
Impact of Chemical Insecticide Application on Beneficial Insects in Maize


Abstract
Simple Summary
Abstract
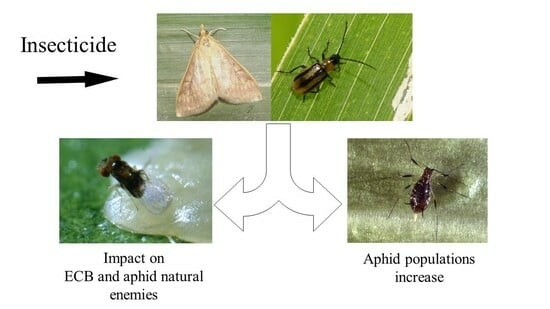
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Fieldwork Methods
2.2.1. Aphids
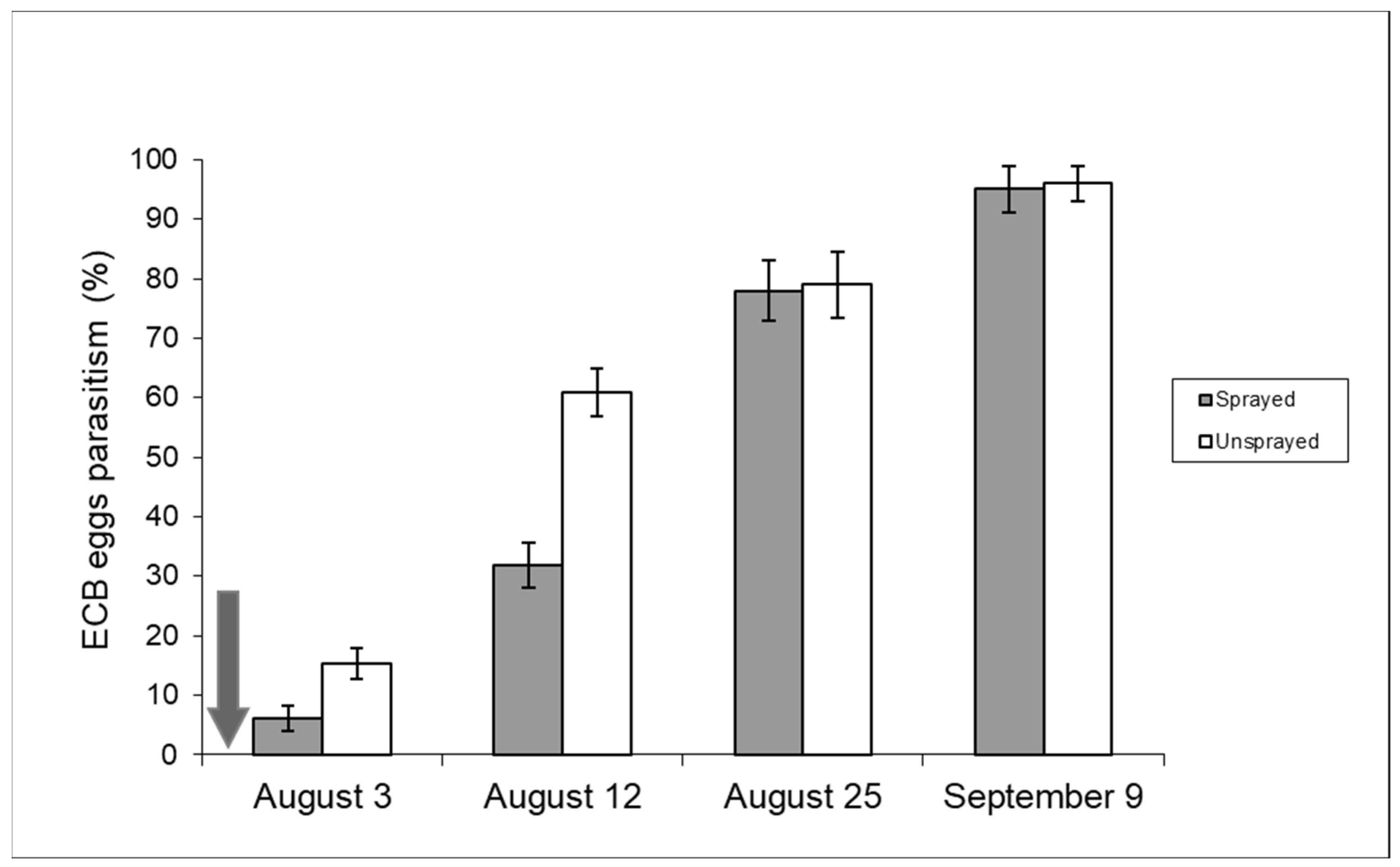
2.2.2. ECB Egg Masses Parasitism by Trichogramma brassicae
2.3. Statistical Analysis
3. Results
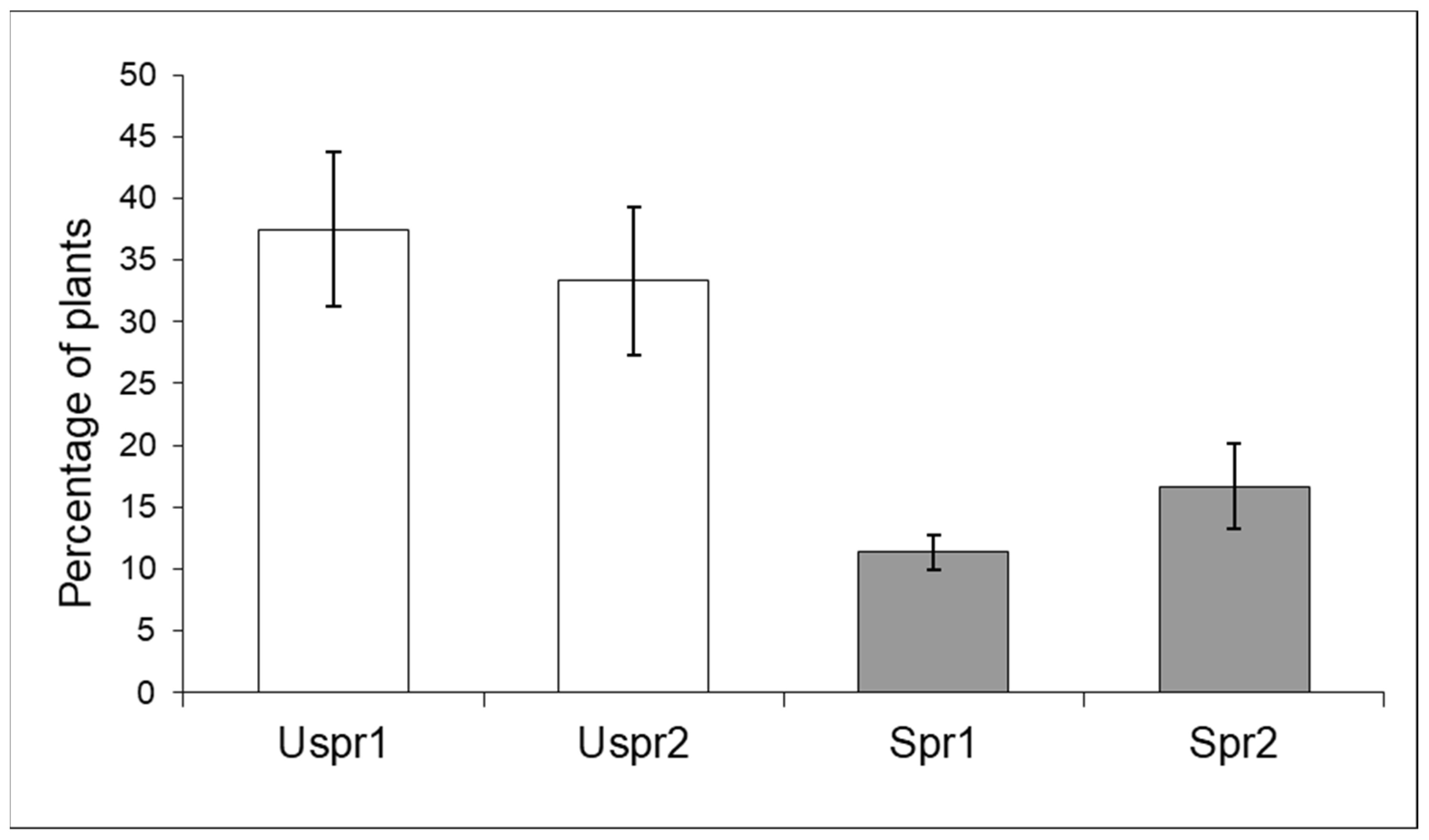
3.1. Impact of Insecticides on Aphids and Their Predators
3.1.1. Aphid Community Composition
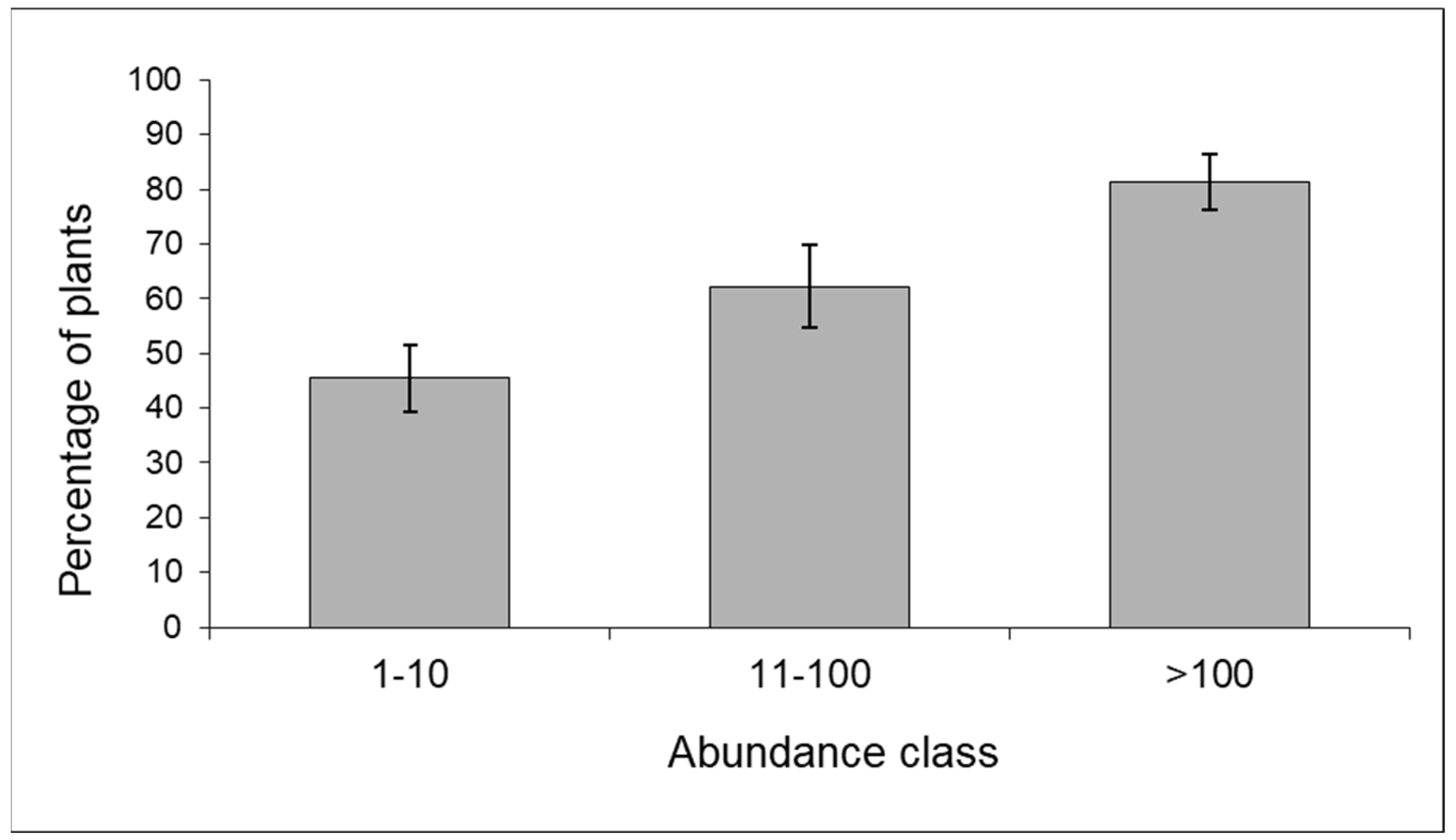
3.1.2. Aphids on Maize and Their Predators (Years 2008–2010)
3.1.3. Predator Community: Composition and Response to Aphid Density
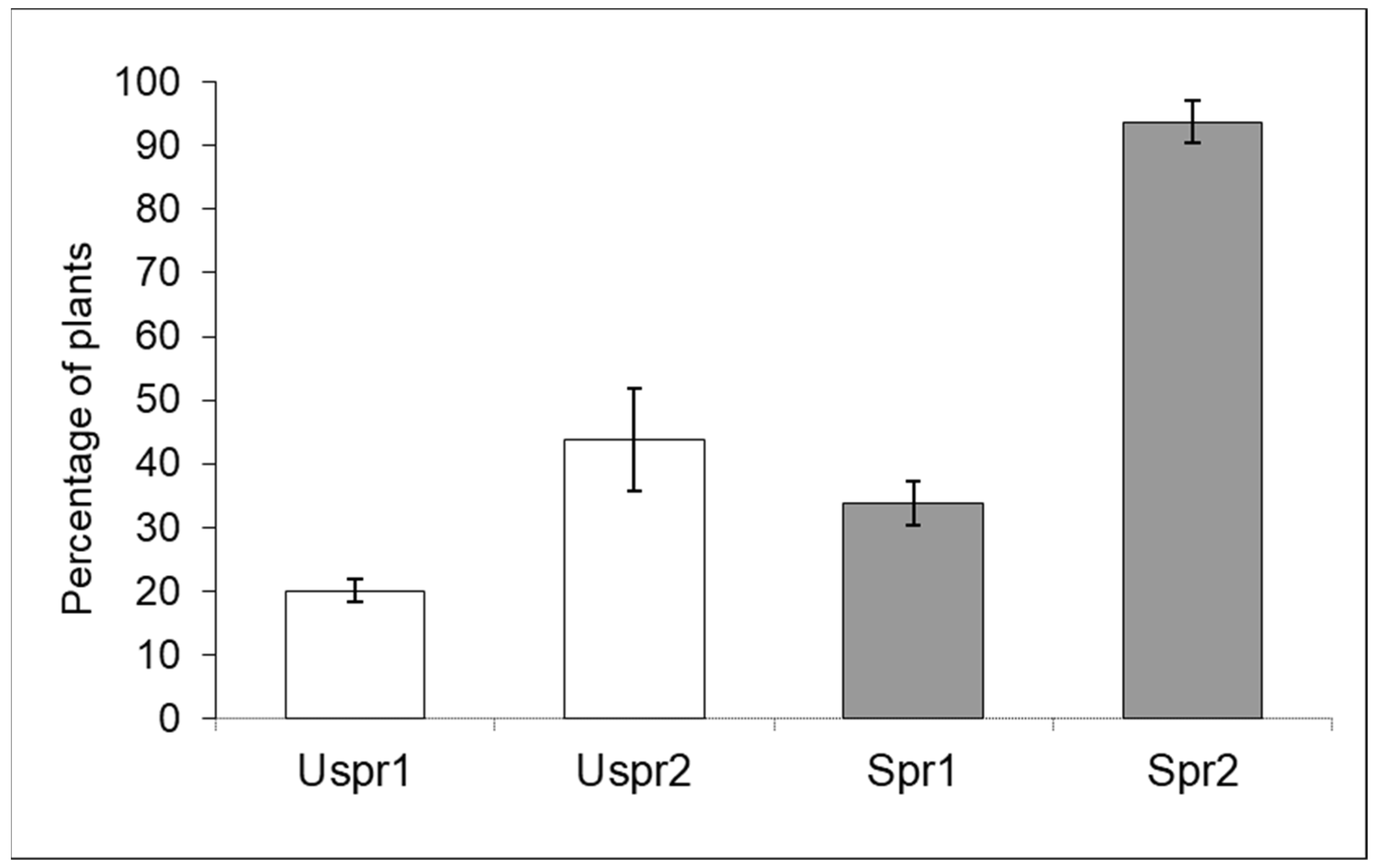
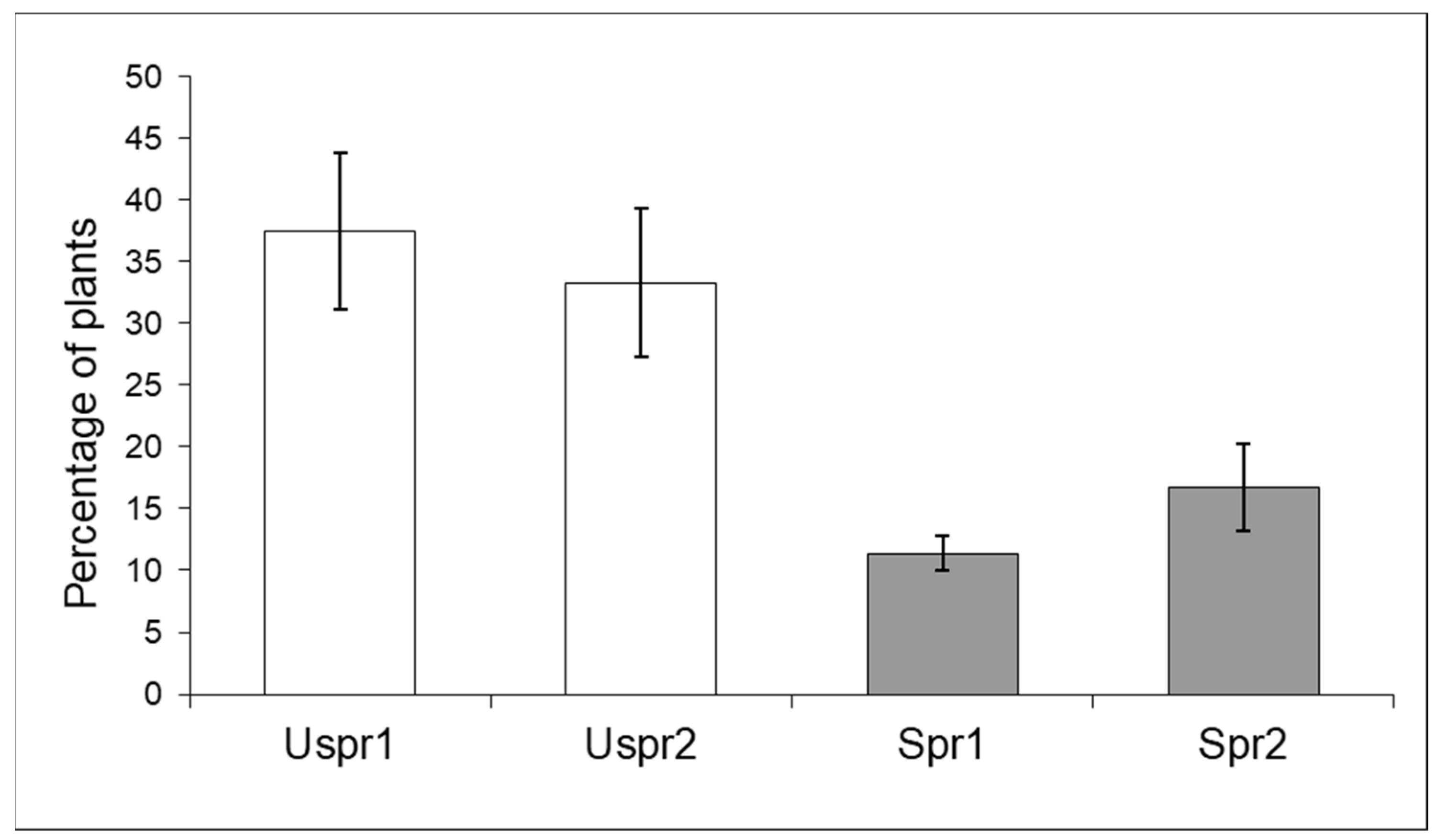
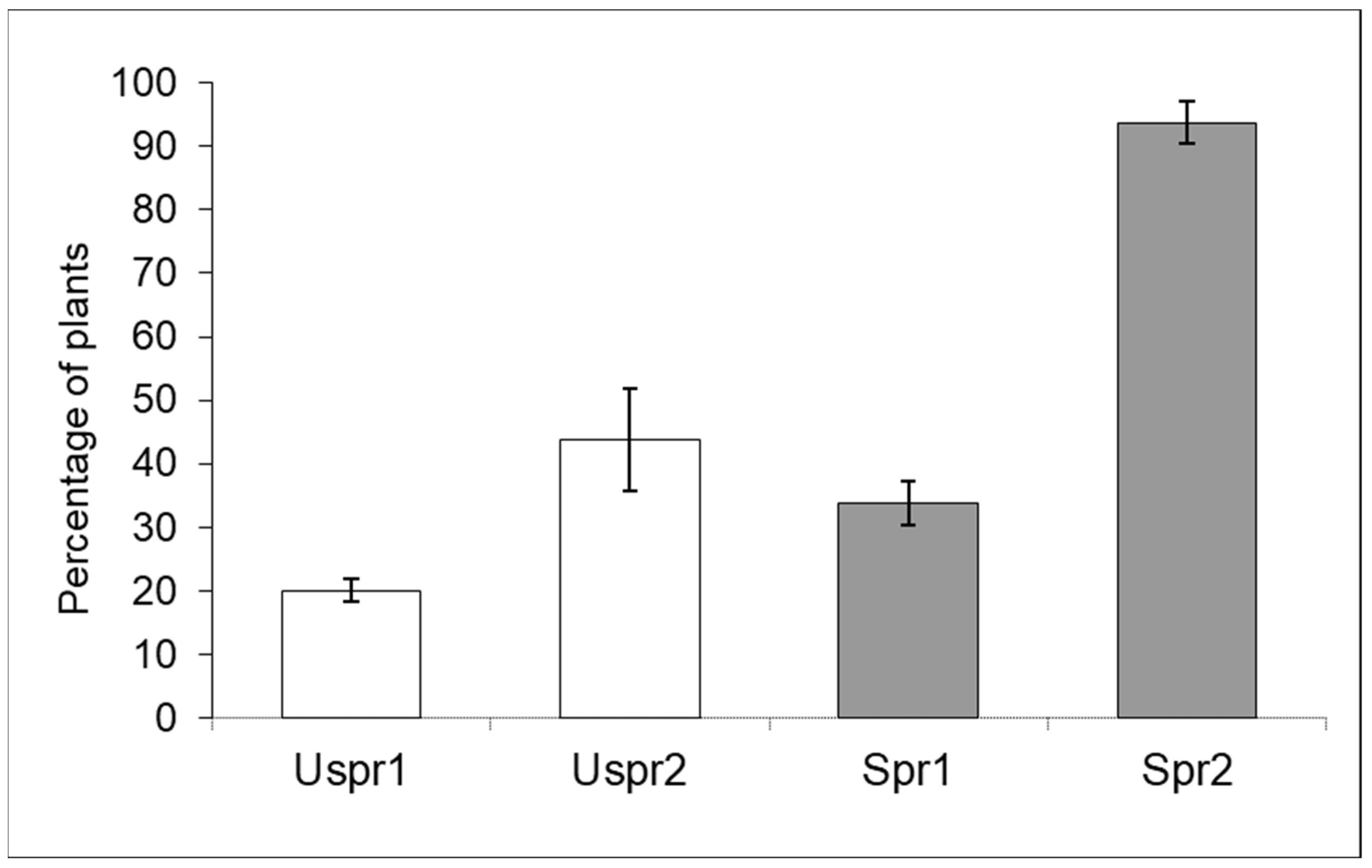
3.2. Impact of Insecticides on Trichogramma brassicae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magagnoli, S.; Lanzoni, A.; Masetti, A.; Depalo, L.; Albertini, M.; Ferrari, R.; Spadola, G.; Degola, F.; Restivo, F.M.; Burgio, G. Sustainability of strategies for Ostrinia nubilalis management in Northern Italy: Potential impact on beneficial arthropods and aflatoxin contamination in years with different meteorological conditions. Crop Prot. 2021, 142, 105529. [Google Scholar] [CrossRef]
- Burgio, G.; Maini, S. Control of European corn borer in sweet corn by Trichogramma brassicae Bezd. (Hym., Trichogrammatidae). J. Appl. Entomol. 1995, 119, 83–87. [Google Scholar] [CrossRef]
