

Field Evaluation of Experimental Maize Hybrids for Resistance to the Fall Armyworm (Lepidoptera: Noctuidae) in a Warm Temperate Climate

 , , ,
, , ,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parental Selection for the Breeding Crosses
2.2. Sources of Insects
2.3. Artificial S. frugiperda Infestation and Injury Rating at the Seedling (Whorl) Stage
2.4. Predator Abundance Survey at the Seedling Stage
2.5. Study Site, Experimental Design, and Data Analysis
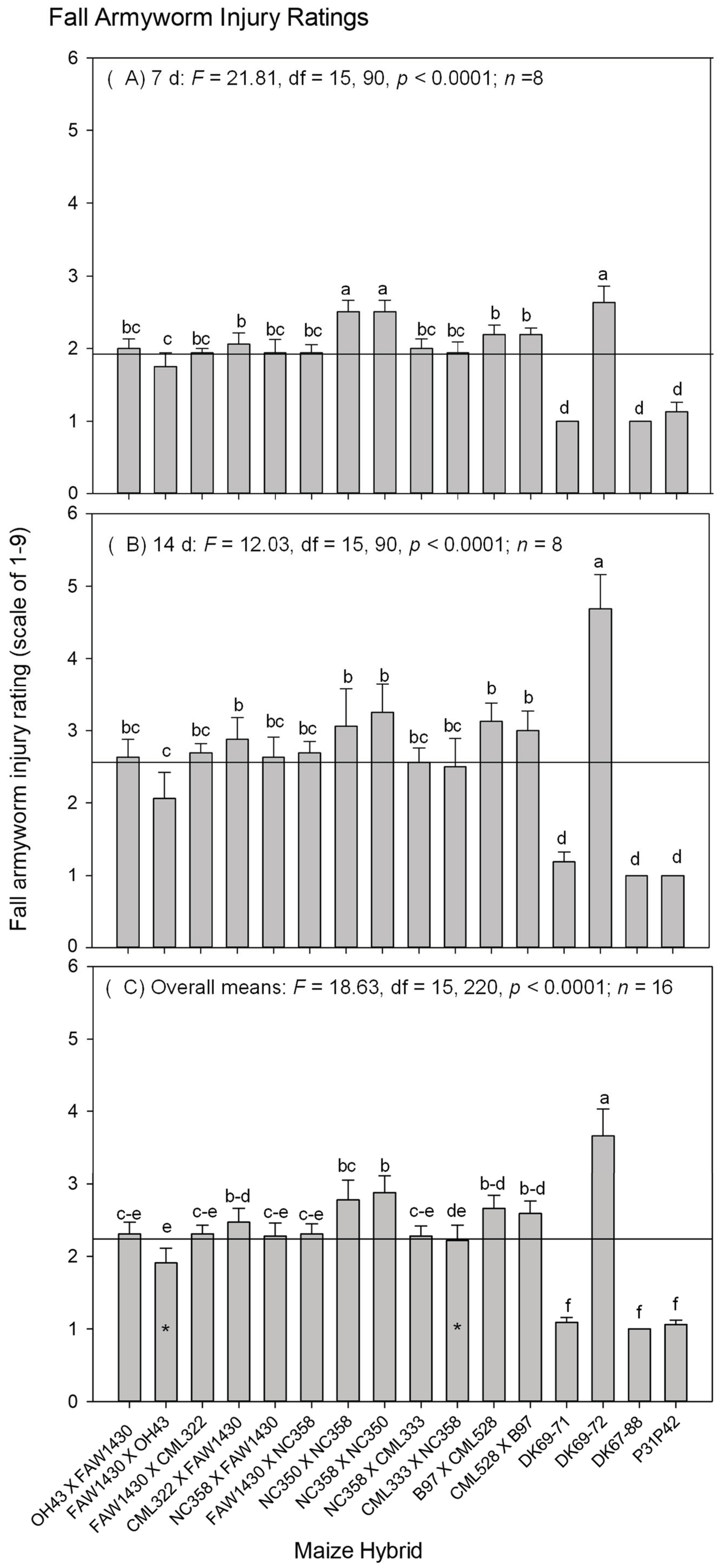
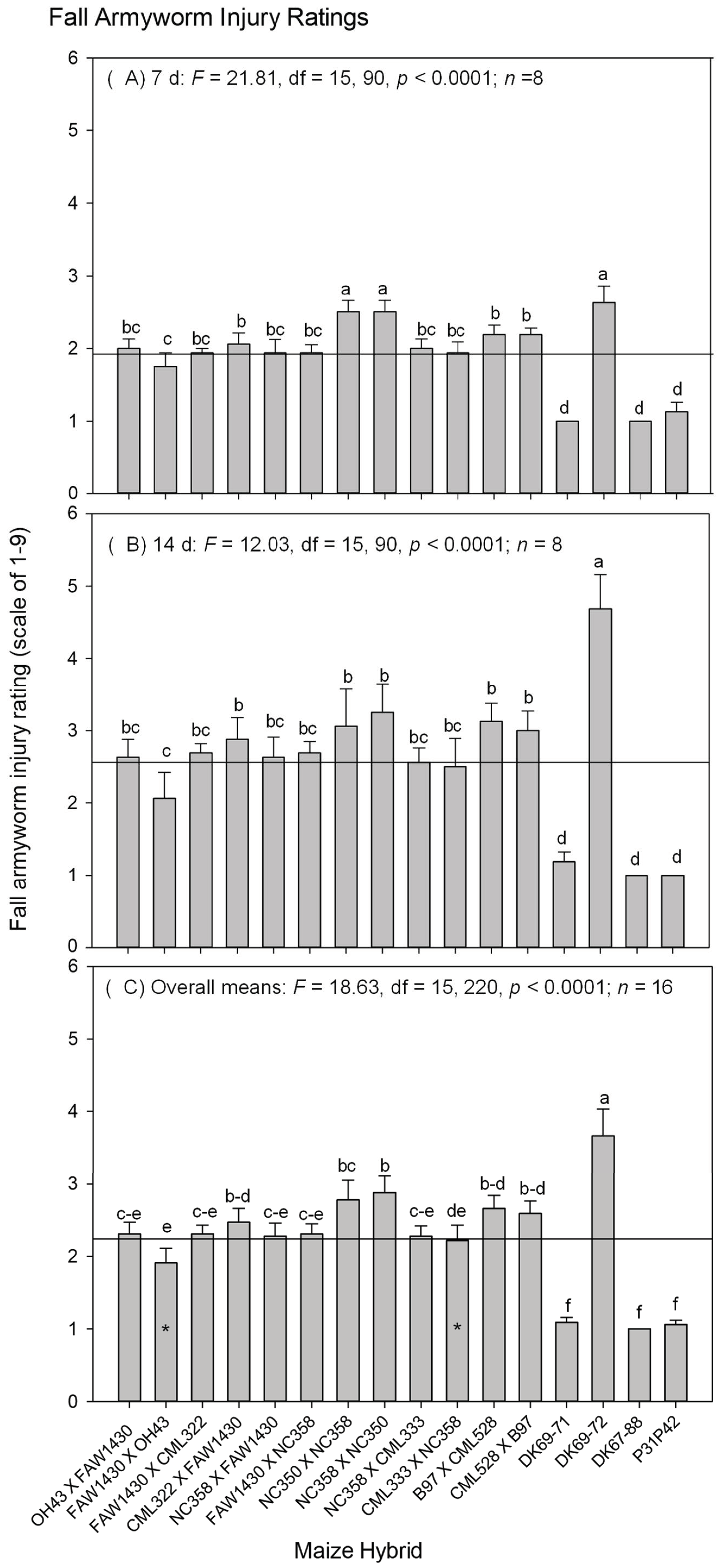
3. Results
3.1. The S. frugiperda Injury at the Seedling Stage
3.2. Predator Abundance and Diversity Survey
3.3. Correlation between S. frugiperda Injury Ratings and among the Predators
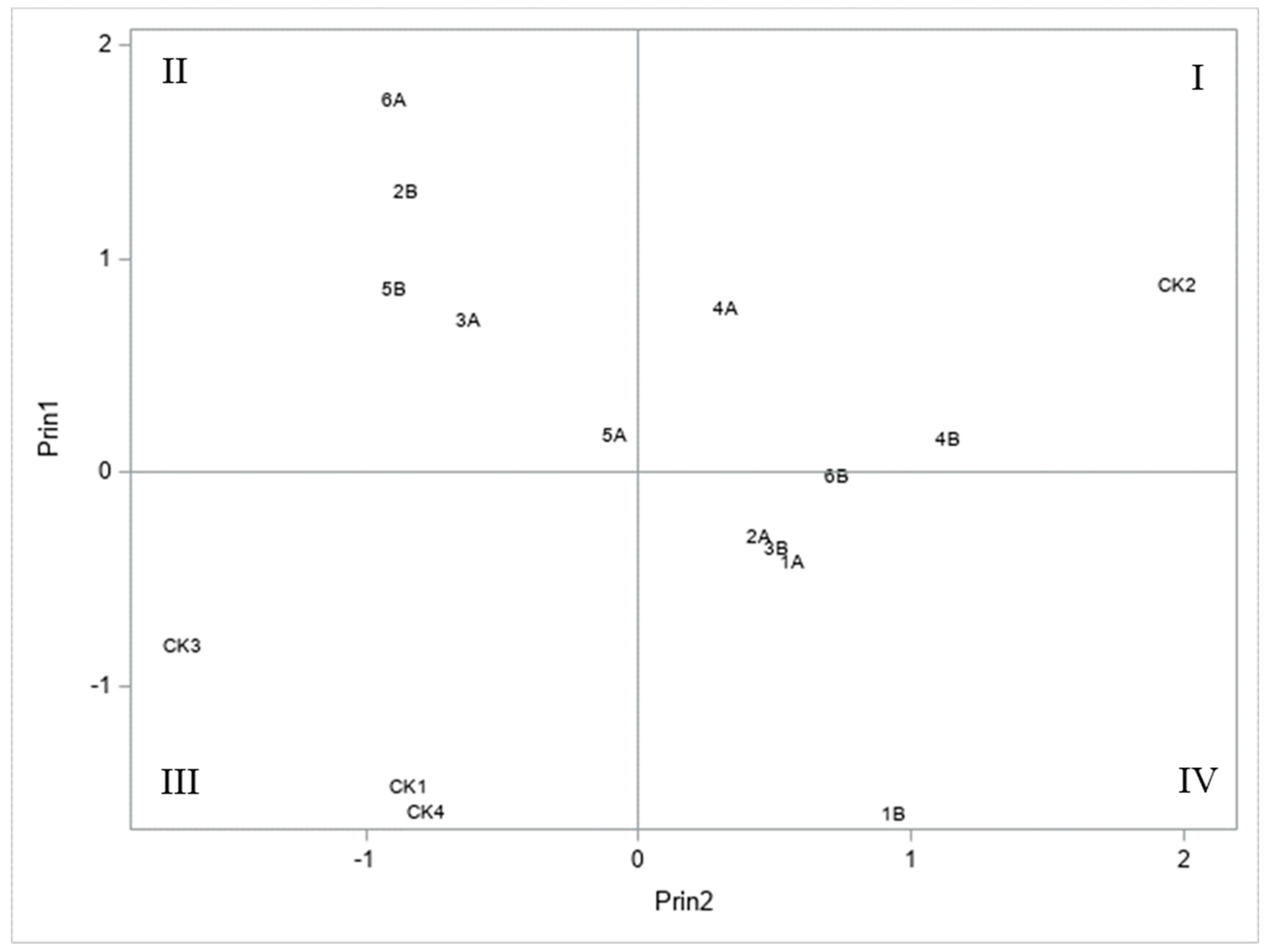
3.4. Principal Component Analysis of S. frugiperda Resistance
3.5. Maternal/Cytoplasmic Effect on Spodoptera frugiperda Resistance in Reciprocal Crosses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burchfield, E.K. Shifting cultivation geographies in the Central and Eastern US. Environ. Res. Lett. 2022, 17, 054049. [Google Scholar] [CrossRef]
- Thome, C.R.; Smith, M.E.; Mihm, J.A. Leaf feeding resistance to multiple insect species in a maize diallel. Crop Sci. 1992, 32, 1460–1463. [Google Scholar] [CrossRef]
- Abel, C.A.; Wilson, R.L.; Wiseman, B.R. Conventional resistance of experimental maize lines to the corn earworm (Lepidoptera: Noctuidae), the fall armyworm (Lepidoptera: Noctuidae), the southwestern corn borer (Lepidoptera: Crambidae), and the sugarcane borer (Lepidoptera: Crambidae). J. Econ. Entomol. 2000, 93, 982–988. [Google Scholar] [CrossRef]
- Ni, X.; Xu, W.; Krakowsky, M.D.; Buntin, G.D.; Brown, S.L.; Lee, R.D.; Coy, A.E. Field screening of experimental maize hybrids and inbred lines for multiple ear-feeding insect resistance. J. Econ. Entomol. 2007, 100, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Krakowsky, M.D.; Buntin, G.D.; Rector, B.G.; Guo, B.; Snook, M.E. Identification of multiple ear-colonizing insect and disease resistance in CIMMYT maize inbred lines with varying levels of silk maysin. J. Econ. Entomol. 2008, 101, 1455–1465. [Google Scholar] [CrossRef]
