



The Dynamics of Pheromone Release in Two Passive Dispensers Commonly Used for Mating Disruption in the Control of Lobesia botrana and Eupoecilia ambiguella in Vineyards



Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Pheromone Dispenser Types
2.2. Effect of Temperature on Pheromone Release
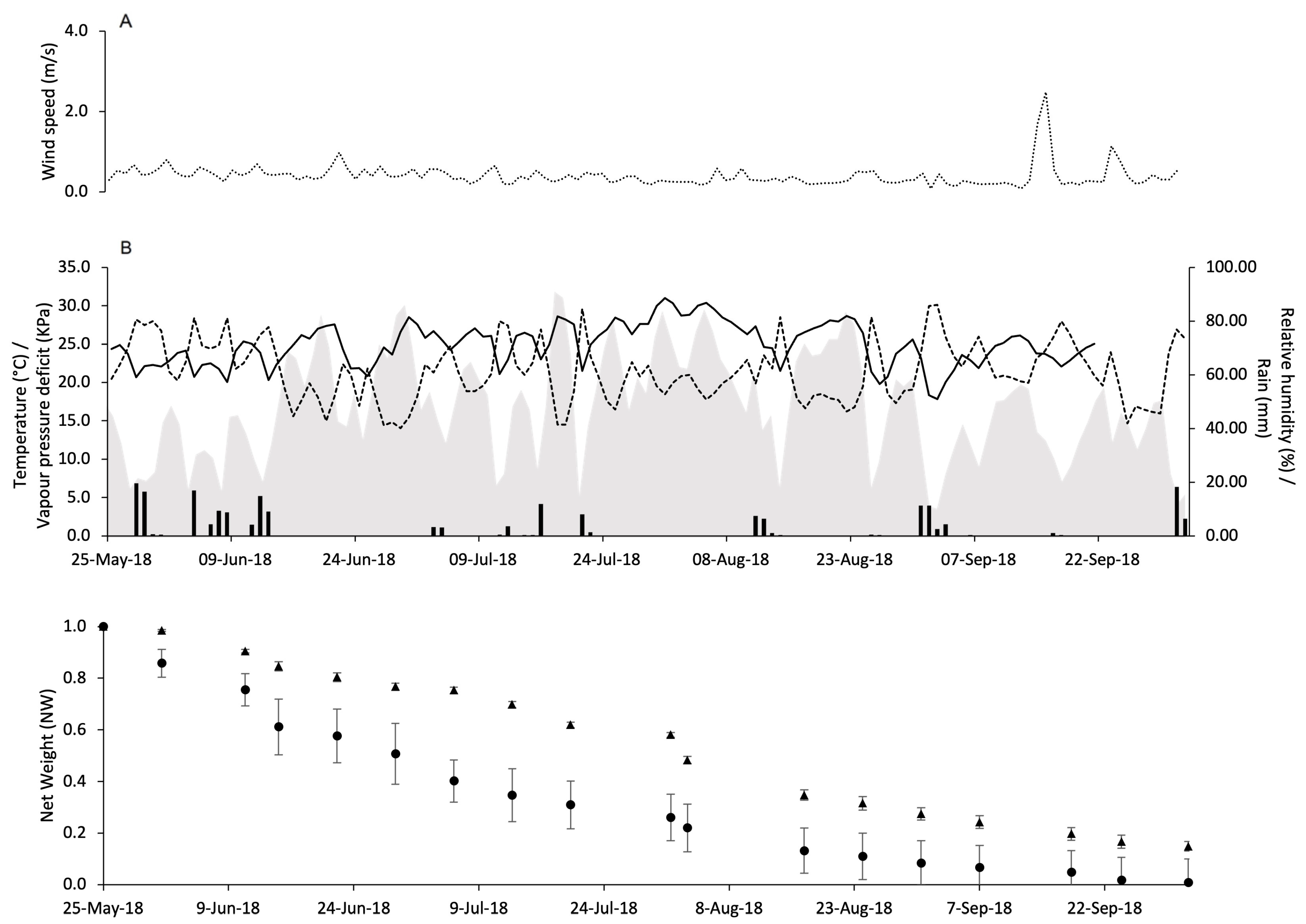
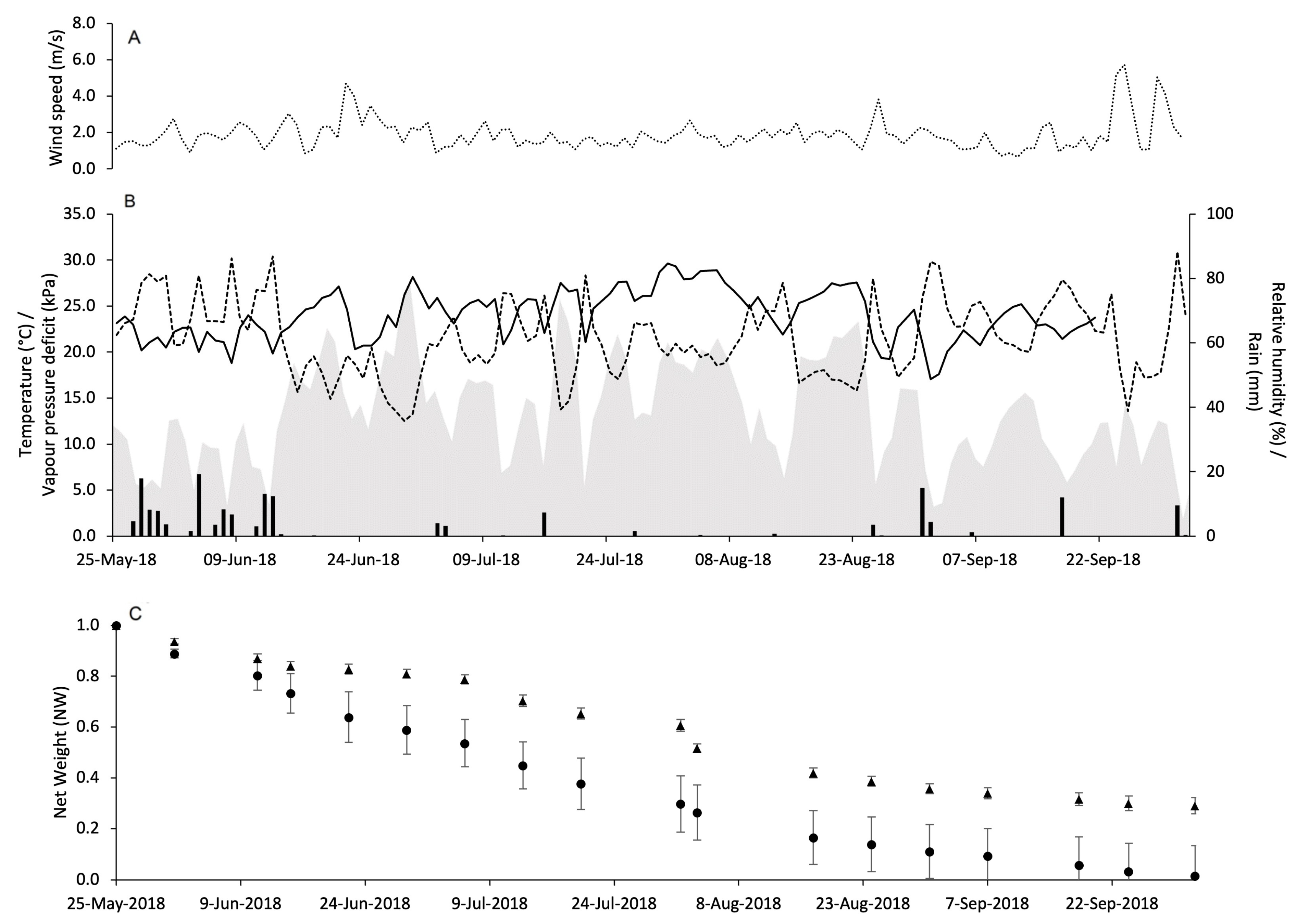
2.3. Pheromone Release Under Vineyard Conditions
3. Results
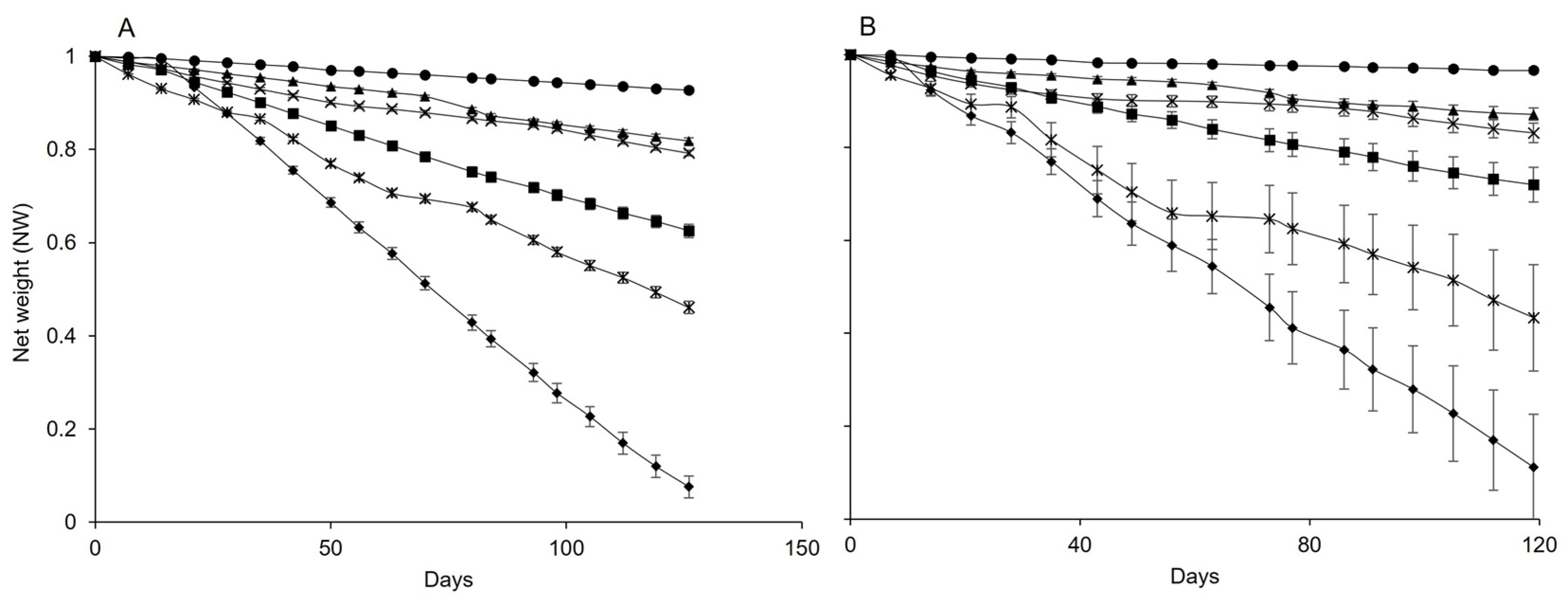
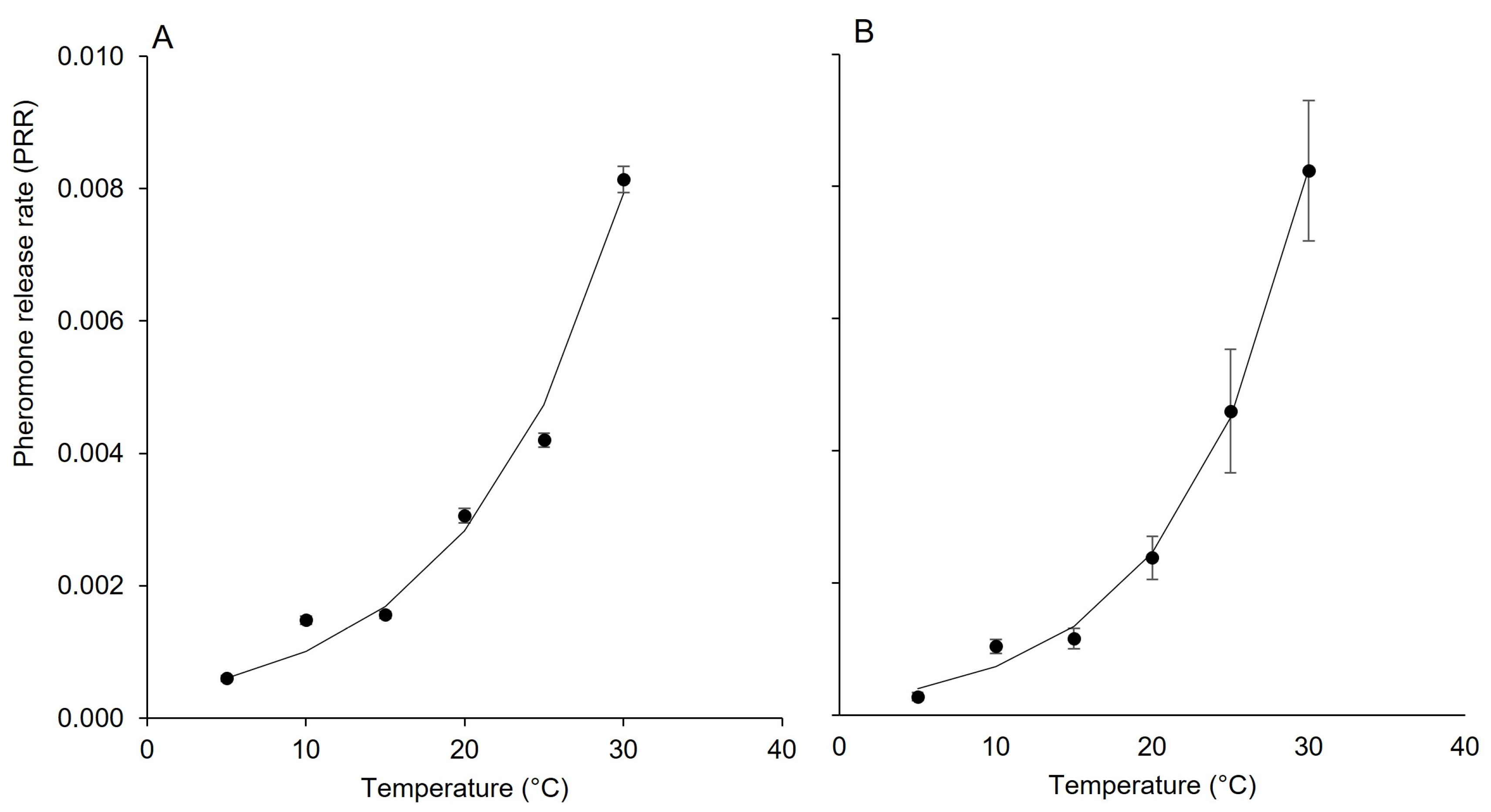
3.1. Effect of Temperature on Pheromone Release
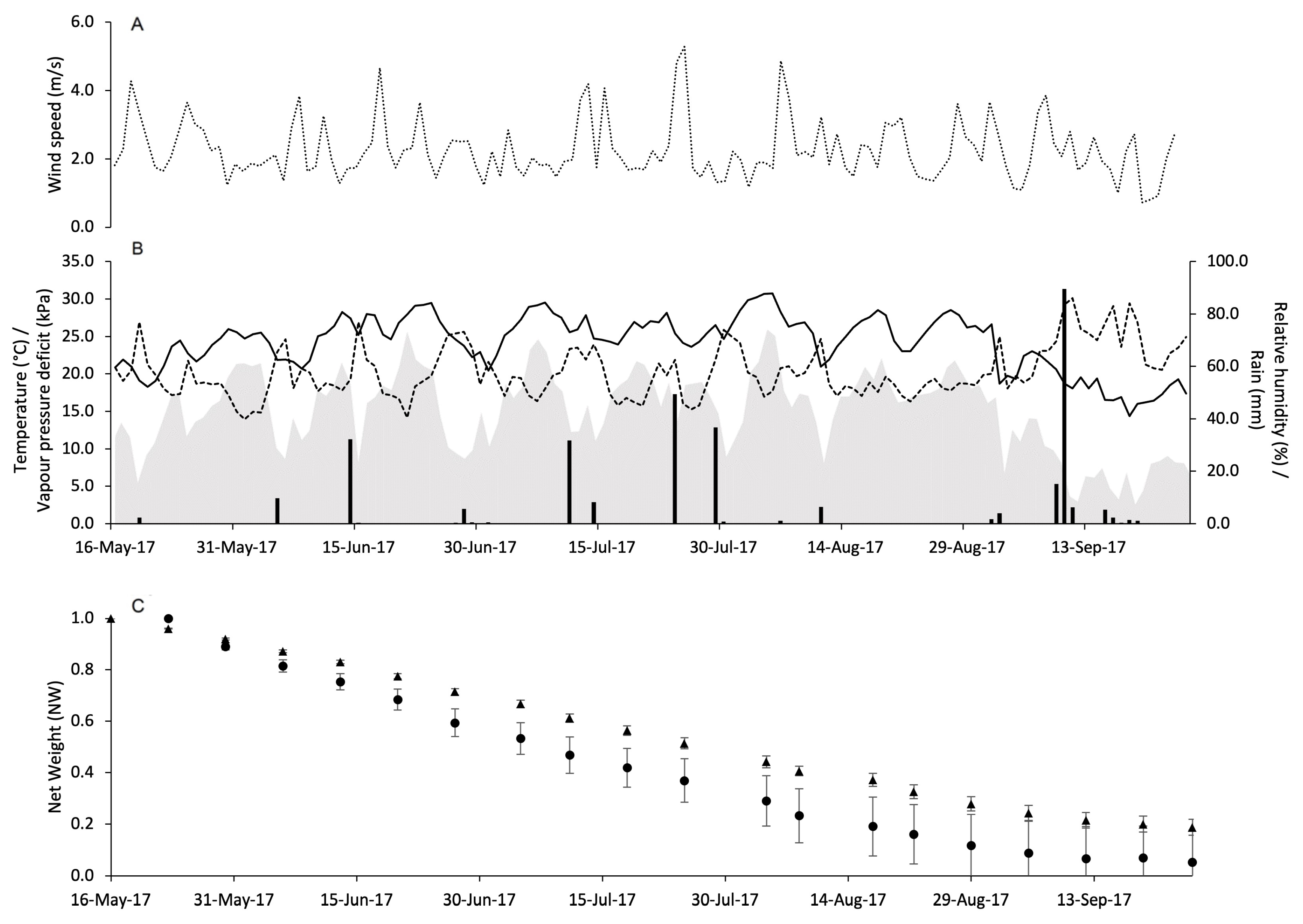
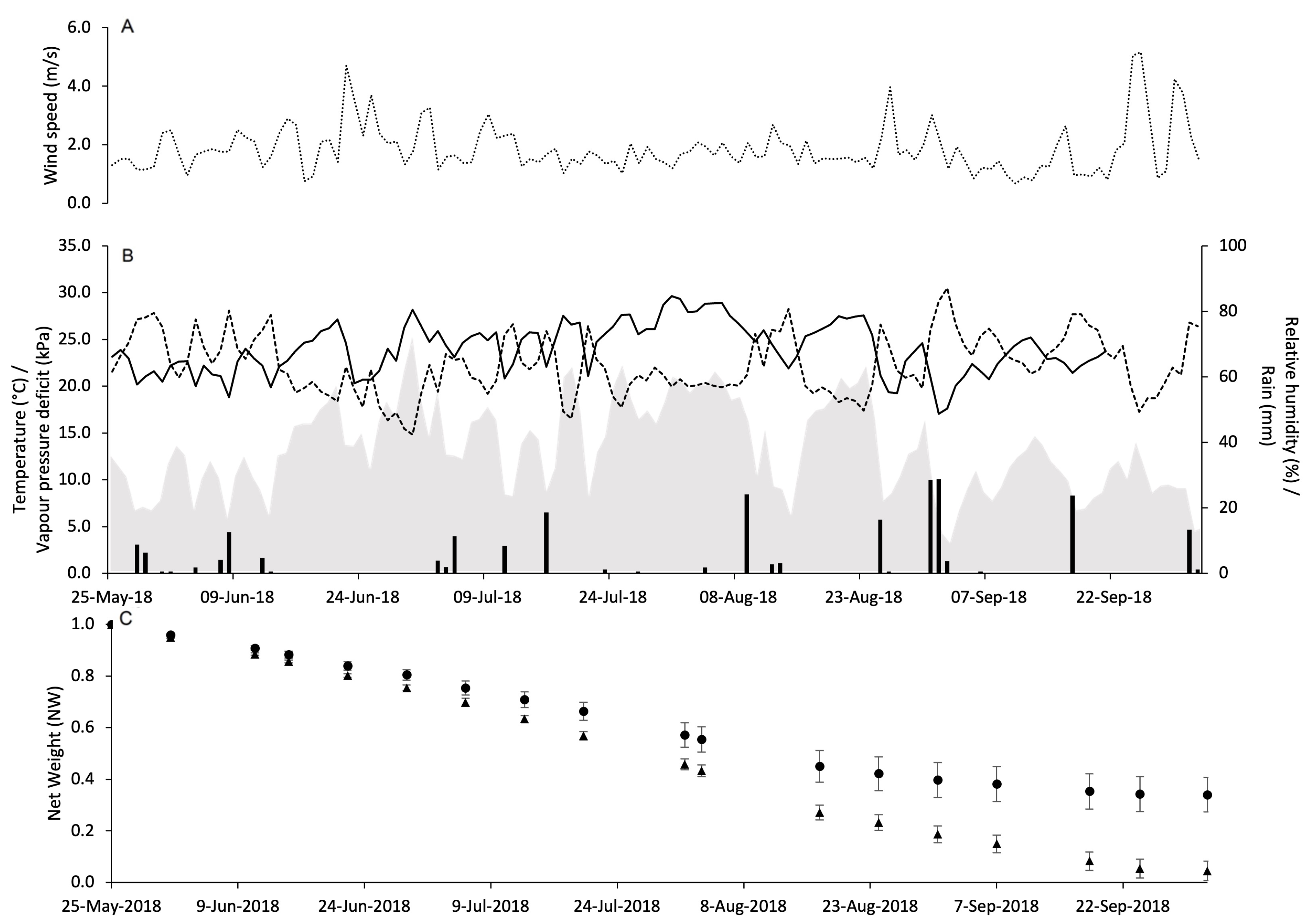
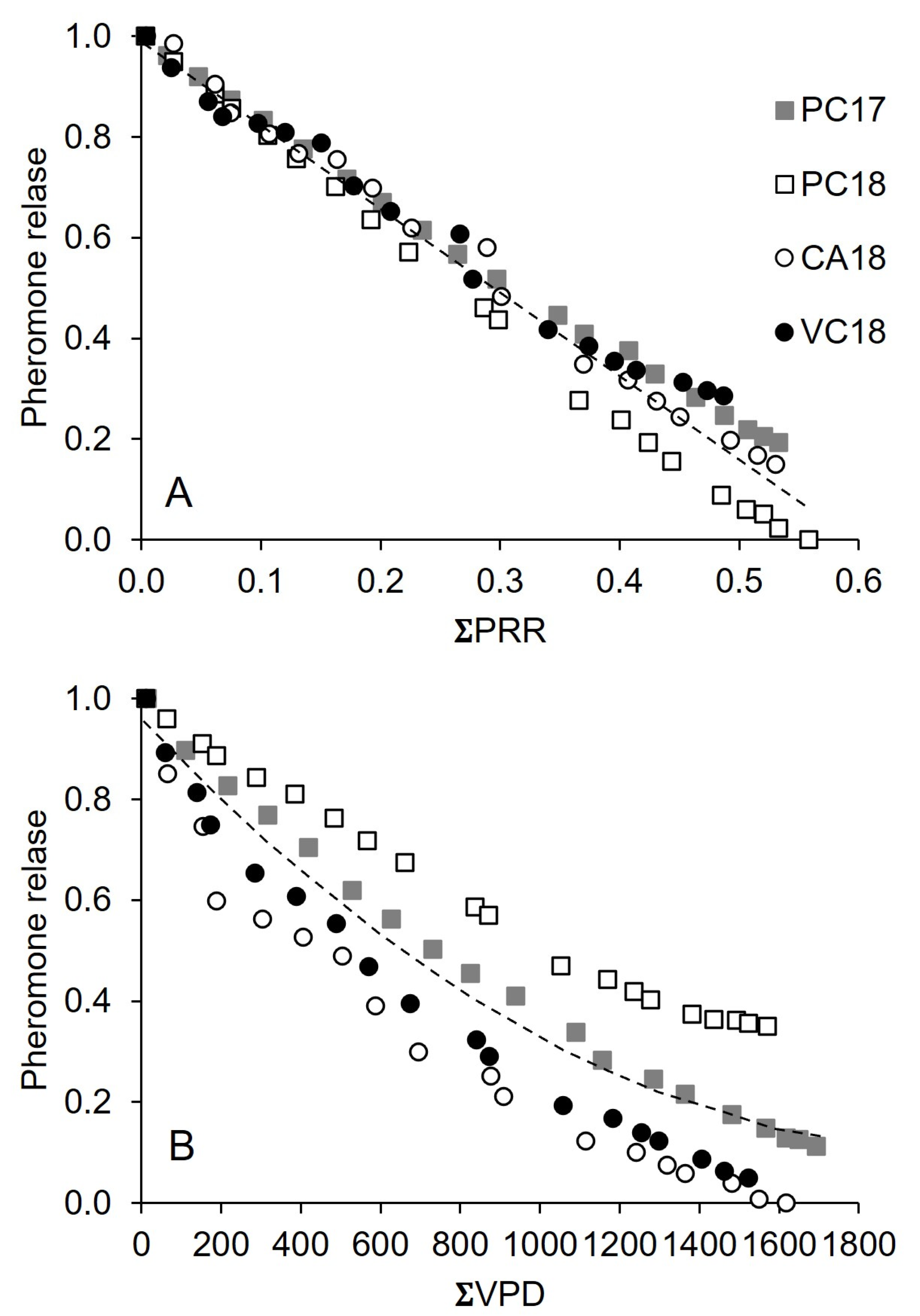
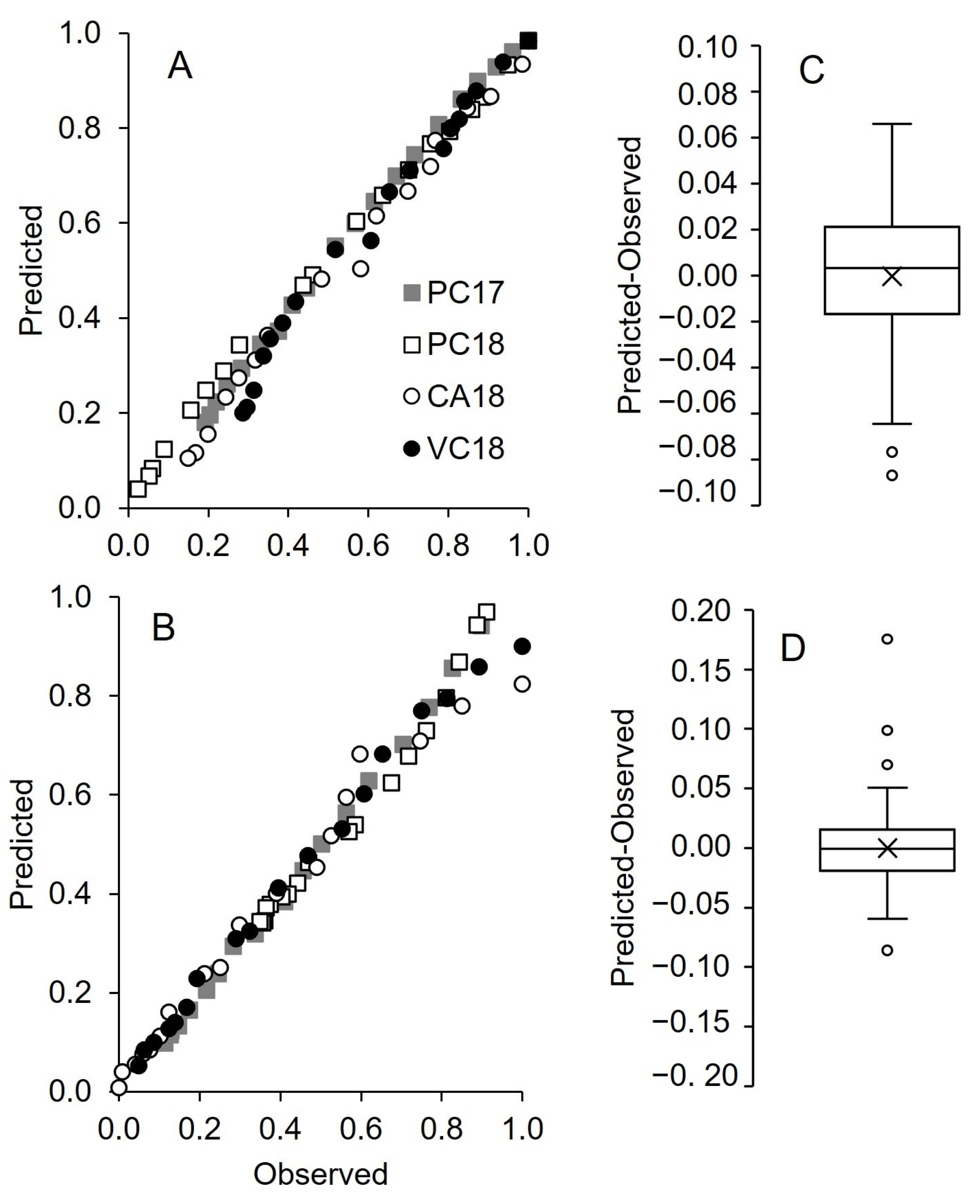
3.2. Pheromone Release Under Vineyard Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Louis, F.; Schirra, K.-J. Mating disruption of Lobesia botrana (Lepidoptera: Tortricidae) in vineyards with very high population densities. IOBC WPRS Bull. 2001, 24, 75–80. [Google Scholar]
- Lucchi, A.; Sambado, P.; Juan Royo, A.B.; Bagnoli, B.; Benelli, G. Lobesia botrana males mainly fly at dusk: Video camera-assisted pheromone traps and implications for mating disruption. J. Pest Sci. 2018, 91, 1327–1334. [Google Scholar] [CrossRef]
- Thiéry, D. Les Vers de la Grappe, les Connaître pour s’en Protéger; Vigne et Vins International: Bordeaux, France, 2005; p. 60. [Google Scholar]
- Varner, M.; Lucin, R.; Mattedi, L.; Forno, F. Experience with mating disruption technique to control grape berry moth, Lobesia botrana. IOBC WPRS Bull. 2001, 24, 81–88. [Google Scholar]