- Camerini, G.; Groppali, R.; Tschorsnig, H.P.; Maini, S. Influence of Ostrinia nubilalis larval density and location in the maize plant on the tachinid fly Lydella thompsoni. Bull. Insectol. 2016, 69, 301–306. [Google Scholar]
- Camerini, G.; Masanta, M.; Maini, S. Ostrinia nubilalis in northern Italy: Egg parasitoids and control trials on three sweet pepper landraces in open field cultivations. Bull. Insectol. 2023, 76, 305–313. [Google Scholar]
- Camerini, G.; Franzini, L.; Groppali, R. Response of spiders (Araneae) to the introduction of an exotic pest (Diabrotica virgifera LeConte, 1868-Coleoptera, Chrysomelidae) in corn cultivations in Northern Italy. Maydica 2021, 66, 9. [Google Scholar]
- Furlan, L.; Chiarini, F.; Contiero, B.; Benvegnù, I.; Horgan, F.G.; Kos, T.; Lemić, D.; Bažok, R. Risk assessment and area-wide crop rotation to keep Western Corn Rootworm below damage thresholds and avoid insecticide use in european maize production. Insects 2022, 13, 415. [Google Scholar] [CrossRef] [PubMed]
- Toepfer, S.; Kiss, J.; Turoczi, G.; Komaromi, J.; Kuhlmann, U. Options for developing integrated control measures against the maize pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) in Europe. IOBC/WPRS Bull. 2005, 28, 1–7. [Google Scholar]
- Maini, S.; Burgio, G. Ostrinia nubilalis on sweet corn: Relationship between adults caught in multibaited traps and ear damages. J. Appl. Entomol. 1999, 123, 179–185. [Google Scholar] [CrossRef]
- Meissle, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z. Pests, pesticide use and alternative options in European maize production: Current status and future prospects. J. Appl. Entomol. 2010, 134, 357–375. [Google Scholar] [CrossRef]
