
Molecular Diversity and Distribution of Whiteflies (Bemisia tabaci) in Cassava Fields Across South West and North Central, Nigeria

 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area, Field Selection, and Sampling Criteria
2.2. Whitefly DNA Extraction
2.3. Mitochondrial COI (mtCOI) Amplification and Sequencing
2.4. Determination of Whitefly Genotype and Phylogenetic Analyses
2.5. Population Genetic Analysis
2.6. Polymerase Chain Reaction (PCR) Amplification for CMB Detection
3. Results
3.1. Genetic Diversity of Whiteflies
3.2. Population Genetic Diversity
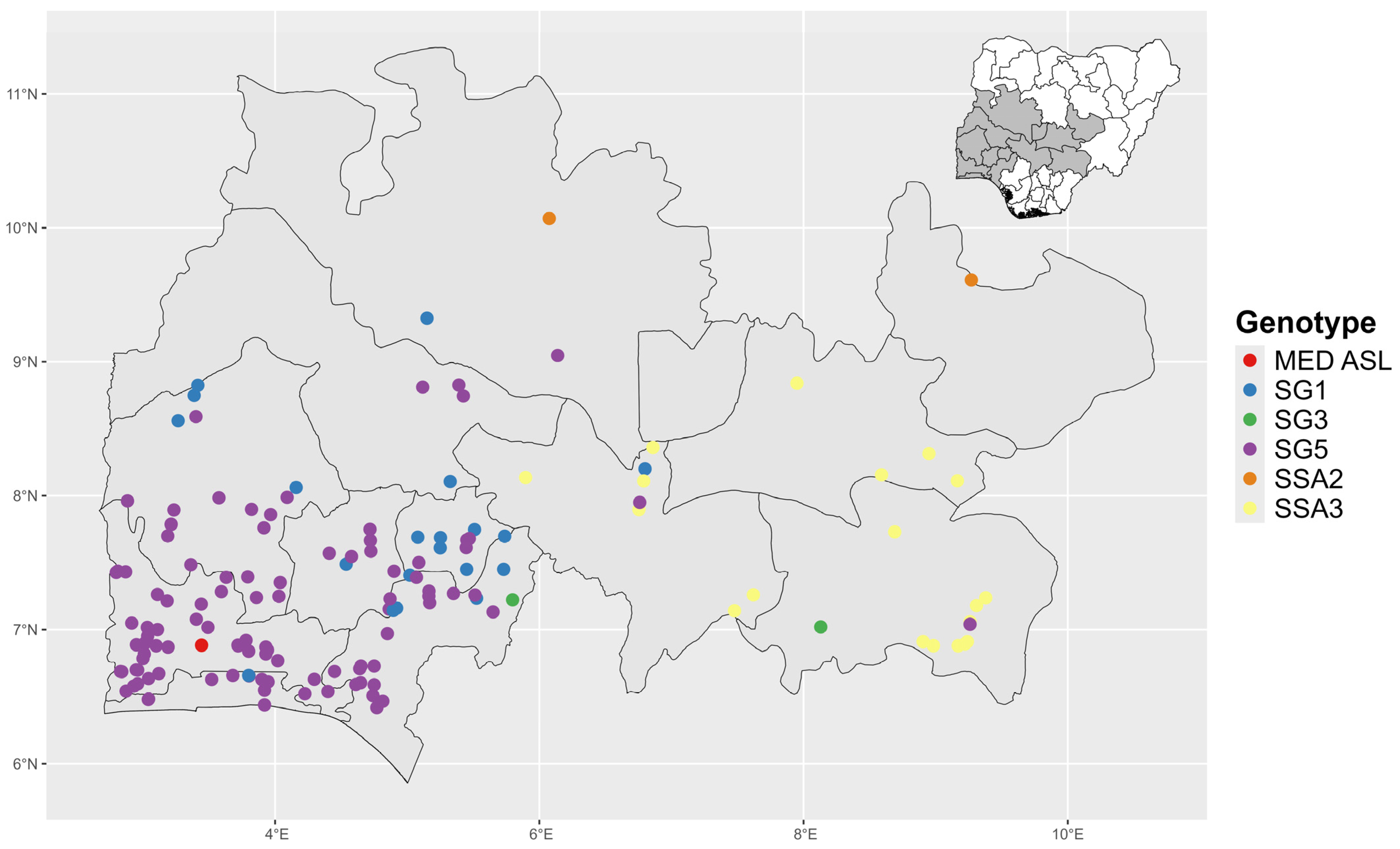
3.3. Distribution of Identified Whitefly Genotypes Across the Surveyed Zones
3.4. Viruses Detected in Whiteflies Sampled from Cassava in South West and North Central, Nigeria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feyisa, A. Current Status, Opportunities and Constraints of Cassava Production in Ethiopia-A Review. J. Agric. Sci. Food Res. 2021, 11, 051. [Google Scholar]
- Alonso-Chavez, V.; Milne, A.E.; van den Bosch, F.; Pita, J.S.; McQuaid, C.F. Modelling Cassava Production and Pest Management under Biotic and Abiotic Constraints. Plant Mol. Biol. 2021, 109, 325–349. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Published 2021; FAO: Rome, Italy, 2021; Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 23 June 2024).