- Fermaund, M.; Le Menn, R. Association of Botrytis cinerea with grape berry moth larvae. Phytopathology 1989, 79, 651–656. [Google Scholar] [CrossRef]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entom. 2011, 104, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Vartholomaiou, A.N.; Navrozidis, E.I.; Payne, C.C.; Salpiggidis, G.A. Agronomic Techniques to Control Lobesia botrana. Phytoparasitica 2008, 36, 264–271. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A. Semiochemical Strategies for Tortricid Moth Control in Apple Orchards and Vineyards in Italy. J. Chem. Ecol. 2016, 42, 571–583. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Lorenzon, M.; Fornasiero, D.; Tirello, P.; Simoni, S.; Bagnoli, B. The Impact of Microbial and Botanical Inseticides on Grape Berry Moths and Their Effects on Secondary Pests and Beneficials. Agronomy 2022, 12, 217. [Google Scholar] [CrossRef]
- Shailaja, D.; Ahmed, S.M.; Yaseen, M. Comparative study of release kinetics of pheromone from polymer dispensers. J. Appl. Polym. Sci. 1997, 64, 1373–1380. [Google Scholar] [CrossRef]
- Gavara, A.; Vacas, S.; Navarro, I.; Primo, J.; Navarro-Llopis, V. Airborne pheromone quantification in treated vineyards with different mating disruption dispensers against lobesia botrana. Insects 2020, 11, 289. [Google Scholar] [CrossRef]
- Harari, A.R.; Zahavi, T.; Steinitz, H. Female detection of the synthetic sex pheromone contributes to the efficacy of mating disruption of the European grapevine moth, Lobesia botrana. Pest Manag. Sci. 2015, 71, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T. Principles of mating disruption. In Behavior-Modifying Chemicals for Pest Management: Applications of Pheromones and Other Attractants; Marcel Dekker: New York, NY, USA, 1990; pp. 47–71. [Google Scholar]
