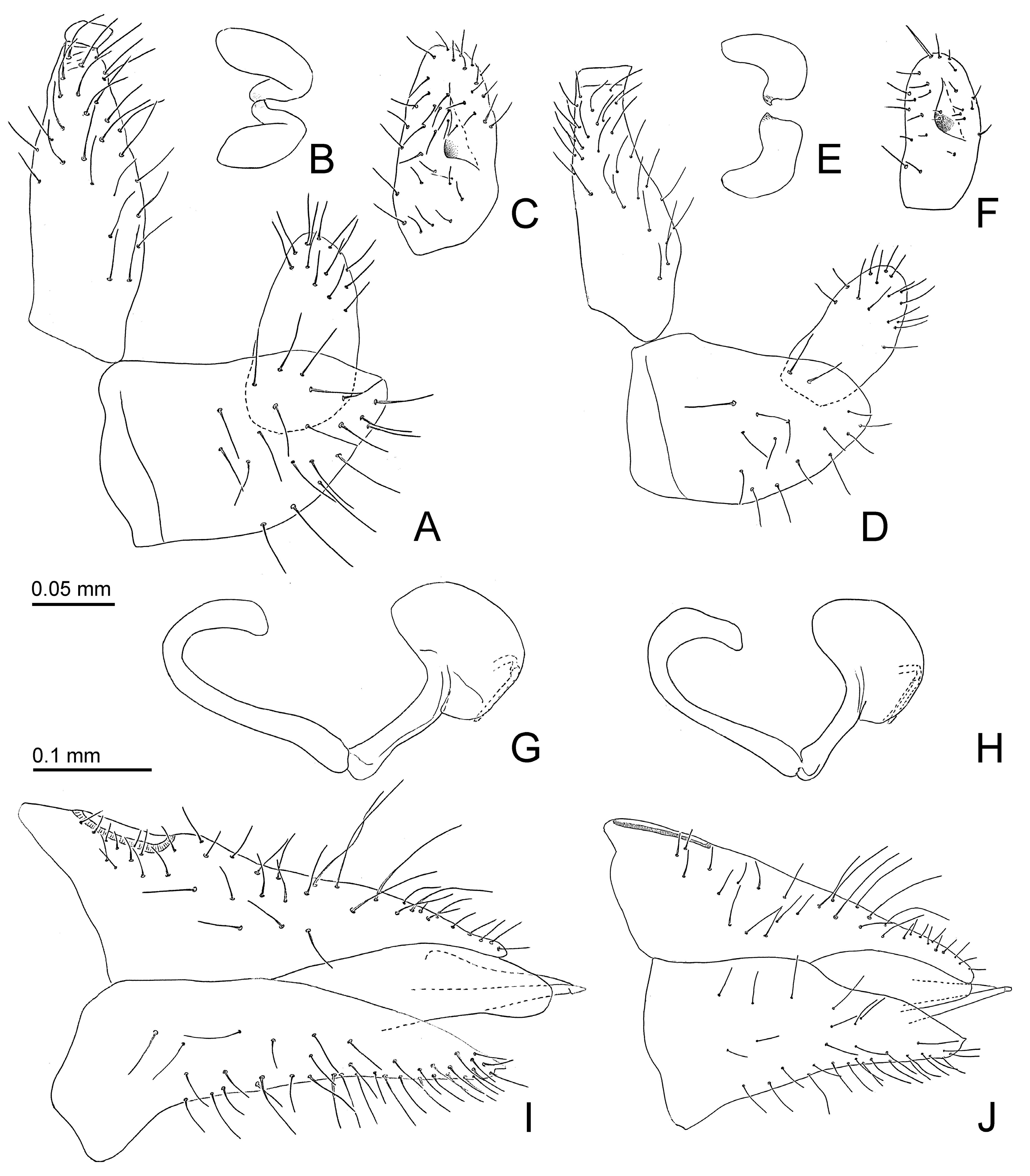
- Genal processes dark brown or black (Figure 2C). Forewing 3.0 or more times as long as head width; m1 cell value ≤ 1.4. Paramere, in lateral view, slightly angular apically (Figure 5A); inner face with relatively large process proximal of the middle (Figure 5B,C). Apical inflation of distal aedeagal segment relatively angular (Figure 5G). Dorsal margin of female proctiger, in lateral view, weakly sinuate (Figure 5I). Probable host plant Ruprechtia laxiflora Primascena empsycha.