- Schaub, L.; Furlan, L.; Tóth, M.; Steinger, T.; Carrasco, L.R.; Toepfer, S. Efficiency of pheromone traps for monitoring Diabrotica virgifera virgifera LeConte. OEPP/EPPO Bull. 2011, 41, 189–194. [Google Scholar] [CrossRef]
- Sappington, T.W. Critical facets of European Corn Borer adult movement ecology relevant to mitigating field resistance to Bt-Corn. Insects 2024, 15, 160. [Google Scholar] [CrossRef]
- Meinke, L.J.; Sappington, T.W.; Onstad, D.W.; Guillemaud, T.; Miller, N.J.; Komáromi, J.; Levay, N.; Furlan, L.; Kiss, J.; Toth, F. Western corn rootworm (Diabrotica virgifera virgifera LeConte) population dynamics. Agr. Forest Entom. 2009, 11, 29–46. [Google Scholar] [CrossRef]
- Pintilie, P.L.; Tălmaciu, M.; Trotuș, E.; Amarghioalei, R.G.; Leonte, A.; Isticioaia, S.F. Parameters analysis of the Ostrinia nubilalis Hbn. attack at maize crops in the conditions of Central Moldova. Sci. Pap. Ser. A Agron. 2022, 65, 482–490. [Google Scholar]
- Camerini, G.; Groppali, R.; Rama, F.; Maini, S. Semiochemicals of Ostrinia nubilalis: Diel response to sex pheromone and phenylacetaldehyde in open field. Bull. Insectol. 2015, 68, 45–50. [Google Scholar]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef] [PubMed]
- Pintureau, B. Les Espèces Europèennes de Trichogrammes; InLibroVeritas: Angers, France, 2008; pp. 1–96. [Google Scholar]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status, and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Caballero-López, B.; Bommarco, R.; Blanco-Moreno, J.M.; Sans, F.X.; Pujade-Villar, J.; Rundlöf, M.; Smith, H.G. Aphids and their natural enemies are differently affected by habitat features at local and landscape scales. Biol. Control 2012, 63, 222–229. [Google Scholar] [CrossRef]