- Bartell, R.J. Mechanisms of communication disruption by pheromone in the control of Lepidoptera: A review. Physio. Entom. 1982, 7, 353–364. [Google Scholar]
- Justum, A.R.; Gordon, R.F.S. Pheromones: Importance to insects and role in pest management. In Insect Pheromones in Plant Protection; John Wiley & Sons: New York, NY, USA, 1989; pp. 1–13. [Google Scholar]
- Ridgway, R.L.; Inscoe, M.N.; Dickerson, W.A. Role of the boll weevil pheromone in pest management. In Behavior-Modifying Chemicals for Insect Management; Ridgway, R.L., Silverstein, R.M., Inscoe, M., Eds.; Marcel Dekker: New York, NY, USA, 1990; pp. 437–471. [Google Scholar]
- Teja, R.R. Novel Green Technologies for Pest Control. Just. Agric. 2022, 2, 1–5. [Google Scholar]
- Pasquier, D.; Charmillot, P. Survey of pheromone emission from different kinds of dispensers used for mating disruption in orchards and vineyards. IOBC WPRS Bull. 2005, 9, 335–339. [Google Scholar]
- Ioriatti, C.; Bagnoli, B.; Lucchi, A.; Veronelli, V. Vine moths control by mating disruption in Italy: Results and future prospects. Redia 2005, 87, 117–128. [Google Scholar]
- EPPO. PP 1/264 (2) Principles of efficacy evaluation for mating disruption pheromones. EPPO Bull. 2019, 49, 426–430. [Google Scholar] [CrossRef]
- Sauer, A.E.; Karg2, G. Variables affecting pheromone concentration in vineyards treated for mating disruption of grape vine moth Lobesia botrana. J. Chem. Ecol. 1998, 24, 289–302. [Google Scholar] [CrossRef]
