Development and Metabolomic Profiles of Bactrocera dorsalis (Diptera: Tephritidae) Larvae Exposed to Phytosanitary Irradiation Dose in Hypoxic Environment Using DI-SPME-GC/MS



Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Modified Atmosphere Treatments
2.3. Irradiation Treatments
2.4. Develpomental Test
2.5. Sample Preparation and Extraction Using DI-SPME
2.6. Metabolites Analysis with GC-MS
2.7. Statistical Analysis
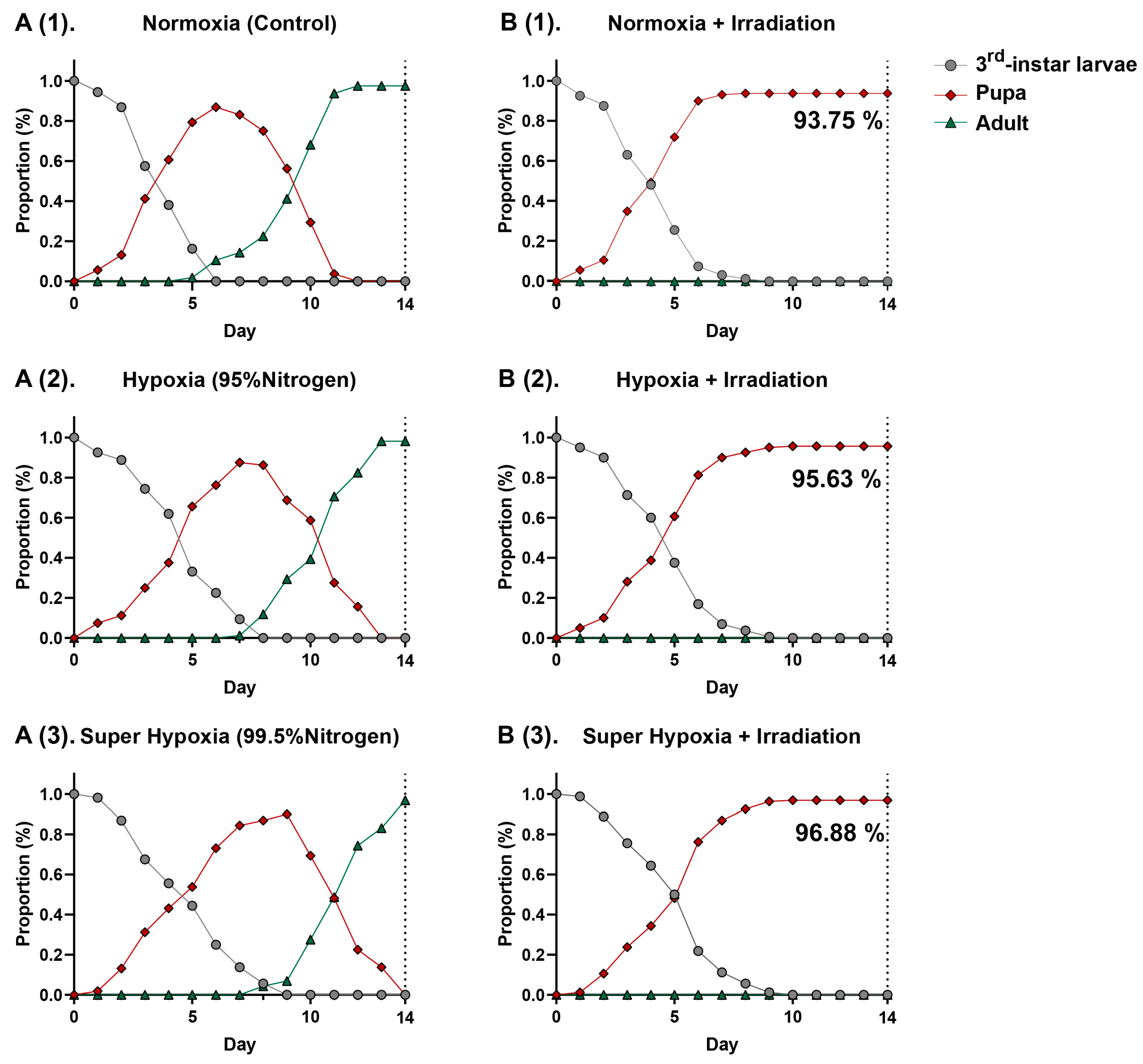
3. Results
3.1. Develpment of Bactrocera dorsalis (Diptera: Tephritidae) Larvae in a Hypoxic Environment with Phytosanitary Irradiation
3.2. Metabolic Profiles Analyzed via GC-MS
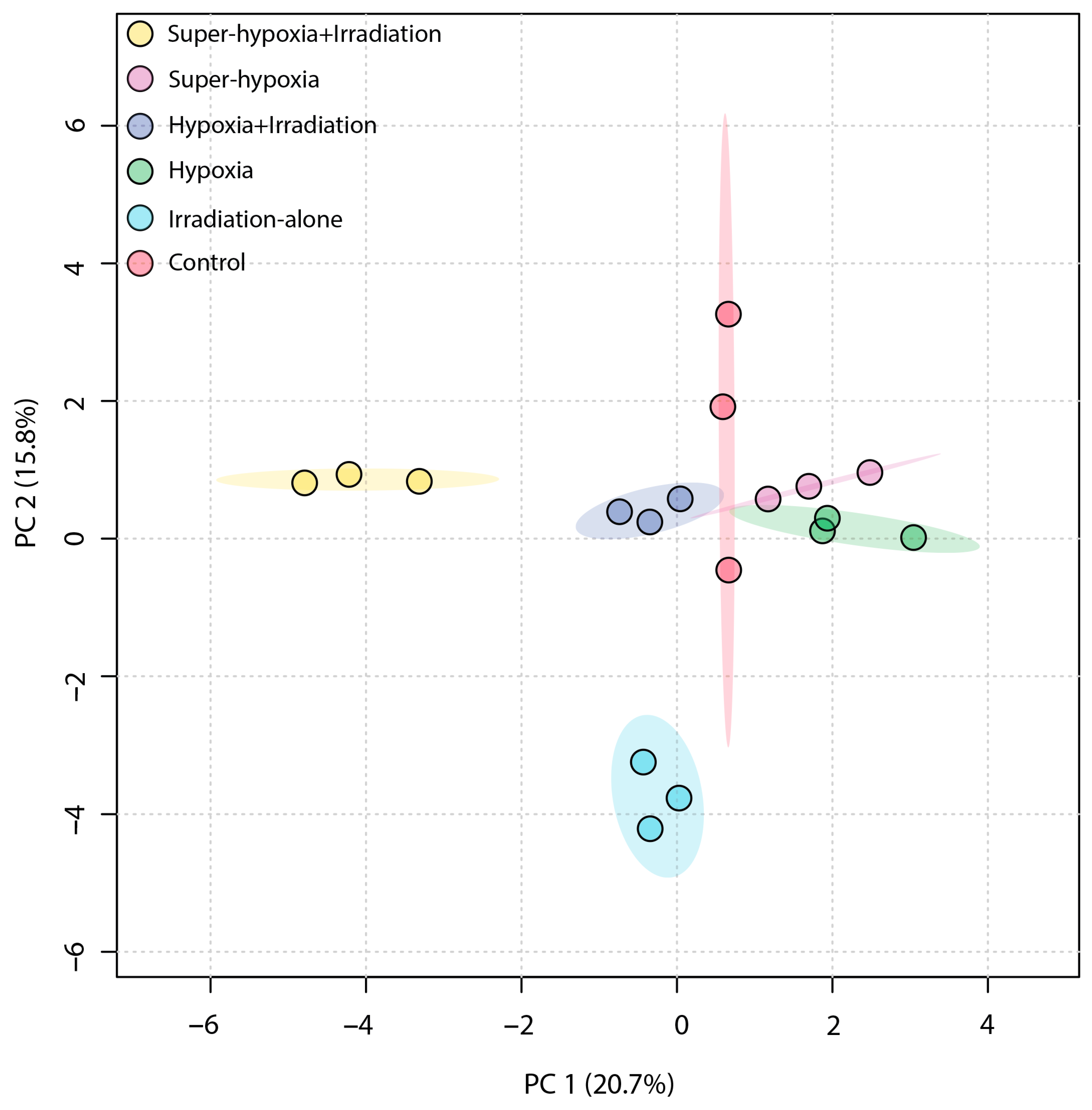
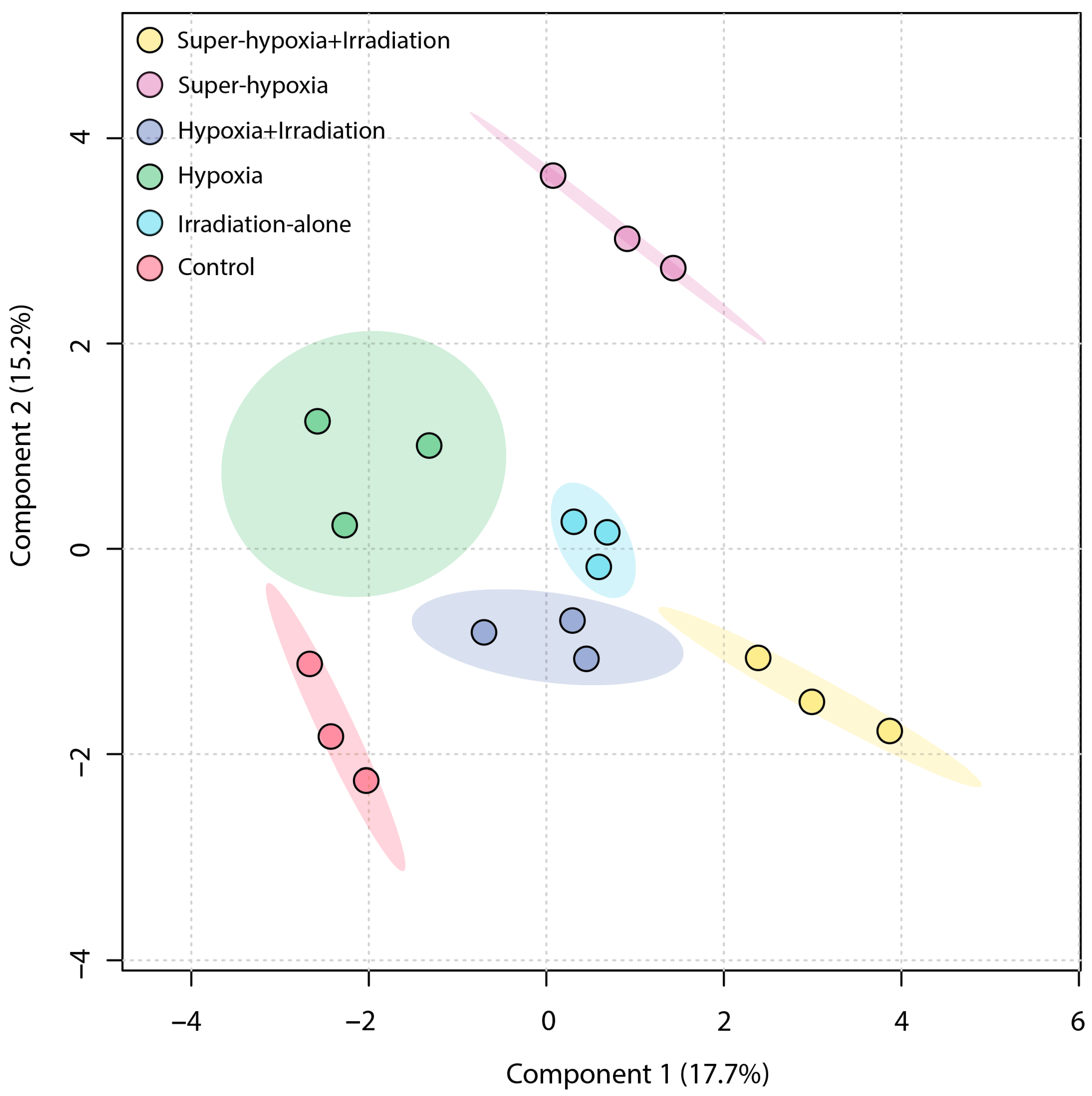
3.3. Multivariate Statistical Analysis of B. dorsalis Larvae Metabolites
3.4. Statistical Analysis and Differentially Regulated Metabolites