- Camerini, G.; Maini, S.; Riedel, M. Ostrinia nubilalis parasitoids in Northern Italy: Past and present. Biol. Control 2018, 12, 76–83. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Froment, B.; Fouillet, P.; Pompanon, F.; Janillon, S.; Boulétreau, M. Inhibition of sex pheromone communications of Trichogramma brassicae (Hymenoptera) by the insecticide chlorpyrifos. Environ. Toxicol. Chem. 1998, 17, 1107–1113. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Gareau, E.; Terrier, O.; Fouillet, P. Sublethal effects of the insecticide chlorpyrifos on the sex pheromonal communication of Trichogramma brassicae. Chemosphere 1998, 36, 1775–1785. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Legallet, B.; Terrier, O.; Fouillet, P. Modifications of the sex pheromonal communication of Trichogramma brassicae by a sublethal dose of deltamethrin. Chemosphere 1999, 38, 729–739. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Legallet, B.; Fouillet, P. Partial compensation of the sublethal effect of deltamethrin on the sex pheromonal communication of Trichogramma brassicae. Chemosphere 2001, 42, 985–991. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Meyet, J. Reduction in the sex ratio of the progeny of a parasitoid wasp (Trichogramma brassicae) surviving the insecticide chlorpyrifos. Arch. Environ. Contam. Toxicol. 2003, 45, 203–208. [Google Scholar] [CrossRef]
- Toft, S. Value of the aphid Rhopalosiphum padi as food for cereal spiders. J. Appl. Ecol. 1995, 32, 552–560. [Google Scholar] [CrossRef]