- -
- Genal processes light (Figure 2C). Forewing shorter than 3.0 times head width; m1 cell value ≥ 1.5. Paramere, in lateral view, rounded apically (Figure 5D); inner face with relatively small process distal of the middle (Figure 5E,F). Apical inflation of distal aedeagal segment relatively rounded (Figure 5H). Dorsal margin of female proctiger, in lateral view, almost straight (Figure 5J). Host plant Ruprechtia exploratricis Primascena ruprechtiae.
3.2.4. Primascena empsycha New Species (Figure 2A–C, Figure 3A,B and Figure 5A–C,G,I)
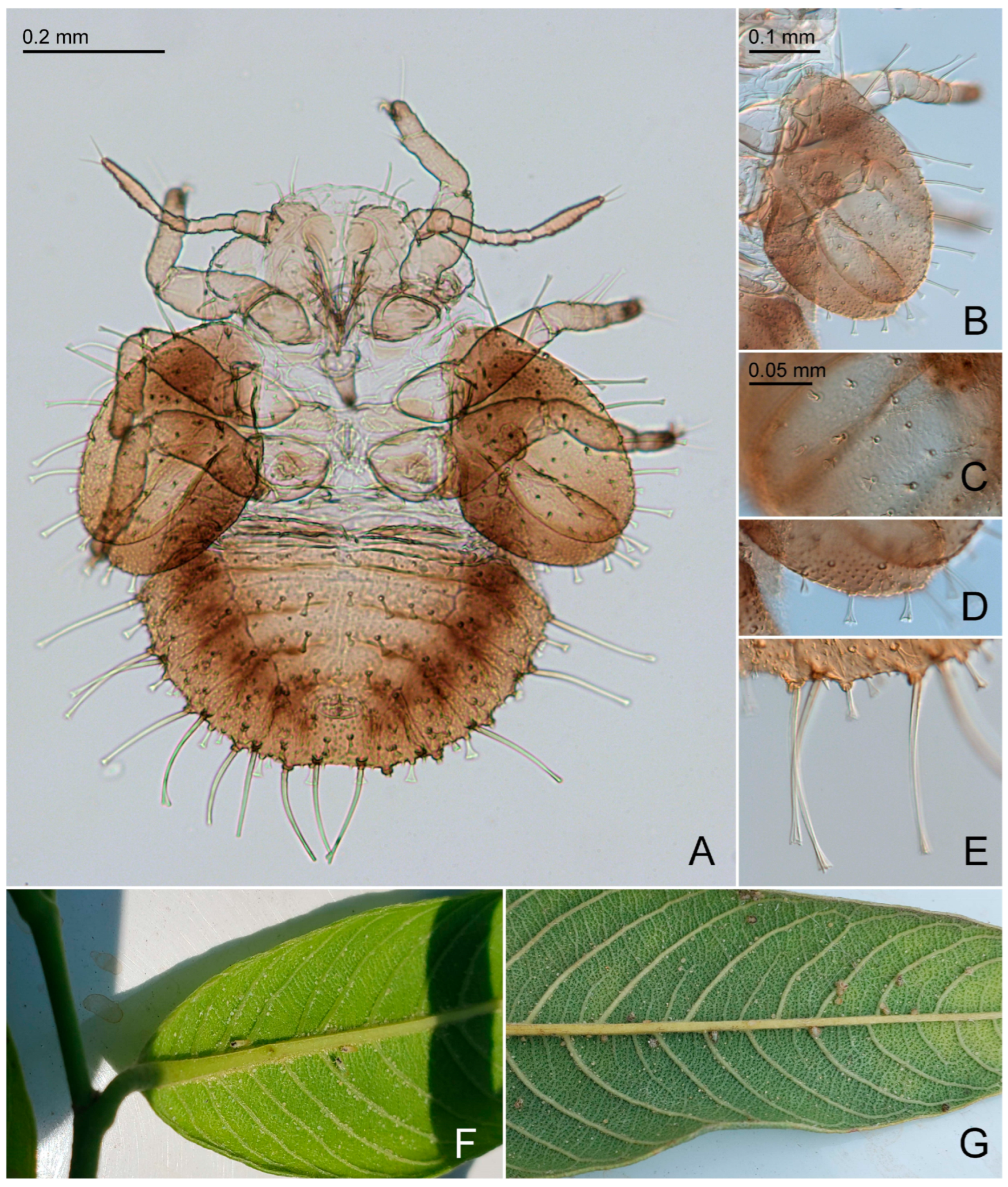
3.2.5. Primascena ruprechtiae New Species (Figure 2D–I, Figure 3C,D, Figure 5D–F,H,J and Figure 6)
3.2.6. Primascena subita Klimaszewsi, 1998 [13] (Figure 4)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouvrard, D.; Burckhardt, D.; Greenwalt, D. The Oldest Jumping Plant-Louse (Insecta: Hemiptera: Sternorrhyncha) with Comments on the Classification and Nomenclature of the Palaeogene Psylloidea. Acta Musei Morav. Sci. Biol. Brno 2013, 98, 21–33. [Google Scholar]
- Burckhardt, D.; Serbina, L.Š.; Malenovský, I.; Queiroz, D.L.; Aléné, D.C.; Cho, G.; Percy, D.M. Phylogeny and Classification of Jumping Plant Lice of the Subfamily Liviinae (Hemiptera: Psylloidea: Liviidae) Based on Molecular and Morphological Data. Zool. J. Linn. Soc. 2023, 1–35. [Google Scholar] [CrossRef]
- Burckhardt, D.; Queiroz, D.L. Neotropical Jumping Plant-Lice (Hemiptera, Psylloidea) Associated with Plants of the Tribe Detarieae (Leguminosae, Detarioideae). Zootaxa 2020, 4733, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Burckhardt, D.; Ouvrard, D.; Percy, D.M. An Updated Classification of the Jumping Plant-Lice (Hemiptera: Psylloidea) Based on Molecular and Morphological Evidence. Eur. J. Taxon. 2021, 736, 137–182. [Google Scholar] [CrossRef]
- Ouvrard, D. Psyl’list—The World Psylloidea Database. Available online: http://www.hemiptera-databases.com/psyllist (accessed on 3 February 2024).
- Burckhardt, D.; Ouvrard, D.; Queiroz, D.L.; Percy, D.M. Psyllid Host-Plants (Hemiptera: Psylloidea): Resolving a Semantic Problem. Fla. Entomol. 2014, 97, 242–246. [Google Scholar] [CrossRef]
- Ouvrard, D.; Burckhardt, D.; Cocquempot, C. An Annotated Checklist of the Jumping Plant-Lice (Insecta: Hemiptera: Psylloidea) from the Mercantour National Park, with Seven New Records for France and One New Synonymy. Zoosystema 2015, 37, 251–271. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Bird, J. Sedge and Rush-Feeding Psyllids of the Subfamily Liviinae (Insecta: Hemiptera: Psylloidea): A Review. Zool. J. Linn. Soc. 2000, 128, 1–49. [Google Scholar] [CrossRef]