- Ni, X.; Chen, Y.; Hibbard, B.E.; Wilson, J.P.; Williams, W.P.; Buntin, G.D.; Ruberson, J.R.; Li, X. Foliar resistance to fall armyworm in corn germplasm lines that confer resistance to root- and ear-feeding insects. Fla. Entomol. 2011, 94, 971–981. [Google Scholar] [CrossRef]
- Pring, R.D.; Lonsdale, M. Cytoplasmic male sterility and maternal inheritance of disease susceptibility in maize. Ann. Rev. Phytopath. 1989, 27, 483–502. [Google Scholar] [CrossRef]
- Habarurema, I.; Asea, G.; Lamo, J.; Gibson, P.; Edema, R.; Séré, Y.; Onasanya, R.O. Genetic analysis of resistance to rice bacterial blight in Uganda. Afr. Crop Sci. J. 2012, 20, 105–112. [Google Scholar]
- Hassine, M.; Bnejdi, F.; Bahri, B.A.; Tissaoui, S.; Mougou-Hamdane, A.; Guesmi, M.; Baraket, M.; Slim-Amara, H. Detection of maternal and cytoplasmic effects on resistance to Zymoseptoria tritici in durum wheat. BioMed Res. Int. 2022, 8497417. [Google Scholar] [CrossRef]
- Valverde Caballero, P.; Trapero Ramírez, C.; Barranco Navero, D.; López-Escudero, F.J.; Gordon Bermúdez-Coronel, A.; Díez, C.M. Assessment of maternal effects and genetic variability in resistance to Verticillium dahliae in olive progenies. Plants 2021, 10, 1534. [Google Scholar] [CrossRef]
- Welz, H.G.; Geiger, H.H. Genes for resistance to northern corn leaf blight in diverse maize populations. Plant Breed. 2000, 119, 1–14. [Google Scholar] [CrossRef]
- García-Lara, S.; Bergvinson, D.J.; Burt, A.J.; Ramputh, A.I.; Díaz-Pontones, D.M.; Arnason, J.T. The role of pericarp cell wall components in maize weevil resistance. Crop Sci. 2004, 44, 1546–1552. [Google Scholar] [CrossRef]