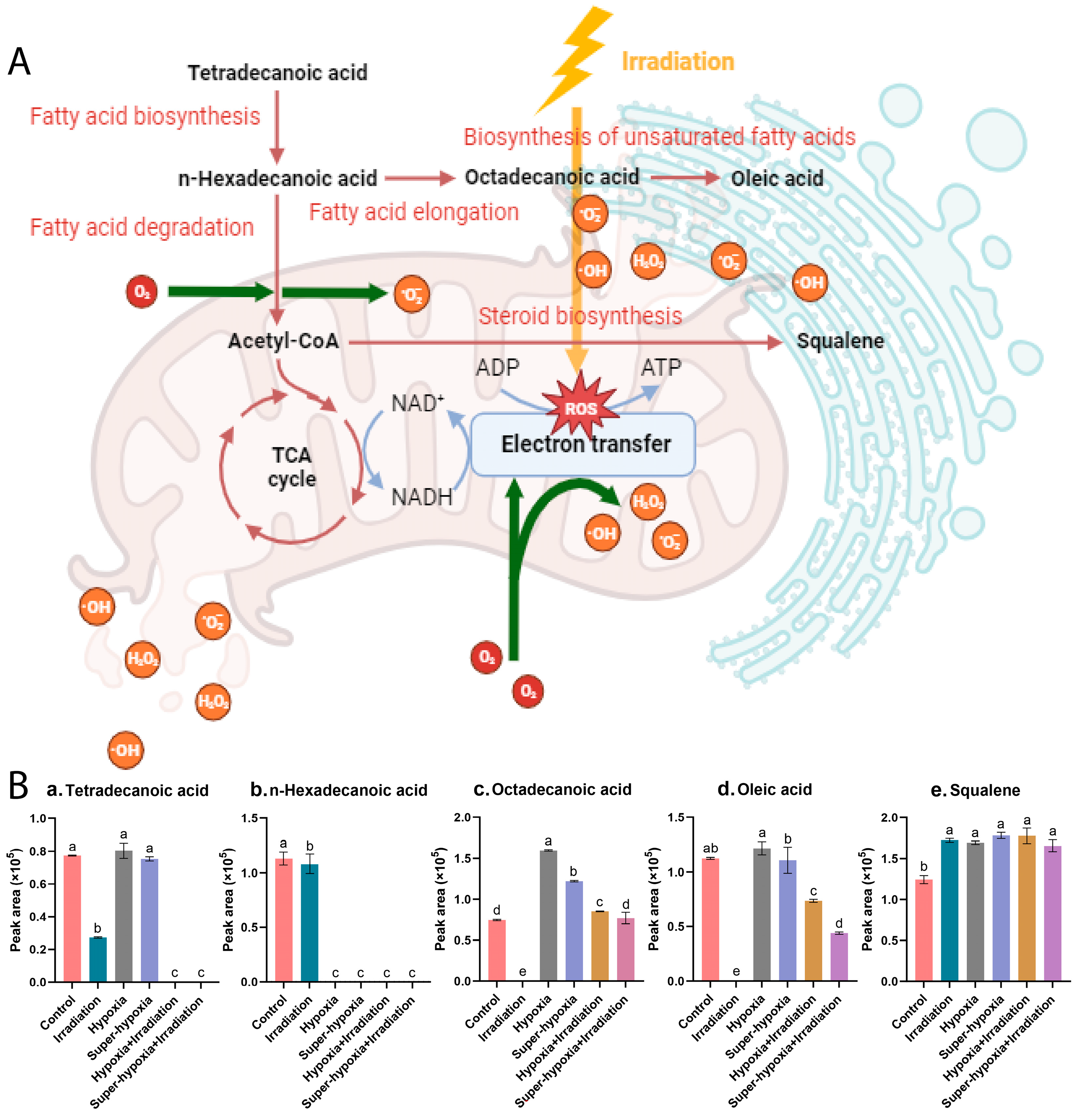
3.5. Key Metabolic Pathway Analysis
3.6. Comparison of the Main Differential Metabolic Responses
4. Discussion
5. Conclusions
Supplementary Materials
 represented compounds that were selected based on a significant p-value threshold (<0.05), while the
represented compounds that were selected based on a significant p-value threshold (<0.05), while the  indicated non-significant compounds.