- Dedryver, C.A. Les pucerons dans le rotations cerealieres. Perspect. Agric. 1980, 37, 54–64. [Google Scholar]
- Mestres, R.; Cabanettes, J.P. Effets intentionnels ou non des traitements chimiques dans la lutte contre la pyrale. Phytoma 1985, 369, 19–21. [Google Scholar]
- Vasileiadis, V.P.; Veres, A.; Loddo, D.; Masin, R.; Sattin, M.; Furlan, L. Careful choice of insecticides in integrated pest management strategies against Ostrinia nubilalis (Hübner) in maize conserves Orius spp. in the field. Crop Prot. 2017, 97, 45–51. [Google Scholar] [CrossRef]
- Ko, K.; Liu, Y.; Hou, M.; Babendreier, D.; Zhang, F.; Song, K. Toxicity of insecticides targeting rice planthoppers to adult and immature stages of Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2015, 108, 69–76. [Google Scholar] [CrossRef]
- Afshari, A.; Hamzepour Chenari, E.; Iraji, A.; Asghari Larimi, M. Lethal and sublethal effects of thiodicarb and hexaflumuron on egg parasitoid, Trichogramma brassicae Bezdenko under laboratory conditions. Plant Prot. 2018, 41, 75–89. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Delahaye, M. The sublethal effects of deltamethrin on Trichogramma behaviors during the exploitation of host patches. Sci. Total Environ. 2013, 447, 274–279. [Google Scholar] [CrossRef]
- Hewa-Kapuge, S.; McDougall, S.; Hoffmann, A.A. Effects of methoxyfenozide, indoxacarb, and other insecticides on the beneficial egg parasitoid Trichogramma nr. brassicae (Hymenoptera: Trichogrammatidae) under laboratory and field conditions. J. Econ. Entomol. 2003, 96, 1083. [Google Scholar] [CrossRef]