- Thresh, J.M.; Cooter, R.J. Strategies for Controlling Cassava Mosaic Virus Disease in Africa. Plant Pathol. 2005, 54, 587–614. [Google Scholar] [CrossRef]
- Chikoti, P.C.; Mulenga, R.M.; Tembo, M.; Sseruwagi, P. Cassava Mosaic Disease: A Review of a Threat to Cassava Production in Zambia. J. Plant Pathol. 2019, 101, 467–477. [Google Scholar] [CrossRef]
- Shirima, R.R.; Wosula, E.N.; Hamza, A.A.; Mohammed, N.A.; Mouigni, H.; Nouhou, S.; Mchinda, N.M.; Ceasar, G.; Amour, M.; Njukwe, E.; et al. Epidemiological Analysis of Cassava Mosaic and Brown Streak Diseases, and Bemisia Tabaci in the Comoros Islands. Viruses 2022, 14, 2165. [Google Scholar] [CrossRef]
- Eni, A.O.; Efekemo, O.P.; Onile-ere, O.A.; Pita, J.S. South West and North Central Nigeria: Assessment of Cassava Mosaic Disease and Field Status of African Cassava Mosaic Virus and East African Cassava Mosaic Virus. Ann. Appl. Biol. 2021, 178, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Ally, H.M.; Hamss, H.E.; Simiand, C.; Maruthi, M.N.; Colvin, J.; Omongo, C.A.; Delatte, H. What Has Changed in the Outbreaking Populations of the Severe Crop Pest Whitefly Species in Cassava in Two Decades? Sci. Rep. 2019, 9, 14796. [Google Scholar] [CrossRef] [PubMed]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, C.A.; Kabaalu, R.; Visendi, P.; Sseruwagi, P.; Seal, S.E. Genetic Diversity of Whitefly (Bemisia spp.) on Crop and Uncultivated Plants in Uganda: Implications for the Control of This Devastating Pest Species Complex in Africa. J. Pest Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Casinga, C.M.; Shirima, R.R.; Mahungu, N.M.; Tata-Hangy, W.; Bashizi, K.B.; Munyerenkana, C.M.; Ughento, H.; Enene, J.; Sikirou, M.; Dhed’a, B.; et al. Expansion of the Cassava Brown Streak Disease Epidemic in Eastern Democratic Republic of Congo. Plant Dis. 2021, 105, 2177–2188. [Google Scholar] [CrossRef]
- Leiva, A.M.; Chittarath, K.; Lopez-Alvarez, D.; Vongphachanh, P.; Gomez, M.I.; Sengsay, S.; Wang, X.-W.; Rodriguez, R.; Newby, J.; Cuellar, W.J. Mitochondrial Genetic Diversity of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) Associated with Cassava in Lao PDR. Insects 2022, 13, 861. [Google Scholar] [CrossRef]
- Elfekih, S.; Tay, W.T.; Polaszek, A.; Gordon, K.H.J.; Kunz, D.; Macfadyen, S.; Walsh, T.K.; Vyskočilová, S.; Colvin, J.; De Barro, P.J. On Species Delimitation, Hybridization and Population Structure of Cassava Whitefly in Africa. Sci. Rep. 2021, 11, 7923. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.L.; Duffy, S.; Sseruwagi, P. Whitefly-Transmitted Viruses Threatening Cassava Production in Africa. Curr. Opin. Virol. 2018, 33, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Casinga, C.M.; Wosula, E.N.; Sikirou, M.; Shirima, R.R.; Munyerenkana, C.M.; Nabahungu, L.N.; Bashizi, B.K.; Ugentho, H.; Monde, G.; Legg, J.P. Diversity and Distribution of Whiteflies Colonizing Cassava in Eastern Democratic Republic of Congo. Insects 2022, 13, 849. [Google Scholar] [CrossRef] [PubMed]