- Widstrom, N.W.; Snook, M.E. Genetic variation for maysin and its analogues in crosses among corn inbreds. Crop Sci. 1998, 38, 372–375. [Google Scholar] [CrossRef]
- Bohn, M.; Groh, S.; Khairallah, M.M.; Hoisington, D.A.; Utz, H.F.; Melchinger, A.E. Re-evaluation of the prospects of marker-assisted selection for improving insect resistance against Diatraea spp. in tropical maize by cross validation and independent validation. Theor. Appl. Genet. 2001, 103, 1059–1067. [Google Scholar] [CrossRef]
- Cardinal, A.J.; Lee, M.; Sharopova, N.; Woodman-Clickeman, W.L.; Long, M.J. Genetic mapping and analysis of quantitative trait loci for resistance to stalk tunneling by the European corn borer in maize. Crop Sci. 2001, 41, 835–845. [Google Scholar] [CrossRef]
- Brooks, T.D.; Bushman, B.S.; Williams, W.P.; McMullen, M.D.; Buckley, P.M. Genetic basis of resistance to fall armyworm (Lepidoptera: Noctuidae) and southwestern corn borer (Lepidoptera: Crambidae) leaf-feeding damage in maize. J. Econ. Entomol. 2007, 100, 1470–1475. [Google Scholar] [CrossRef] [PubMed]
- Nuambote-Yobila, O.; Bruce, A.Y.; Okuku, G.O.; Marangu, C.; Makumbi, D.; Beyene, Y.; Mahungu, N.-M.; Prasanna, B.M.; Marion-Poll, F.; Calatayud, P.-A. Assessment of resistance mechanisms to fall armyworm, Spodoptera frugiperda in tropical maize inbred lines. Agronomy 2003, 13, 203. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Kaplan, F.; Huffaker, A.; Dafoe, N.J.; Vaughan, M.M.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E.A. Identity, regulation and activity of inducible diterpenoid phytoalexins in maize. Proc. Natl. Acad. Sci. USA 2011, 108, 5455–5460. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Kaplan, F.; Vaughan, M.M.; Dafoe, N.J.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E.A.; Schmelz, E.A. Novel acidic sesquiterpenoids constitute a dominant class of pathogen-induced phytoalexins in maize. Plant Physiol. 2011, 156, 2082–2097. [Google Scholar] [CrossRef]