- Shirazi, J.; Bahrami, A.; Rezapanah, M. Side effects of five pesticides on Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae). J. Agric. For. 2018, 66, 57–68. [Google Scholar]
- Jin, T.; Lin, Y.; Ma, G.; Liu, J.; Hao, Z.; Han, S.; Peng, Z. Biocontrol potential of Trichogramma species against Spodoptera frugiperda and their field efficacy in maize. Crop Prot. 2021, 150, 105790. [Google Scholar] [CrossRef]
- Liu, T.X.; Zhang, Y. Side effects of two reduced-risk insecticides, indoxacarb and spinosad, on two species of Trichogramma (Hymenoptera: Trichogrammatidae) on cabbage. Ecotoxicology 2012, 21, 2254–2263. [Google Scholar] [CrossRef]
- Nozad-Bonab, Z.; Hejazi, M.J.; Iranipour, S.; Arzanlou, M.; Biondi, A. Lethal and sublethal effects of synthetic and bio-insecticides on Trichogramma brassicae parasitizing Tuta absoluta. PLoS ONE 2021, 6, e0243334. [Google Scholar] [CrossRef]
- Parsaeyan, E.; Safavi, S.A.; Saber, M.; Poorjavad, N. Effects of emamectin benzoate and cypermethrin on the demography of Trichogramma brassicae Bezdenko. Crop Prot. 2018, 110, 269–274. [Google Scholar] [CrossRef]
- Parsaeyan, E.; Saber, M.; Safavi, S.A.; Poorjavad, N.; Biondi, A. Side effects of chlorantraniliprole, phosalone and spinosad on the egg parasitoid, Trichogramma brassicae. Ecotoxicology 2020, 29, 1052–1061. [Google Scholar] [CrossRef]
- Saber, M.; Ghorbani, M.; Vaez, N.; Armak, A. Effects of diazinon and fipronil on functional response of Trichogramma brassicae Bezdenko (Hym.; Trichogrammatidae) in the laboratory conditions. J. Crop Prot. 2020, 9, 275–283. [Google Scholar]