- Koch, U.T.; Quasthoff, M.; Klemm, M.; Becker, J. Methods for reliable measurement of pheromone dispenser performance. IOBC WPRS Bull. 2002, 25, 95–99. [Google Scholar]
- Hofmeyr, J.H.; Burger, B.V. Controlled-release pheromone dispenser for use in traps to monitor flight activity of false codling moth. J. Chem. Ecol. 1995, 21, 355–363. [Google Scholar] [CrossRef]
- Zhu, H.; Thistle, H.W.; Ranger, C.M.; Zhou, H.; Strom, B.L. Measurement of semiochemical release rates with a dedicated environmental control system. Biosyst. Eng. 2015, 129, 277–287. [Google Scholar] [CrossRef]
- Vidal, A.G. Cuantificación de Feromonas en Aire y su Aplicación en Métodos de Control de Plagas. Ph.D. Thesis, Universitat Politècnica de València, Valencia, Spain, 2021. [Google Scholar]
- Akaike, H. Likelihood of a model and information criteria. J. Econ. 1981, 16, 3–14. [Google Scholar] [CrossRef]
- Lin, L.I. A concordance correlation coefficient to evaluate reproducibility. Biometrcs 1989, 45, 255–268. [Google Scholar] [CrossRef]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models part I–a discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Signorell, A. DescTools: Tools for Descriptive Statistics, R Package Version 0.99.53. 2024.
- Madden, L.V.; Gareth, H.; van den, B.F. The Study of Plant Disease Epidemics; APS Publications: Rockville, MD, USA, 2017; p. 421. [Google Scholar]
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth Stages of the Grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Aust. J. Grape. Wine. Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- Buck, A.L. New equations for computing vapor pressure and enhancement factor. J. Appl. Meteorol. Climatol. 1981, 20, 1527–1532. [Google Scholar] [CrossRef]