- Burckhardt, D. Ehrendorferiana, a New Genus of Neotropical Jumping Plant Lice (Insecta: Hemiptera: Psylloidea) Associated with Conifers (Cupressaceae). Org. Divers. Evol. 2005, 5, 317–319. [Google Scholar] [CrossRef]
- Bekker Migdisova, E.E. Iskopajemije Nasekomije Psillomorfy; Izdatelstvo “Natshka” Ed.; Trudy paleontologitshekogo Instituta: Moscow, Russia, 1985; Volume 206, p. 92. [Google Scholar]
- Iturralde-Vinent, M.A.; Macphee, R.D.E. Remarks on the Age of Dominican Amber. Palaeoentomology 2019, 2, 236–240. [Google Scholar] [CrossRef]
- Klimaszewski, S.M. New Psyllids (Homoptera, Psylloidea) from Dominican Amber. Acta Biol. Silesiana 1996, 29, 24–44. [Google Scholar]
- Klimaszewski, S.M. Further Data Concerning Dominican amber Jumping Plant Lice (Homoptera, Psylloidea). Acta Biol. Silesiana 1997, 30, 19–27. [Google Scholar]
- Burckhardt, D.; Drohojowska, J.; Serbina, L.Š.; Malenovský, I. First Record of Jumping Plant Lice of the Family Liviidae (Insecta, Hemiptera, Psylloidea) from Dominican Amber. Neues Jahrb. Für Geol. Und Palaontol. 2024. accepted. [Google Scholar]
- Burckhardt, D.; Mifsud, D. Jumping Plant-Lice of the Paurocephalinae (Insecta, Hemiptera, Psylloidea): Systematics and Phylogeny. Contrib. Nat. Hist. 2003, 2, 3–34. [Google Scholar]
- Burckhardt, D. Jumping Plant Lice (Homoptera: Psylloidea) of the Temperate Neotropical Region: Part 1. Psyllidae (subfamilies Aphalarinae, Rhinocolinae and Aphalaroidinae). Zool. J. Linn. Soc. 1987, 89, 299–392. [Google Scholar] [CrossRef]
- Percy, D.M.; Crampton-Platt, A.; Sveinsson, S.; Lemmon, A.R.; Lemmon, E.M.; Ouvrard, D.; Burckhardt, D. Resolving the Psyllid Tree of Life: Phylogenomic Analyses of the Superfamily Psylloidea (Hemiptera). Syst. Entomol. 2018, 43, 762–776. [Google Scholar] [CrossRef]
- Vondráček, K. Jumping Plant-Lice (Psylloidea-Homoptera) of Central Africa. Part 1 (Congo). Acta Entomol. Musei Natl. Pragae 1963, 35, 263–290. [Google Scholar]
- Burckhardt, D.; Ouvrard, D. A Revised Classification of the Jumping Plant-Lice (Hemiptera: Psylloidea). Zootaxa 2012, 3509, 1–34. [Google Scholar] [CrossRef]