- Ni, X.; Xu, W.; Blanco, M.H.; Williams, W.P. Evaluation of fall armyworm resistance in maize germplasm lines using visual leaf injury rating and predator survey. Insect Sci. 2014, 21, 541–555. [Google Scholar] [CrossRef]
- Johnson, E.T.; Skory, C.; Dowd, P.F. Identification of a bioactive Bowman-Birk Inhibitor from an insect-resistant early maize inbred. J. Agric. Food Chem. 2014, 62, 5458–5465. [Google Scholar] [CrossRef] [PubMed]
- Widstrom, N.W.; Williams, W.P.; Wiseman, B.R.; Davis, F.M. Registration of GT-FAWCC(C5) maize germplasm. Crop Sci. 1993, 33, 1422. [Google Scholar] [CrossRef]
- Betran, F.J.; Isakeit, T.; Odvody, G. Aflatoxin accumulation of white and yellow maize inbreds in diallel crosses. Crop Sci. 2002, 42, 1894–1901. [Google Scholar] [CrossRef]
- Cook, J.P.; McMullen, M.D.; Holland, J.B.; Tian, F.; Bradbury, P.; Ross-Ibarra, J.; Buckler, E.S.; Flint-Garcia, S.A. Genetic architecture of maize kernel composition in the nested association mapping and inbred association panels. Plant Physiol. 2012, 158, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, B.R.; Davis, F.M.; Williams, W.P.; Widstrom, N.W. Resistance of a maize population, FAWCC(C5), to fall armyworm larvae (Lepidoptera: Noctuidae). Fla. Entomol. 1996, 79, 329–336. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J. Evaluation of plant resistance to the maize stalk borer, Busseola fusca (Fuller) in maize germplasm improved for resistance to other borer species. South Afr. J. Plant Soil 1998, 15, 34–37. [Google Scholar] [CrossRef]
- Song, K.; Kim, K.; Kim, H.; Moon, J.; Kim, J.; Baek, S.; Kwon, Y.; Lee, B. Evaluation of drought tolerance in maize seedling using leaf rolling. Korean J. Crop Sci. 2015, 60, 8–16. [Google Scholar] [CrossRef]
- Davis, F.M.; Ng, S.S.; Williams, W.P. Visual rating scales for screening whorl-stage corn for resistance to fall armyworm. Tech. Bull. Miss. Agric. For. Exp. Stn. 1992, 186, 9. [Google Scholar]