- Thubru, D.P.; Firake, D.M.; Behere, G.T. Assessing risks of pesticides targeting lepidopteran pests in cruciferous ecosystems to eggs parasitoid, Trichogramma brassicae (Bezdenko). Saudi J. Biol. Sci. 2018, 25, 680–688. [Google Scholar] [CrossRef]
- Saber, M. Sublethal effects of methoxyfenozide and emamectin benzoate on Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae). Agric. Pest Manag. 2015, 2, 12–20. [Google Scholar]
- Russo, J.; Voegelé, J. Influence de la température sur quatre espèces de trichogrammes (Hym. Trichogrammatidae) parasites de la pyrale du maïs, Ostrinia nubilalis Hubn. (Lep. Pyralidae). I.—Développement préimaginal. Agronomie 1982, 2, 509–516. [Google Scholar] [CrossRef]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? Proc. R. Soc. B 2020, 287, 20201265. [Google Scholar] [CrossRef]
- Helps, J.C.; Paveley, N.D.; Van Den Bosch, F. Identifying circumstances under which high insecticide dose increases or decreases resistance selection. J. Theor. Biol. 2017, 428, 153–167. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- European Commission. Farm to Fork Strategy. 2020, pp. 1–20. Available online: https://food.ec.europa.eu/horizontal-topics/farm-fork-strategy_en (accessed on 5 December 2023).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Area 1 | Area 2 |
|---|---|---|
| Rhopalosiphum padi (L.) | 64.8 | 59.9 |
| Rhopalosiphum maidis (Fitch) | 20.8 | 25.6 |
| Sipha maidis Passerini | 10.4 | 11.3 |
| Sitobion avenae (F.) | 4 | 3.2 |
| Aphis sp. | 0 | 0 |
| Sample | Year | 1–10 | 11–100 | >100 | n |