- Ouvrard, D.; Chalise, P.; Percy, D.M. Host-Plant Leaps Versus Host-Plant Shuffle: A Global Survey Reveals Contrasting Patterns in an Oligophagous Insect Group (Hemiptera, Psylloidea). Syst. Biodivers. 2015, 13, 434–454. [Google Scholar] [CrossRef]
- Crawford, D.L. A Monograph of the Jumping Plant-Lice or Psyllidae of the New World; Smithsonian Institution United States National Museum, Ed.; Government Printing Office: Washington, DC, USA, 1914; Volume 85. [Google Scholar]
- Burckhardt, D.; Espirito-Santo, M.M.; Fernandes, G.W.; Malenovsky, I. Gall-Inducing Jumping Plant-Lice of the Neotropical genus Baccharopelma (Hemiptera, Psylloidea) Associated with Baccharis (Asteraceae). J. Nat. Hist. 2004, 38, 2051–2071. [Google Scholar] [CrossRef]
- Šulc, K. Eine neue Panisopelma (Connectopelma sbgn. n.) und eine neue Trioza aus Chile. Sitzungsber. Königl.-Böhm. Ges. Wissensch. Math.-Naturw. Classe 1914, 5, 1–8. [Google Scholar]
- Aleman, D.J. Frey-Suila Dugesii. La Nat. 1887, 1, 21–26. [Google Scholar]
- Loginova, M.M. Ergebnisse der Zoologischen Nubien-Expedition 1962. Teil XXXIII. Homoptera-Psylloidea. Ann. Naturhist. Mus. Wien 1967, 70, 401–409. [Google Scholar]
- Enderlein, G. Panisopelma quadrigibbiceps, eine neue Psyllidengattung aus Argentinien. (Psyllidologica I). Zool. Anz. 1910, 36, 280–281. [Google Scholar]
- Tuthill, L.D. Los Psyllidae del Peru Central (Insecta, Homoptera). Rev. Peru. De Entomol. Agric. 1959, 2, 1–27. [Google Scholar]
- Blanchard, E.; Zoologia, V. Afidideos. In Historia Fisica y Politica de Chile; Museo de Historia Natural de Santiago: Santiago, Chile, 1852; Volume 7, pp. 306–320. [Google Scholar]
- Fang, S.J. Psylloidea of Taiwan: Supplement II. (Homoptera). J. Taiwan Mus. 1990, 43, 103–117. [Google Scholar]
- Bastin, S.; Burckhardt, D.; Reyes-Betancort, J.A.; Hernandez-Suarez, E.; Ouvrard, D. A Review of the Jumping Plant-Lice (Hemiptera: Psylloidea) of the Canary Islands, with Descriptions of Two New Genera and Sixteen New Species. Zootaxa 2023, 5313, 1–98. [Google Scholar] [CrossRef]
- Ouvrard, D.; Bourgoin, T.; Campbell, B.C. Comparative Morphological Assessment of the Psyllid Pleuron (Insecta, Hemiptera, Sternorrhyncha). J. Morphol. 2002, 252, 276–290. [Google Scholar] [CrossRef]
- Drohojowska, J. Thorax Morphology and Its Iiportance in Establishing Relationships within Psylloidea (Hemiptera, Sternorrhyncha); Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2015; Volume 3414, p. 172. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 23 March 2024).