- Tobin, P.C.; Zhang, A.; Onufrieva, K.; Leonard, D.S. Field evaluation of effect of temperature on release of disparlure from a pheromone-baited trapping system used to monitor gypsy moth (Lepidoptera: Lymantriidae). J. Econ. Entomol. 2011, 104, 1265–1271. [Google Scholar] [CrossRef]
- Klassen, D.; Lennox, M.D.; Dumont, M.J.; Chouinard, G.; Tavares, J.R. Dispensers for pheromonal pest control. J. Environ. Manag. 2023, 325, 116590. [Google Scholar] [CrossRef]
- Il’ichev, A.L.; Williams, D.G.; Gut, L.J. Dual pheromone dispenser for combined control of codling moth Cydia pomonella L. and oriental fruit moth Grapholita molesta (Busck) (Lep., Tortricidae) in pears. J. Appl. Entomol. 2007, 131, 368–376. [Google Scholar] [CrossRef]
- Stelinski, L.L.; McGhee, P.; Grieshop, M.; Brunner, J.; Gut, L.J. Efficacy and mode of action of female-equivalent dispensers of pheromone for mating disruption of codling moth. Agric. For. Entomol. 2008, 10, 389–397. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex pheromone aerosol devices for mating disruption: Challenges for a brighter future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Evenden, M.L. Mating disruption of moth pests in integrated pest management: A mechanistic approach. In Pheromone Communication in Moths: Evolution, Behavior and Application; University of California Press: Oakland, CA, USA, 2019; Volume 24, pp. 365–394. [Google Scholar]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [PubMed]