- Davis, F.M.; Wiseman, B.R.; Williams, W.P.; Widstrom, N.W. Insect colony, planting date, and plant growth stage effects on screening maize for leaf-feeding resistance to fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 1996, 79, 317–328. [Google Scholar] [CrossRef]
- Smith, C.M.; Khan, Z.R.; Pathak, M.D. Evaluation of plants for insect resistance. In Techniques for Evaluating Insect Resistance in Crop Plants; Smith, C.M., Khan, Z.R., Pathak, M.D., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 17–114. [Google Scholar]
- Meagher, R.L.; Nuessly, G.S.; Nagoshi, R.N.; Hay-Roe, M.M. Parasitoids attacking fall armyworm (Lepidoptera: Noctuidae) in sweet corn habitats. Biol. Control 2016, 95, 66–72. [Google Scholar] [CrossRef]
- Abbas, A.; Ullah, F.; Hafeez, M.; Han, X.; Dara, M.Z.N.; Gul, H.; Zhao, C.R. Biological control of fall armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2704. [Google Scholar] [CrossRef]
- Pal, S.; Bhattacharya, S.; Dhar, T.; Gupta, A.; Ghosh, A.; Debnath, S.; Gangavarapu, N.; Pati, P.; Chaudhuri, N.; Chatterjee, H.; et al. Hymenopteran parasitoid complex and fall armyworm: A case study in eastern India. Sci. Rep. 2024, 14, 4029. [Google Scholar] [CrossRef]
- Der, G.; Everitt, B.S. Principal components analysis and factor analysis: Olympic decathlon and statements about pain. In A Handbook of Statistical Analyses Using SAS, 3rd ed.; CRC Press: Boca Raton, FL, USA, 1991; pp. 295–315. [Google Scholar]
- Hoballah, M.E.; Degen, T.; Bergvinson, D.; Savidan, A.; Tamò, C.; Turlings, T.C.J. Occurrence and direct control potential of parasitoids and predators of the fall armyworm (Lepidoptera: Noctuidae) on maize in the subtropical lowlands of Mexico. Agric. For. Entomol. 2004, 6, 83–88. [Google Scholar] [CrossRef]
- Sueldo, M.R.; Bruzzone, O.A.; Virla, E.G. Characterization of the earwig, Doru lineare, as a predator of larvae of the fall armyworm, Spodoptera frugiperda: A functional response study. J. Insect Sci. 2010, 10, 38. [Google Scholar] [CrossRef]