- Zachos, F.E. Species Concepts in Biology, Historical Development, Theoretical Foundations and Practical Relevance; Springer: Berlin/Heidelberg, Germany, 2016; p. 220. [Google Scholar] [CrossRef]
- Nelson, G.; Platnick, N.I. Systematics and Biogeography: Cladistics and Vicariance; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Burckhardt, D. Insects of Saudi Arabia. Sternorrhyncha: Suborder Psyllodea. Fauna Saudi Arab. 1981, 3, 213–226. [Google Scholar]
- Nixon, K.C. Winclada; v.1.00.08; K.C. Nixon: Ithaca, NY, USA, 2002. [Google Scholar]
- Goloboff, P.A.; Catalano, S.A. Tnt V.1.5, Including a Full Implementation of Phylogenetic Morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Lipscomb, D.L. Winclada, a Windows Program for Creating, Editing, and Analyzing Systematic Data Sets: Basic Users Guide; George Washington University, Ed.; George Washington University: Washington, DC, USA, 2002. [Google Scholar]
- Hodkinson, I.D. Life Cycle Variation and Adaptation in Jumping Plant Lice (Insecta: Hemiptera: Psylloidea): A Global Synthesis. J. Nat. Hist. 2009, 43, 65–179. [Google Scholar] [CrossRef]
- Burckhardt, D.; Lauterer, P. A Taxonomic Reassessment of the Triozid Genus Bactericera (Hemiptera: Psylloidea). J. Nat. Hist. 1997, 31, 99–153. [Google Scholar] [CrossRef]
- Burckhardt, D.; Cort, G.D.; Queiroz, D.L. Jumping Plant Lice of the Genus Aphalara (Hemiptera, Psylloidea, Aphalaridae) in the Neotropics. Zookeys 2020, 980, 119–140. [Google Scholar] [CrossRef] [PubMed]
- Hodkinson, I.D. New World Legume-Feeding Psyllids of the Genus Aphalaroida Crawford (Insecta: Homoptera: Psylloidea). J. Nat. Hist. 1991, 25, 1281–1296. [Google Scholar] [CrossRef]
- Burckhardt, D.; Wyniger, D. The Systematic Position of Psylla rhorodendri Tuthill with Comments on the New World Genus Freysuila Aleman (Hemiptera, Psylloidea, Aphalaroidinae). Mitteilungen Schweiz. Entomol. Ges. 2007, 80, 63–70. [Google Scholar] [CrossRef]
- Burckhardt, D.; Ouvrard, D. The Temperate Neotropical Psyllid Genus Sphinia (Hemiptera, Psylloidea). Rev. Fr. d’entomologie 2001, 23, 35–46. [Google Scholar]
- Serbina, L.; Burckhardt, D. Systematics, Biogeography and Host-Plant Relationships of the Neotropical Jumping Plant-Louse Genus Russelliana (Hemiptera: Psylloidea). Zootaxa 2017, 4266, 1–114. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burckhardt, D.; Queiroz, D.L. Thought to Be Extinct, but Still Alive Today: The Miocene Genus Primascena Klimaszewsi, 1997 (Hemiptera: Psyllidae) in the Light of Two Extant Species from Brazil. Insects 2024, 15, 382. https://doi.org/10.3390/insects15060382
Burckhardt D, Queiroz DL. Thought to Be Extinct, but Still Alive Today: The Miocene Genus Primascena Klimaszewsi, 1997 (Hemiptera: Psyllidae) in the Light of Two Extant Species from Brazil. Insects. 2024; 15(6):382. https://doi.org/10.3390/insects15060382
Chicago/Turabian StyleBurckhardt, Daniel, and Dalva L. Queiroz. 2024. "Thought to Be Extinct, but Still Alive Today: The Miocene Genus Primascena Klimaszewsi, 1997 (Hemiptera: Psyllidae) in the Light of Two Extant Species from Brazil" Insects 15, no. 6: 382. https://doi.org/10.3390/insects15060382
APA StyleBurckhardt, D., & Queiroz, D. L. (2024). Thought to Be Extinct, but Still Alive Today: The Miocene Genus Primascena Klimaszewsi, 1997 (Hemiptera: Psyllidae) in the Light of Two Extant Species from Brazil. Insects, 15(6), 382. https://doi.org/10.3390/insects15060382