indicated non-significant compounds.Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Clarke, A.R.; Li, Z.H.; Qin, Y.J.; Zhao, Z.H.; Liu, L.J.; Schutze, M.K. Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) is not invasive through Asia: It’s been there all along. J. Appl. Entomol. 2019, 143, 797–801. [Google Scholar] [CrossRef]
- DeMeyer, M.; Ekesi, S. Exotic invasive fruit flies (Diptera: Tephritidae): In and out of Africa. In Fruit Fly Research and Development in Africa—Towards a Sustainable Management Strategy to Improve Horticulture; Springer: Cham, Switzerland, 2016; pp. 127–150. [Google Scholar]
- Clarke, A.R.; Measham, P.F. Competition: A Missing component of fruit fly (Diptera: Tephritidae) risk assessment and planning. Insects 2022, 13, 1065. [Google Scholar] [CrossRef]
- Ihsanullah, I.; Rashid, A. Current activities in food irradiation as a sanitary and phytosanitary treatment in the Asia and the Pacific Region and a comparison with advanced countries. Food Control 2017, 72, 345–359. [Google Scholar] [CrossRef]
- Follett, P.A. Phytosanitary irradiation for fresh horticultural commodities: Generic treatments, current issues, and next steps. Stewart Postharvest Rev. 2014, 3, 2014. [Google Scholar]
- USDA; APHIS. Treatments for fruits and vegetables. Rules and Regulations. Fed. Regist. 2006, 71, 4451–4464. [Google Scholar]
- Rashvand, M.; Matera, A.; Altieri, G.; Genovese, F.; Fadiji, T.; Opara, U.L.; Di Renzo, G.C. Recent advances in the potential of modeling and simulation to assess the performance of modified atmosphere packaging (MAP) systems for the fresh agricultural product: Challenges and development. Trends Food Sci. Technol. 2023, 136, 48–63. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, W.; Meng, J.; Speakmon, M.; Qiu, J.; Pillai, S.; Zhu-Salzman, K. Hypoxic environment protects cowpea bruchid (Callosobruchus maculatus) from electron beam irradiation damage. Pest Manag. Sci. 2019, 75, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Follett, P.A.; Swedman, A.; Mackey, B. Effect of low-oxygen conditions created by modified atmosphere packaging on radiation tolerance in Drosophila suzukii (Diptera: Drosophilidae) in sweet cherries. J. Econ. Entomol. 2018, 111, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.S.; Hallman, G.J.; Martínez-Barrera, O.Y.; Hurtado, N.V.; Cardoso, A.A.; Parker, A.G.; Myers, S.W. Modified atmosphere does not reduce the efficacy of phytosanitary irradiation doses recommended for tephritid fruit flies. Insects 2020, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Follett, P.A.; Neven, L.G. Phytosanitary irradiation: Does modified atmosphere packaging or controlled atmosphere storage creating a low oxygen environment threaten treatment efficacy. Radiat. Phys. Chem. 2020, 173, 108874. [Google Scholar] [CrossRef]
- Hallman, G.J. Explanatory Document on International Standard for Phytosanitary Measures No. 18: Guidelines for the Use of Irradiation as a Phytosanitary Treatment: ISPM No. 18/Explanatory Document; International Plant Protection Convention (IPPC): Rome, Italy, 2006. [Google Scholar]
- Alexovič, M.; Andruch, V.; Balogh, I.S.; Šandrejová, J. A single-valve sequential injection manifold (SV-SIA) for automation of air-assisted liquid-phase microextraction: Stopped flow spectrophotometric determination of chromium (VI). Anal. Methods 2013, 5, 2497–2502. [Google Scholar] [CrossRef]
- Alexovič, M.; Horstkotte, B.; Solich, P. Automation of static and dynamic non-dispersive liquid phase microextraction. Part 1: Approaches based on extractant drop-, plug-, film-and microflow-formation. Anal. Chim. Acta 2016, 906, 22–40. [Google Scholar] [CrossRef]
- Gallo, M.; Ferranti, P. The evolution of analytical chemistry methods in foodomics. J. Chromatogr. A 2016, 1428, 3–15. [Google Scholar] [CrossRef]
- Zheng, J.; Kuang, Y.; Zhou, S.; Gong, X.; Ouyang, G. Latest improvements and expanding applications of solid-phase microextraction. Anal. Chem. 2023, 95, 218–237. [Google Scholar] [CrossRef]
- Shan, C.; Li, B.; Li, L.; Li, B.; Ren, Y.; Liu, T. Correlation between irradiation treatment and metabolite changes in Bactrocera dorsalis (Diptera: Tephritidae) Larvae using solid-phase microextraction (SPME) coupled with gas chromatography-mass spectrometry (GC-MS). Molecules 2022, 27, 4641. [Google Scholar] [CrossRef] [PubMed]