- Silva, R.B.; Cruz, I.; Zanuncio, J.C.; Figueiredo, M.D.L.C.; Zanuncio, T.V.; Serrão, J.E. Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) eggs as alternative food for rearing of lady beetles Eriopis connexa (Germar) (Coleoptera: Coccinellidae). Biol. Control 2013, 64, 101–105. [Google Scholar] [CrossRef]
- Silva, G.A.; Picanço, M.C.; Ferreira, L.R.; Ferreira, D.O.; Farias, E.S.; Souza, T.C.; Rodrigues-Silva, N.; Pereira, E.J.G. Yield losses in transgenic Cry1Ab and non-Bt corn as assessed using a crop-life-table approach. J. Econ. Entomol. 2018, 111, 218–226. [Google Scholar] [CrossRef]
- Smith, W.E.C. Detection and Characterization of a Volatile Compound as a Response to Fall Armyworm (Spodoptera frugiperda) Feeding in Maize (Zea mays). Master’s Thesis, Mississippi State University, Starkville, MS, USA, 2010. [Google Scholar]
- Warburton, M.L.; Woolfolk, S.W.; Smith, J.S.; Hawkins, L.K.; Castano-Duque, L.; Lebar, M.D.; Williams, W.P. Genes and genetic mechanisms contributing to fall armyworm resistance in maize. Plant Genome 2023, 16, e20311. [Google Scholar] [CrossRef]
- Ni, X.; Lei, Z.; He, K.; Li, X.; Li, X.; Xu, W. Integrated pest management is the lucrative bridge connecting the ever-emerging knowledge islands of genetics and ecology. Insect Sci. 2014, 21, 537–540. [Google Scholar] [CrossRef]
- Huang, K.; He, H.; Wang, S.; Zhang, M.; Chen, X.; Deng, Z.; Ni, X.; Li, X. Sequential and simultaneous interactions of plant allelochemical flavone, Bt toxin Vip3A and insecticide emamectin benzoate in Spodoptera Frugiperda. Insects 2023, 14, 736. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Entry | Pedigree (Female × Male) | References |
|---|---|---|
| 1A | Oh43 × FAW1430 | Oh43: [18,19,21] |
| 1B | FAW1430 × Oh43 | FAW1430: derived from FAW CC(C)5 [22] |