- Munguti, F.M.; Kilalo, D.C.; Nyaboga, E.N.; Wosula, E.N.; Macharia, I.; Mwango’mbe, A.W. Distribution and Molecular Diversity of Whitefly Species Colonizing Cassava in Kenya. Insects 2021, 12, 875. [Google Scholar] [CrossRef]
- Nwezeobi, J.; Onyegbule, O.; Nkere, C.; Onyeka, J.; Van Brunschot, S.; Seal, S.; Colvin, J. Cassava Whitefly Species in Eastern Nigeria and the Threat of Vector-Borne Pandemics from East and Central Africa. PLoS ONE 2020, 15, e0232616. [Google Scholar] [CrossRef]
- Akintola, A.A.; Hwang, H.-S.; Khatun, M.F.; Ande, A.T.; Lee, K.-Y. Genetic Diversity of Bemisia Tabaci Cryptic Species in Nigeria and Their Relationships with Endosymbionts and Acquired Begomoviruses. J. Asia Pac. Entomol. 2020, 23, 1003–1009. [Google Scholar] [CrossRef]
- Asante-Pok, A. Analysis of Incentives and Disincentives for Cassava in Nigeria. Gates Open Res. 2019, 3, 915. [Google Scholar]
- Sseruwagi, P.; Sserubombwe, W.S.; Legg, J.P.; Ndunguru, J.; Thresh, J.M. Methods of Surveying the Incidence and Severity of Cassava Mosaic Disease and Whitefly Vector Populations on Cassava in Africa: A Review. Virus Res. 2004, 100, 129–142. [Google Scholar] [CrossRef]
- Boykin, L.M.; De Barro, P.J. A Practical Guide to Identifying Members of the Bemisia Tabaci Species Complex: And Other Morphologically Identical Species. Front. Ecol. Evol. 2014, 2, 45. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Boykin, L.M.; Kinene, T.; Wainaina, J.M.; Savill, A.; Seal, S.; Mugerwa, H.; Macfadyen, S.; Tay, W.T.; De Barro, P.; Kubatko, L.; et al. Review and Guide to a Future Naming System of African Bemisia tabaci Species. Syst. Entomol. 2018, 43, 427–433. [Google Scholar] [CrossRef]
- Mugerwa, H.; Rey, M.E.C.; Alicai, T.; Ateka, E.; Atuncha, H.; Ndunguru, J.; Sseruwagi, P. Genetic Diversity and Geographic Distribution of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) Genotypes Associated with Cassava in East Africa. Ecol. Evol. 2012, 2, 2749–2762. [Google Scholar] [CrossRef] [PubMed]
- Legg, J.P.; Sseruwagi, P.; Boniface, S.; Okao-Okuja, G.; Shirima, R.; Bigirimana, S.; Gashaka, G.; Herrmann, H.-W.; Jeremiah, S.; Obiero, H.; et al. Spatio-Temporal Patterns of Genetic Change amongst Populations of Cassava Bemisia tabaci Whiteflies Driving Virus Pandemics in East and Central Africa. Virus Res. 2014, 186, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.R.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Matic, S.; Pais da Cunha, A.T.; Thompson, J.R.; Tepfer, M. Short Communication an Analysis of Viruses Associated with Cassava Mosaic Disease in Three Angolan Provinces. J. Plant Pathol. 2012, 94, 443–450. [Google Scholar] [CrossRef]
- Macfadyen, S.; Tay, W.T.; Hulthen, A.D.; Paull, C.; Kalyebi, A.; Jacomb, F.; Parry, H.; Sseruwagi, P.; Seguni, Z.; Omongo, C.A.; et al. Landscape Factors and How They Influence Whitefly Pests in Cassava Fields across East Africa. Landsc. Ecol. 2021, 36, 45–67. [Google Scholar] [CrossRef]