- Alnajim, I.; Agarwal, M.; Liu, T.; Li, B.; Du, X.; Ren, Y. Preliminary study on the differences in hydrocarbons between phosphine-susceptible and-resistant strains of Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbst) using direct immersion solid-phase microextraction coupled with GC-MS. Molecules 2020, 25, 1565. [Google Scholar] [CrossRef] [PubMed]
- Vargas, R.I.; Miyashita, D.; Nishida, T. Life history and demographic parameters of three laboratory-reared tephritids (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 1984, 77, 651–656. [Google Scholar] [CrossRef]
- Shan, C.; Li, B.; Li, L.; Du, X.; Ren, Y.; McKirdy, S.J.; Liu, T. Comparison of fumigation efficacy of methyl bromide alone and phosphine applied either alone or simultaneously or sequentially against Bactrocera correcta in Selenicereus undatus (red pitaya) fruit. Pest Manag. Sci. 2023, 79, 4942–4951. [Google Scholar] [CrossRef]
- Zhao, J.; Ma, J.; Wu, M.; Jiao, X.; Wang, Z.; Liang, F.; Zhan, G. Gamma radiation as a phytosanitary treatment against larvae and pupae of Bactrocera dorsalis (Diptera: Tephritidae) in guava fruits. Food Control 2017, 72, 360–366. [Google Scholar] [CrossRef]
- Cao, K.; Lan, R.; Yang, X.; Gong, B.; Zhang, J.; Zhou, X.; Jin, L. Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae). Agriculture 2023, 13, 1254. [Google Scholar] [CrossRef]
- Pan, K.; Xin, T.; Chen, Y.; Wang, H.; Wen, K.; Liu, Y.; Xia, B. Age-Stage, Two-Sex Life Table and Functional Response of Amblyseius orientalis (Acari: Phytoseiidae) Feeding on Different Nutrient Sources. Insects 2022, 13, 983. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shan, C.; Liu, Q.; Li, B.; Liu, T. Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry. Molecules 2023, 28, 7721. [Google Scholar] [CrossRef]
- Hou, J.; Han, Z.P.; Jing, Y.Y.; Yang, X.; Zhang, S.S.; Sun, K.; Wei, L.X. Autophagy prevents irradiation injury and maintains stemness through decreasing ROS generation in mesenchymal stem cells. Cell Death Dis. 2013, 4, 844. [Google Scholar] [CrossRef] [PubMed]
- Arvold, N.D.; Guha, N.; Wang, D.; Matli, M.; Deen, D.F.; Warren, R.S.; Haas-Kogan, D.A. Hypoxia-induced radioresistance is independent of hypoxia-inducible factor-1A in vitro. Int. J. Radiat. Oncol. Biol. Phys. 2005, 62, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.H.S.; Surendraraj, A.; Anandan, R. Protective effect of squalene on mitochondrial alterations in isoprenaline-induced myocardial injury. Int. J. Biomed. Pharmacol. Sci. 2010, 4, 32–37. [Google Scholar]
- Hallman, G.J.; Hénon, Y.M.; Parker, A.G.; Blackburn, C.M. Phytosanitary irradiation: An overview. Fla. Entomol. 2016, 99, 1–13. [Google Scholar]
- USDA Treatment Manual. Available online: https://www.aphis.usda.gov/import_export/plants/manuals/ports/downloads/treatment.pdf (accessed on 1 May 2020).
- Ali, H.M.; Moustafa, H.Z.; Sayed, R.M. Alterations in Biomarkers Associated with Sterility in Pectinophora gossypiella (Saunders) Induced by Gamma Irradiation. Braz. Arch. Biol. Technol. 2018, 60, e17160634. [Google Scholar] [CrossRef]
- Vimal, N.; Angmo, N.; Sengupta, M.; Seth, R.K. Radiation Hormesis to Improve the Quality of Adult Spodoptera litura (Fabr.). Insects 2022, 13, 933. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Boguś, M. The metabolism and role of free fatty acids in key physiological processes in insects of medical, veterinary and forensic importance. PeerJ 2021, 9, e12563. [Google Scholar] [CrossRef]
- Zhu, H.; Han, M. Exploring developmental and physiological functions of fatty acid and lipid variants through worm and fly genetics. Annu. Rev. Genet. 2014, 48, 119–148. [Google Scholar] [CrossRef]
- Rawat, N.; Singla-Pareek, S.L.; Pareek, A. Membrane dynamics during individual and combined abiotic stresses in plants and tools to study the same. Physiol. Plant. 2021, 171, 653–676. [Google Scholar] [CrossRef]
- Inayat, M.; Abbas, F.; Rehman, M.H.; Mahmud, A. Physico-chemical parameters, oxidative stress, and fatty acid profile of American Pekin ducks (Anas platyrhynchos domesticus) raised under different production systems. Braz. J. Poult. Sci. 2023, 25, 1–8. [Google Scholar] [CrossRef]