| 2A | FAW1430 × CML322 | CML322: [18,23,24] |
| 2B | CML322 × FAW1430 | |
| 3A | NC358 × FAW1430 | NC358: [18,19] |
| 3B | FAW1430 × NC358 | |
| 4A | NC350 × NC358 | NC350: [18,19] |
| 4B | NC358 × NC350 | |
| 5A | NC358 × CML333 | CML333: [18,19] |
| 5B | CML333 × NC358 | |
| 6A | B97 × CML528 | B97: [18,19] |
| 6B | CML528 × B97 | CML528: tropical inbred (procured from CIMMYT in 2011) |
| CK1 | DKC69-71 | Commercial check, Bt transgenic (Bayer, Monsanto) |
| CK2 | DKC69-72 | Commercial check, non-Bt control (Bayer, Monsanto) |
| CK3 | DKC67-88 | Commercial check, Bt transgenic (Bayer, Monsanto) |
| CK4 | P31P42 | Commercial check, Bt transgenic (DuPont, Pioneer) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, X.; Huffaker, A.; Schmelz, E.A.; Xu, W.; Williams, W.P.; Guo, B.; Li, X.; Huang, F. Field Evaluation of Experimental Maize Hybrids for Resistance to the Fall Armyworm (Lepidoptera: Noctuidae) in a Warm Temperate Climate. Insects 2024, 15, 289. https://doi.org/10.3390/insects15040289
Ni X, Huffaker A, Schmelz EA, Xu W, Williams WP, Guo B, Li X, Huang F. Field Evaluation of Experimental Maize Hybrids for Resistance to the Fall Armyworm (Lepidoptera: Noctuidae) in a Warm Temperate Climate. Insects. 2024; 15(4):289. https://doi.org/10.3390/insects15040289
Chicago/Turabian StyleNi, Xinzhi, Alisa Huffaker, Eric A. Schmelz, Wenwei Xu, W. Paul Williams, Baozhu Guo, Xianchun Li, and Fangneng Huang. 2024. "Field Evaluation of Experimental Maize Hybrids for Resistance to the Fall Armyworm (Lepidoptera: Noctuidae) in a Warm Temperate Climate" Insects 15, no. 4: 289. https://doi.org/10.3390/insects15040289
APA StyleNi, X., Huffaker, A., Schmelz, E. A., Xu, W., Williams, W. P., Guo, B., Li, X., & Huang, F. (2024). Field Evaluation of Experimental Maize Hybrids for Resistance to the Fall Armyworm (Lepidoptera: Noctuidae) in a Warm Temperate Climate. Insects, 15(4), 289. https://doi.org/10.3390/insects15040289