- Akyol, B.; Aslan, M.M. Investigations on Efficiency of Mating Disruption Technique Against the European Grapevine Moth (Lobesia botrana Den. Et. Schiff.) (Lepidoptera; Tortricidae) in Vineyard, Turkey. J. Vet. Adv. 2010, 9, 730–735. [Google Scholar] [CrossRef]
- Morando, A.; Lavezzaro, S.; Ferro, S. Controllo della tignoletta con il metodo della confusione sessuale in Piemonte. Atti Giorante Fitopatol. 2016, 1, 273–280. [Google Scholar]
- Vassiliou, V.A. Control of Lobesia botrana (Lepidoptera: Tortricidae) in vineyards in Cyprus using the Mating Disruption Technique. Crop Prot. 2009, 28, 145–150. [Google Scholar] [CrossRef]
- Leonhardt, B.A.; Dickerson, W.A.; Ridgway, R.L.; Devilbiss, E.D. Laboratory and Field Evaluation of Controlled Release Dispensers Containing Grandlure, the Pheromone of the Boll Weevil (Coleoptera: Curculionidae). J. Econ. Entomol. 1988, 81, 937–943. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Anfora, G.; Bagnoli, B.; Botton, M.; Campos-Herrera, R.; Cristina, C.; Matthew, P.D.; César, G.; Ally, R.H.; et al. European grapevine moth, Lobesia botrana Part II: Prevention and management. Entomol. Gen. 2023, 43, 281–304. [Google Scholar] [CrossRef]
- Ferrer, B.F. Desenvolupament D’emissors Degradables de les Feromones de Lobesia Botrana denis i Schiffermüller i Cydia Pomonella Linnaeus (Lepidoptera Tortricidae) per a la Tècnica de Confusió Sexual. Avaluació de la Seua Eficàcia al Camp. Ph.D. Thesis, Universitat Politècnica de València, Valencia, Spain, 2011. [Google Scholar]
- Dhari, R.; Elem, O. Surface Area to Volume Ratio Calculator. Available online: https://www.omnicalculator.com/math/surface-area-volume-ratio (accessed on 5 July 2024).