- Qiu, K.; Zhao, Q.; Wang, J.; Qi, G.H.; Wu, S.G.; Zhang, H.J. Effects of pyrroloquinoline quinone on lipid metabolism and anti-oxidative capacity in a high-fat-diet metabolic dysfunction-associated fatty liver disease chick model. Int. J. Mol. Sci. 2021, 22, 1458. [Google Scholar] [CrossRef] [PubMed]
- El-Maradny, A. Identification of Fatty Acid Methyl Esters, Long Chain Aldehydes and Free Fatty Acids in a Sponge Chalinula saudensis along the coast of Jeddah, Saudi Arabia. J. Chem. Soc. Pak. 2012, 34, 673–678. [Google Scholar]
- Liu, Y.; Liu, J.; Deng, C.; Zhang, X. Graphene and graphene oxide: Two ideal choices for the enrichment and ionization of long-chain fatty acids free from matrix-assisted laser desorption/ionization matrix interference. Rapid Commun. Mass Spectrom. 2011, 25, 3223–3234. [Google Scholar] [CrossRef]
- Senbagalakshmi, P.; Muthukrishnan, S.; Jebasingh, T.; Kumar, T.S.; Rao, M.; Kumar, T.S.; Rao, M.V. Squalene, Biosynthesis and its role in production of bioactive compounds, a Proper Scientific Challenge—A Review. J. Emerg. Technol. Innov. Res. 2019, 6, 505–526. [Google Scholar]
- Atamna, H.; Mackey, J.; Dhahbi, J.M. Mitochondrial pharmacology: Electron transport chain bypass as strategies to treat mitochondrial dysfunction. Biofactors 2012, 38, 158–166. [Google Scholar] [CrossRef]
- Aretz, I.; Hardt, C.; Wittig, I.; Meierhofer, D. An impaired respiratory electron chain triggers down-regulation of the energy metabolism and de-ubiquitination of solute carrier amino acid transporters. Mol. Cell. Proteom. 2016, 15, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kitazawa, H.; Wang, X.; Sun, H. Regulation of respiratory pathway and electron transport chain in relation to senescence of postharvest white mushroom (Agaricus bisporus) under high O2/CO2 controlled atmospheres. J. Agric. Food Chem. 2017, 65, 3351–3359. [Google Scholar] [CrossRef]
- Bochicchio, P.A.; Pérez, M.M.; Quesada-Allué, L.A.; Rabossi, A. Completion of metamorphosis after adult emergence in Ceratitis capitata (Diptera: Tephritidae). Curr. Res. Insect Sci. 2021, 1, 100017. [Google Scholar] [CrossRef]
- Song, Y.; Gu, F.; Liu, Z.; Li, Z.; Wu, F.A.; Sheng, S. The key role of fatty acid synthase in lipid metabolism and metamorphic development in a destructive insect pest, Spodoptera litura (Lepidoptera: Noctuidae). Int. J. Mol. Sci. 2022, 23, 9064. [Google Scholar] [CrossRef]
- Nathaniel, T.I.; Otukonyong, E.; Abdellatif, A.; Soyinka, J.O. Effect of hypoxia on metabolic rate, core body temperature, and c-fos expression in the naked mole rat. Int. J. Dev. Neurosci. 2012, 30, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Scisciola, L.; Chianese, U.; Caponigro, V.; Basilicata, M.G.; Salviati, E.; Altucci, L.; Sommella, E. Multi-omics analysis reveals attenuation of cellular stress by empagliflozin in high glucose-treated human cardiomyocytes. J. Transl. Med. 2023, 21, 662. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.E.; Bucarey, J.L.; Espinosa, A. Muscle lipid metabolism: Role of lipid droplets and perilipins. J. Diabetes Res. 2017, 2017, 1789395. [Google Scholar] [CrossRef] [PubMed]
- Welte, M.A. Expanding roles for lipid droplets. Curr. Biol. 2015, 25, 470–481. [Google Scholar] [CrossRef]
- Clanton, T.L. Hypoxia-induced reactive oxygen species formation in skeletal muscle. J. Appl. Physiol. 2007, 102, 2379–2388. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef]
- Szumiel, I. Ionizing radiation-induced oxidative stress, epigenetic changes and genomic instability: The pivotal role of mitochondria. Int. J. Radiat. Biol. 2015, 91, 1–12. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol. 2008, 51, 167–173. [Google Scholar] [CrossRef]
- Naeini, S.H.; Mavaddatiyan, L.; Kalkhoran, Z.R.; Taherkhani, S.; Talkhabi, M. Alpha-ketoglutarate as a potent regulator for lifespan and healthspan: Evidences and perspectives. Exp. Gerontol. 2023, 175, 112154. [Google Scholar] [CrossRef]
- Lv, C.; Ma, X.; Liang, C.; Chen, Y.; Qin, F.; Zhou, C.; Hou, L. The interaction of pterostilbene with Kelch-like ECH-associated protein 1 and its regulation on the nuclear factor erythroid 2-related factor 2/antioxidant response element pathway. Process Biochem. 2023, 132, 228–235. [Google Scholar] [CrossRef]