|---|---|---|---|---|---|
| Edge | 2009 | 30.1 | 24.8 | 45.1 | 133 |
| Inner | 2009 | 27.8 | 42.6 | 29.6 | 108 |
| Edge | 2010 | 64.6 | 30.2 | 5.2 | 96 |
| Inner | 2010 | 71 | 20.4 | 8.6 | 89 |
| Sample | Plants with Aphids (%) | Plants with Predators (%) |
|---|---|---|
| Area 1 | 48.8 ± 6 | 97.1 ± 1.9 |
| Area 2 (Robecco S.N.) | 15.6 ± 1.2 | 61.7 ± 7.8 |
| Area 2 (S. Stefano T.) | 57.5 ± 2.5 | 44 ± 4.9 |
| Predators | Year 2008 | Year 2009 | Year 2010 | |||
|---|---|---|---|---|---|---|
| Area 1 | Area 2 | Area 1 | Area 2 | Area 1 | Area 2 | |
| Ladybirds | 84.3 | 26.4 | 38.4 | 26.4 | 78.3 | 26.7 |
| Orius | 9.1 | 18.5 | 37.8 | 19.2 | 13 | 63.9 |
| Hoverflies | 6.2 | 49.9 | 9.7 | 33.4 | 1.7 | 7.5 |
| Lacewings | 0.3 | 0.2 | 9.4 | 17.7 | 4.4 | 1.9 |
| Others | 0.1 | 5 | 4.7 | 3.3 | 2.6 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camerini, G.; Maini, S.; Limonta, L. Impact of Chemical Insecticide Application on Beneficial Insects in Maize. Insects 2024, 15, 224. https://doi.org/10.3390/insects15040224
Camerini G, Maini S, Limonta L. Impact of Chemical Insecticide Application on Beneficial Insects in Maize. Insects. 2024; 15(4):224. https://doi.org/10.3390/insects15040224
Chicago/Turabian StyleCamerini, Giuseppe, Stefano Maini, and Lidia Limonta. 2024. "Impact of Chemical Insecticide Application on Beneficial Insects in Maize" Insects 15, no. 4: 224. https://doi.org/10.3390/insects15040224
APA StyleCamerini, G., Maini, S., & Limonta, L. (2024). Impact of Chemical Insecticide Application on Beneficial Insects in Maize. Insects, 15(4), 224. https://doi.org/10.3390/insects15040224