- Mah, J.J. VPD Calculator & Chart (Vapor Pressure Deficit). Available online: https://www.omnicalculator.com/biology/vapor-pressure-deficit (accessed on 5 July 2024).
- Dolcetta, R.A.C. Physics of Fluids; Springer Nature: London, UK, 2023; pp. 1–136. [Google Scholar]
- Arn, H.; Louis, F. Mating disruption in European vineyards. In Insect Pheromone Research; Cardé, R.T., Minks, A.K., Eds.; Springer: Boston, MA, USA, 1997. [Google Scholar]
- Mcdonough, L.M.; Brown, D.F.; Aller, W.C. Insect sex pheromones effect of Temperature on Evaporation Rates of Acetates from Rubber Septa. J. Chem. Ecol. 1989, 15, 779–790. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Sansom, C.E.; Larsen, L.; Worner, S.P.; Rostás, M.; Chapman, R.B.; Butler, R.C.; de Kogel, W.J.; Davidson, M.M.; Perry, N.B.; et al. Volatile compounds as insect lures: Factors affecting release from passive dispenser systems. N. Z. J. Crop Hortic. Sci. 2019, 47, 208–223. [Google Scholar] [CrossRef]
- Pop, L.; Arn, H.; Buser, H.R. Determination of release rates of pheromone dispensers by air sampling with C-18 bonded silica. J. Chem. Ecol. 1993, 19, 2513–2519. [Google Scholar] [CrossRef]
- Moschos, T.; Souliotis, C.; Broumas, T.; Kapothanassi, V. Control of the European grapevine moth Lobesia botrana in Greece by the mating disruption technique: A three-year survey. Phytoparasitica 2004, 32, 83–96. [Google Scholar] [CrossRef]
- der Kraan, C.V.; Ebbers, A. Release rates of tetradecen-1-ol acetates from polymeric formulations in relation to temperature and air velocity. J. Chem. Ecol. 1990, 16, 1041–1058. [Google Scholar] [CrossRef] [PubMed]
- Kydonieus, A.F. Insect Suppression with Controlled Release Pheromone Systems, 1st ed.; CRC Press: Boca Raton, FL, USA, 1982; pp. 1–328. [Google Scholar]
- Wiesner, C.J.; Wiesner, C.J. Monitoring the Performance of Eastern Spruce Budworm Pheromone Formulations. Insect Pheromone Technol. Chem. Appl. 1982, 11, 209–218. [Google Scholar]
- IOBC/WPRS Working Group. Use of Pheromones and Other Semiochemicals in Integrated Control. In Proceedings of the Technology Transfer In Mating Disruption Proceedings Working Group Meeting, Montpellier, France, 9–10 September 1996. [Google Scholar]
- Johansson, B.G.; Anderbrant, O.L.L.E.; Simandl, J.; Avtzis, N.D.; Salvadori, C.; HedenstrÖm, E.; Edlund, H.; HÖgberg, H.E. Release rates for pine sawfly pheromones from two types of dispensers and phenology of Neodiprion sertifer. Neodiprion Sertifer. J. Chem. Ecol. 2001, 27, 733–745. [Google Scholar] [CrossRef]
- Shem, P.M.; Shiundu, P.M.; Gikonyo, N.K.; Ali, A.H.; Saini, R.K. Release Kinetics of a Synthetic Tsetse Allomone Derived from Waterbuck Odour from a Tygon silicon dispenser under Laboratory and Semifield conditions. Am.-Eurasian J. Agric. Environ. Sci. 2009, 6, 625–636. [Google Scholar]
- Alfaro, C.; Domínguez, J.; Navarro-Llopis, V.; Primo, J. Evaluation of trimedlure dispensers by a method based on thermal desorption coupled with gas chromatography-mass spectrometry. J. Appl. Entomol. 2008, 132, 772–777. [Google Scholar] [CrossRef]
- Lalic, B.; Eitzinger, J.; Marta, A.D.; Orlandini, S.; Sremac, A.F.; Pacher, B. Agricultural Meteorology and Climatology; Firenze University Press: Florence, Italy, 2018; pp. 1–352. [Google Scholar]
- Gavara, A.; Navarro-Llopis, V.; Primo, J.; Vacas, S. Influence of weather conditions on Lobesia botrana (Lepidoptera: Tortricidae) mating disruption dispensers’ emission rates and efficacy. Crop Prot. 2022, 1, 155. [Google Scholar] [CrossRef]
- Karg, G.; Suckling, D.M.; Bradley, S.J. Absorption and release of pheromone of Epiphyas postvittana (Lepidoptera: Tortricidae) by apple leaves. J. Chem. Ecol. 1994, 20, 1825–1841. [Google Scholar] [CrossRef]
- Schmitz, V.; Roehrich, R.; Stockel, J.P. Etude du mécanisme de la confusion sexuelle chez l’Eudémis de la Vigne Lobesia botrana Den. et Schiff. Rôles respectifs de la compétition, du camouflage, de la piste odorante et de la modification du signal phéromonal. J. Appl. Entom. 1995, 119, 131–138. [Google Scholar] [CrossRef]
- Schmitz, V.; Renou, M.; Roehrich, R.; Stockel, J.; Lecharpentier, P. Disruption mechanisms of pheromone communication in the European grape moth Lobesia botrana Den & Schiff. III. Sensory adaptation and habituation. J. Chem. Ecol. 1997, 23, 83–96. [Google Scholar]
- Cross, J.H.; Cross, J.H. A vapor collection and thermal desorption method to measure semiochemical release rates from controlled release formulations. J. Chem. Ecol. 1980, 6, 781–787. [Google Scholar] [CrossRef]
- Ozlem Altindisli, F.; Ozsemerci, F.; Koclu, T.; Akkan, Ü.; Keskin, N. Isonet LTT, a new alternative material for mating disruption of Lobesia botrana (Den. & Schiff.) in Turkey. BIO Web Conf. 2016, 7, 01029. [Google Scholar]