- Pan, C.; Ai, C.; Liang, L.; Zhang, B.; Li, Q.; Pu, L.; Wang, X. Sestrin2 protects against hypoxic nerve injury by regulating mitophagy through SESN2/AMPK pathway. Front. Mol. Biosci. 2023, 10, 1266243. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Liu, R.; Zhou, Y.; Zhang, Z.; Jiang, X.; Xu, J.; Hu, Z. FOXO3a-dependent up-regulation of HSP90 alleviates cisplatin-induced apoptosis by activating FUNDC1-mediated mitophagy in hypoxic osteosarcoma cells. Cell. Signal. 2023, 101, 110500. [Google Scholar] [CrossRef] [PubMed]
- Olazcuaga, L.; Baltenweck, R.; Leménager, N.; Maia-Grondard, A.; Claudel, P.; Hugueney, P.; Foucaud, J. Metabolic consequences of various fruit-based diets in a generalist insect species. eLife 2023, 12, e84370. [Google Scholar] [CrossRef] [PubMed]
- Bayer, R.K.; Cagiao, M.E.; Calleja, F.B. Structure development in amorphous starch as revealed by X-ray scattering: Influence of the network structure and water content. J. Appl. Polym. Sci. 2006, 99, 1880–1886. [Google Scholar] [CrossRef]
- Jacobs, H.; Mischenko, N.; Koch, M.H.; Eerlingen, R.C.; Delcour, J.A.; Reynaers, H. Evaluation of the impact of annealing on gelatinisation at intermediate water content of wheat and potato starches: A differential scanning calorimetry and small angle X-ray scattering study. Carbohydr. Res. 1998, 306, 1–10. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological effects of physical postharvest treatments on insects. HortTechnology 2003, 13, 272–275. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Conditions | Control | Irradiation |
|---|---|---|---|
| Larvae | Normoxia | 3.09 ± 1.25 bA | 3.55 ± 1.52 bB |
| Hypoxia (5%) | 4.00 ± 1.51 aA | 4.02 ± 1.74 abA | |
| Sup-Hypoxia (0.50%) | 4.05 ± 2.07 aA | 4.23 ± 1.94 aA | |
| Pupa | Normoxia | 5.48 ± 0.70 c | N.A. |
| Hypoxia (5%) | 5.89 ± 0.51 b | N.A. | |
| Sup-Hypoxia (0.50%) | 6.52 ± 0.78 a | N.A. | |
| Adult | Normoxia | N.A. | N |
| Hypoxia (5%) | N.A. | N | |
| Sup-Hypoxia (0.50%) | N.A. | N |
| 1 RT | 2 RI | Metabolites | Peak Area ± 4 SD | p-Value | CAS | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Normoxia | Hypoxia | Hypoxia + Irradiation (116 Gy) | ||||||||
| Control | Irradiation-Alone | 95% N2 | 99.5% N2 | 95% N2 | 99.5% N2 | |||||
| 12.130 | 1141.22 | (E)-2-Decenal | 0.255 ± 0.013 a | 3 N.D. c | 0.212 ± 0.010 b | N.D. c | N.D. c | N.D. c | 1.75 × 10−13 | 003913-81-3 |
| 13.206 | 1256.58 | (E,E)-2,4-Decadienal | N.D. c | N.D. c | 0.241 ± 0.041 a | 0.221 ± 0.020 ab | 0.240 ± 0.026 a | 0.237 ± 0.014 b | 7.27 × 10−14 | 025152-84-5 |
| 14.093 | 1322.34 | 2-Dodecenal | 0.397 ± 0.068 a | N.D. b | 0.348 ± 0.013 a | N.D. b | N.D. b | N.D. b | 6.04 × 10−11 | 4826-62-4 |
| 14.726 | 1400.00 | Tetradecane | 0.559 ± 0.002 abc | 0.533 ± 0.007 bc | 0.560 ± 0.021 ab | 0.560 ± 0.002 abc | 0.574 ± 0.240 a | 0.564 ± 0.077 c | 7.83 × 10−2 | 000629-59-4 |
| 14.909 | 1419.12 | Tetradecanal | 0.660 ± 0.005 a | 0.359 ± 0.001 b | N.D. c | N.D. c | N.D. c | N.D. c | 4.39 × 10−10 | 000124-25-4 |
| 16.383 | 1509.89 | 1-Hexadecanol | 0.961 ± 0.004 a | N.D. c | 0.935 ± 0.002 a | 0.673 ± 0.001 b | N.D. c | N.D. c | 8.43 × 10−5 | 36653-82-4 |
| 16.719 | 1540.38 | 4-methyl-Heptadecane | 0.659 ± 0.036 a | N.D. b | N.D. b | N.D. b | N.D. b | N.D. b | 3.28 × 10−8 | 26429-11-8 |
| 18.204 | 1600.00 | Hexadecane | 0.761 ± 0.042 b | 0.760 ± 0.030 b | 0.789 ± 0.033 b | 0.788 ± 0.032 b | 0.902 ± 0.199 a | 0.968 ± 0.021 a | 2.31 × 10−5 | 000544-76-3 |
| 19.123 | 1634.95 | 2-Pentadecanone | 0.840 ± 0.100 a | N.D. b | N.D. b | N.D. b | N.D. b | N.D. b | 1.34 × 10−6 | 2345-28-0 |
| 20.741 | 1642.73 | Tetradecanoic acid | 0.773 ± 0.001 a | 0.272 ± 0.007 b | 0.807 ± 0.057 a | 0.757 ± 0.011 a | N.D. c | N.D. c | 8.39 × 10−9 | 000544-63-8 |
| 21.334 | 1700.00 | Heptadecane | 1.229 ± 0.066 a | 1.104 ± 0.515 ab | 1.065 ± 0.057 bc | 0.942 ± 0.198 c | 1.043 ± 0.210 bc | 1.153 ± 0.081 ab | 7.26 × 10−3 | 000629-78-7 |
| 21.413 | 1900.00 | Nonadecane | 0.461 ± 0.014 a | N.D. b | N.D. b | N.D. b | N.D. b | N.D. b | 1.01 × 10−9 | 629-92-5 |
| 22.268 | 1942.36 | 1-Octadecene | 1.591 ± 0.046 a | 1.263 ± 0.263 b | 0.775 ± 0.148 c | 0.035 ± 0.001 e | 0.645 ± 0.102 d | N.D. e | 1.97 × 10−11 | 112-88-9 |
| 22.865 | 1984.96 | 2-Nonadecanone | 0.949 ± 0.002 bc | 0.211 ± 0.27 d | 1.015 ± 0.104 ab | 1.042 ± 0.131 abc | 0.913 ± 0.004 c | 1.135 ± 0.090 a | 6.70 × 10−8 | 000629-66-3 |
| 23.470 | 2026.34 | n-Hexadecanoic acid | 1.131 ± 0.159 a | 1.085 ± 0.184 b | N.D. c | N.D. c | N.D. c | N.D. c | 8.56 × 10−8 | 000112-62-9 |
| 23.811 | 2088.36 | Hexadecenoic acid | 1.067 ± 0.018 a | N.D. e | 0.960 ± 0.006 ab | 0.962 ± 0.007 b | 0.748 ± 0.024 c | 0.162 ± 0.069 d | 1.11 × 10−13 | 000057-10-3 |
| 24.300 | 2144.53 | Hexadecanoic acid, ethyl ester | N.D. c | N.D. c | 1.043 ± 0.020 b | 1.135 ± 0.002 a | N.D. c | N.D. c | 3.36 × 10−6 | 000112-95-8 |
| 26.376 | 2177.57 | Oleic acid | 1.120 ± 0.013 ab | N.D. e | 1.150 ± 0.053 a | 1.099 ± 0.102 b | 0.731 ± 0.008 c | 0.439 ± 0.004 d | 2.40 × 10−9 | 000060-33-3 |
| 28.870 | 2228.92 | Octadecanoic acid | 0.745 ± 0.001 d | N.D. e | 1.599 ± 0.004 a | 1.224 ± 0.010 b | 0.855 ± 0.002 c | 0.771 ± 0.095 d | 1.10 × 10−10 | 9011-21-6 |
| 30.558 | 2262.71 | Cyclotetracosane | 1.279 ± 0.223 a | 1.239 ± 0.089 a | 1.068 ± 0.021 b | 0.935 ± 0.002 c | 0.545 ± 0.011 d | N.D. e | 3.44 × 10−11 | 000297-03-0 |
| 31.879 | 2304.94 | 11-Tricosene | 1.335 ± 0.071 a | 1.110 ± 0.012 b | 1.311 ± 0.081 a | 1.304 ± 0.072 a | 1.169 ± 0.002 b | 0.895 ± 0.107 c | 6.06 × 10−5 | 052078-56-5 |
| 33.506 | 2352.39 | 1-Docosene | 1.300 ± 0.051 a | N.D. b | N.D. b | N.D. b | N.D. b | N.D. b | 1.03 × 10−9 | 001599-67-3 |
| 34.741 | 2400.00 | Tetracosane | 1.539 ± 0.019 a | 1.096 ± 0.023 c | 1.410 ± 0.126 b | 1.560 ± 0.128 a | 0.690 ± 0.002 d | N.D. e | 3.09 × 10−11 | 000646-31-1 |
| 36.627 | 2687.78 | Squalene | 1.236 ± 0.007 b | 1.747 ± 0.008 a | 1.710 ± 0.020 a | 1.738 ± 0.091 a | 1.749 ± 0.017 a | 1.694 ± 0.012 a | 5.37 × 10−7 | 007683-64-9 |
| 42.658 | 2924.03 | Sitosterol | 1.756 ± 0.036 a | N.D. c | 0.629 ± 0.015 b | N.D. c | N.D. c | N.D. c | 3.42 × 10−11 | 000083-46-5 |
| Group | Pathway | Hits | Expect | p-Value | Holm P | FDR |
|---|---|---|---|---|---|---|
| CON vs. IR | Biosynthesis of unsaturated fatty acids | 3 | 0.117 | 0.000115 | 0.00963 | 0.00963 |
| Fatty acid biosynthesis | 2 | 0.153 | 0.00864 | 0.717 | 0.363 | |
| Fatty acid elongation | 1 | 0.124 | 0.118 | 1 | 1 | |
| Fatty acid degradation | 1 | 0.127 | 0.121 | 1 | 1 | |
| Steroid biosynthesis | 1 | 0.137 | 0.13 | 1 | 1 | |
| CON vs. HY | Biosynthesis of unsaturated fatty acids | 2 | 0.0938 | 0.00311 | 0.261 | 0.261 |
| Steroid biosynthesis | 1 | 0.109 | 0.105 | 1 | 1 | |
| Fatty acid biosynthesis | 1 | 0.122 | 0.117 | 1 | 1 | |
| CON vs. Sup-HY | Biosynthesis of unsaturated fatty acids | 1 | 0.0469 | 0.0463 | 1 | 1 |
| Fatty acid biosynthesis | 1 | 0.0612 | 0.0603 | 1 | 1 | |
| CON vs. HY + IR | Biosynthesis of unsaturated fatty acids | 3 | 0.141 | 0.000226 | 0.019 | 0.019 |
| Steroid biosynthesis | 2 | 0.164 | 0.0102 | 0.848 | 0.356 | |
| Fatty acid biosynthesis | 2 | 0.184 | 0.0127 | 1 | 0.356 | |
| Fatty acid elongation | 1 | 0.148 | 0.14 | 1 | 1 | |
| Fatty acid degradation | 1 | 0.152 | 0.143 | 1 | 1 | |
| CON vs. Sup-HY + IR | Biosynthesis of unsaturated fatty acids | 2 | 0.0703 | 0.00158 | 0.133 | 0.133 |
| Fatty acid elongation | 1 | 0.0742 | 0.0724 | 1 | 1 | |
| Fatty acid degradation | 1 | 0.0762 | 0.0743 | 1 | 1 | |
| Steroid biosynthesis | 1 | 0.0820 | 0.0799 | 1 | 1 | |
| Fatty acid biosynthesis | 1 | 0.0918 | 0.0891 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, C.; Li, B.; Li, L.; Liu, Q.; Zou, H.; Liu, T. Development and Metabolomic Profiles of Bactrocera dorsalis (Diptera: Tephritidae) Larvae Exposed to Phytosanitary Irradiation Dose in Hypoxic Environment Using DI-SPME-GC/MS. Insects 2024, 15, 177. https://doi.org/10.3390/insects15030177
Shan C, Li B, Li L, Liu Q, Zou H, Liu T. Development and Metabolomic Profiles of Bactrocera dorsalis (Diptera: Tephritidae) Larvae Exposed to Phytosanitary Irradiation Dose in Hypoxic Environment Using DI-SPME-GC/MS. Insects. 2024; 15(3):177. https://doi.org/10.3390/insects15030177
Chicago/Turabian StyleShan, Changyao, Baishu Li, Li Li, Qun Liu, Hang Zou, and Tao Liu. 2024. "Development and Metabolomic Profiles of Bactrocera dorsalis (Diptera: Tephritidae) Larvae Exposed to Phytosanitary Irradiation Dose in Hypoxic Environment Using DI-SPME-GC/MS" Insects 15, no. 3: 177. https://doi.org/10.3390/insects15030177
APA StyleShan, C., Li, B., Li, L., Liu, Q., Zou, H., & Liu, T. (2024). Development and Metabolomic Profiles of Bactrocera dorsalis (Diptera: Tephritidae) Larvae Exposed to Phytosanitary Irradiation Dose in Hypoxic Environment Using DI-SPME-GC/MS. Insects, 15(3), 177. https://doi.org/10.3390/insects15030177