- Marchesini, E.; Maines, R.; Angeli, G. Controllare le tignole della vite con il disorientamento sessuale. L’informatore Agrar. 2007, 17, 52–56. [Google Scholar]
- Gilioli, G.; Pasquali, S.; Marchesini, E. A modelling framework for pest population dynamics and management: An application to the grape berry moth. Ecol. Modell. 2016, 320, 348–357. [Google Scholar] [CrossRef]
- Bleyer, G.; Kassemeyer, H.H.; Breuer, M.; Dubuis, P.H.; Viret, O.; Naef, A.; Krause, R. Prediction of Population Dynamics of the Grape Berry Moth (Eupoecilia ambiguella) and the European Grapevine Moth (Lobesia botrana) Using the Simulation Model “TWickler”. In Proceedings of the Deutsche Pflanzenschutztagung “Pflanzenschutz—Alternativlos”, Braunschweig, Germany, 10–14 September 2012. [Google Scholar]
- Bradley, S.J.; Suckling, D.M.; McNaughton, K.G.; Wearing, C.H.; Karg, G. A temperature-dependent model for predicting release rates of pheromone from a polyethylene tubing dispenser. J. Chem. Ecol. 1995, 21, 745–760. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T (°C) | Isonet | |||||
|---|---|---|---|---|---|---|
| Wi a | Wf b | τ c | NWf d | Slope e | Radj2, f | |
| 5 | 1.405 | 1.367 | - | 0.927 | 0.004 | 0.998 |
| 10 | 1.382 | 1.291 | - | 0.819 | 0.011 | 0.989 |
| 15 | 1.401 | 1.292 | - | 0.792 | 0.011 | 0.989 |
| 20 | 1.8 | 1.203 | - | 0.625 | 0.022 | 0.997 |
| 25 | 1.398 | 1.117 | - | 0.461 | 0.030 | 0.996 |
| 30 | 1.396 | 0.917 | 0.876 | 0.076 | 0.058 | 0.997 |
| T (°C) | Rak | |||||
| Wi a | Wf b | τ c | NWf d | Slope e | Radj2, f | |
| 5 | 4.878 | 4.864 | - | 0.966 | 0.002 | 0.977 |
| 10 | 4.940 | 4.877 | - | 0.872 | 0.007 | 0.984 |
| 15 | 4.932 | 4.852 | - | 0.831 | 0.008 | 0.928 |
| 20 | 4.859 | 4.748 | - | 0.720 | 0.017 | 0.996 |
| 25 | 4.787 | 4.618 | - | 0.434 | 0.033 | 0.976 |
| 30 | 4.841 | 4.503 | 4.434 | 0.111 | 0.054 | 0.998 |
| Dispenser | Parameters a | Statistics b | ||||
|---|---|---|---|---|---|---|
| α | β | Radj2 | RMSE | CRM | CCC | |
| Isonet | 3.597 × 10−4 | 1.031 × 10−1 | 0.980 | 3.224 × 10−4 | 1.258 × 10−2 | 0.992 |
| Rak | 2.172 × 10−4 | 1.213 × 10−2 | 0.995 | 1.620 × 10−4 | 4.924 × 10−4 | 0.998 |
| X a | Parameters | Selection Criteria b | Goodness-of-Fit c | |||||
|---|---|---|---|---|---|---|---|---|
| α | β | SEest | Radj2 | AIC | RMSE | CRM | CCC | |
| 1. DOY | 1.986 | −0.007 | 0.055 | 0.965 | −433 | 0.061 | 0.026 | 0.979 |
| 2. ΣPRR | 0.993 | −1.662 | 0.046 | 0.975 | −458 | 0.047 | 0.003 | 0.987 |
| 3. ΣDD | 1.006 | −0.00028 | 0.048 | 0.973 | −454 | 0.050 | 0.004 | 0.985 |
| 4. ΣDP | 0.955 | −0.00333 | 0.145 | 0.758 | −288 | 0.142 | −0.002 | 0.865 |
| 5. ΣVPD | 0.984 | −0.00053 | 0.075 | 0.935 | −387 | 0.074 | 0.002 | 0.967 |
| 6. ΣPRR and ΣVPD | 0.089 | −3.441 0.00058 | 0.032 | 0.988 | −522 | 0.031 | 0.001 | 0.994 |
| X a | Parameters b | Selection Criteria c | Goodness-of-Fit d | ||||||
|---|---|---|---|---|---|---|---|---|---|
| α | β1 | β2 | SEest | Radj2 | AIC | RMSE | CRM | CCC | |
| 1. DOY | 3.885 | −0.027 | 0.049 | 0.121 | 0.832 | −314 | 0.114 | −0.008 | 0.918 |
| 2. ΣPRR | 0.982 | −3.126 | 3.034 | 0.123 | 0.827 | −311 | 0.115 | −0.002 | 0.915 |
| 3. ΣDD | 1.014 | −0.005 | 0.00008 | 0.121 | 0.832 | −314 | 0.114 | −0.002 | 0.917 |
| 4. ΣDP | 1.026 | −0.0068 | 0.013 | 0.140 | 0.776 | −292 | 0.136 | 0.002 | 0.879 |
| 5. ΣVPD | 0.963 | −0.0008 | 0.00021 | 0.118 | 0.840 | −317 | 0.110 | −0.001 | 0.923 |
| 6. ΣPRR2 ΣVPD | 0.974 | 2.456 −0.0009 | − | 0.111 | 0.859 | −327 | 0.102 | −0.001 | 0.935 |
| 7. ΣPRR2 ΣVPD PC17 CA18 VC18 | 1.095 | 2.171 −0.0009 −0.059 −0.260 −0.185 | − | 0.039 | 0.982 | −480 | <0.001 | −0.001 | 0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corbetta, M.; Bricchi, L.; Rossi, V.; Fedele, G. The Dynamics of Pheromone Release in Two Passive Dispensers Commonly Used for Mating Disruption in the Control of Lobesia botrana and Eupoecilia ambiguella in Vineyards. Insects 2024, 15, 962. https://doi.org/10.3390/insects15120962
Corbetta M, Bricchi L, Rossi V, Fedele G. The Dynamics of Pheromone Release in Two Passive Dispensers Commonly Used for Mating Disruption in the Control of Lobesia botrana and Eupoecilia ambiguella in Vineyards. Insects. 2024; 15(12):962. https://doi.org/10.3390/insects15120962
Chicago/Turabian StyleCorbetta, Marta, Luca Bricchi, Vittorio Rossi, and Giorgia Fedele. 2024. "The Dynamics of Pheromone Release in Two Passive Dispensers Commonly Used for Mating Disruption in the Control of Lobesia botrana and Eupoecilia ambiguella in Vineyards" Insects 15, no. 12: 962. https://doi.org/10.3390/insects15120962
APA StyleCorbetta, M., Bricchi, L., Rossi, V., & Fedele, G. (2024). The Dynamics of Pheromone Release in Two Passive Dispensers Commonly Used for Mating Disruption in the Control of Lobesia botrana and Eupoecilia ambiguella in Vineyards. Insects, 15(12), 962. https://doi.org/10.3390/insects15120962