- Kreye, C.; Hauser, S.; Pypers, P.; Vanlauwe, B. Intensification Options of Small Holders’ Cassava Production in South-west Nigeria. Agron. J. 2020, 112, 5312–5324. [Google Scholar] [CrossRef]
- Ghosh, S.; Bouvaine, S.; Maruthi, M. Prevalence and Genetic Diversity of Endosymbiotic Bacteria Infecting Cassava Whiteflies in Africa. BMC Microbiol. 2015, 15, 93. [Google Scholar] [CrossRef]
- Alicai, T.; Omongo, C.A.; Maruthi, M.N.; Hillocks, R.J.; Baguma, Y.; Kawuki, R.; Bua, A.; Otim-Nape, G.W.; Colvin, J. Re-Emergence of Cassava Brown Streak Disease in Uganda. Plant Dis. 2007, 91, 24–29. [Google Scholar] [CrossRef]
- Alicai, T.; Szyniszewska, A.M.; Omongo, C.A.; Abidrabo, P.; Okao-Okuja, G.; Baguma, Y.; Ogwok, E.; Kawuki, R.; Esuma, W.; Tairo, F.; et al. Expansion of the Cassava Brown Streak Pandemic in Uganda Revealed by Annual Field Survey Data for 2004 to 2017. Sci. Data 2019, 6, 327. [Google Scholar] [CrossRef]
- Legg, J.P.; Shirima, R.; Tajebe, L.S.; Guastella, D.; Boniface, S.; Jeremiah, S.; Nsami, E.; Chikoti, P.; Rapisarda, C. Biology and Management of Bemisia Whitefly Vectors of Cassava Virus Pandemics in Africa. Pest Manag. Sci. 2014, 70, 1446–1453. [Google Scholar] [CrossRef] [PubMed]
- Namuddu, A.; Seal, S.; Van Brunschot, S.; Malka, O.; Kabaalu, R.; Morin, S.; Omongo, C.; Colvin, J. Distribution of Bemisia Tabaci in Different Agro-Ecological Regions in Uganda and the Threat of Vector-Borne Pandemics into New Cassava Growing Areas. Front. Sustain. Food Syst. 2023, 7, 1068109. [Google Scholar] [CrossRef]
- Chen, W.; Wosula, E.N.; Hasegawa, D.K.; Casinga, C.; Shirima, R.R.; Fiaboe, K.K.M.; Hanna, R.; Fosto, A.; Goergen, G.; Tamò, M.; et al. Genome of the African Cassava Whitefly Bemisia Tabaci and Distribution and Genetic Diversity of Cassava-Colonizing Whiteflies in Africa. Insect Biochem. Mol. Biol. 2019, 110, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Mugerwa, H.; Seal, S.; Wang, H.-L.; Patel, M.V.; Kabaalu, R.; Omongo, C.A.; Alicai, T.; Tairo, F.; Ndunguru, J.; Sseruwagi, P.; et al. African Ancestry of New World, Bemisia Tabaci-Whitefly Species. Sci. Rep. 2018, 8, 2734. [Google Scholar] [CrossRef]
- Shadmany, M.; Boykin, L.M.; Muhamad, R.; Omar, D. Genetic Diversity of Bemisia tabaci (Hemiptera: Aleyrodidae) Species Complex Across Malaysia. J. Econ. Entomol. 2019, 112, 75–84. [Google Scholar] [CrossRef]
- Sartor, C.; Demichelis, S.; Cenis, J.L.; Coulibaly, A.K.; Bosco, D. Genetic Variability of Bemisia Tabaci in the Mediterranean and Sahel Regions. Bull. Insectol. 2008, 61, 161–162. [Google Scholar]
- Pita, J.S.; Fondong, V.N.; Sangaré, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, Pseudorecombination and Synergism of Geminiviruses Are Determinant Keys to the Epidemic of Severe Cassava Mosaic Disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef]
- Alabi, O.J.; Kumar, P.L.; Naidu, R.A. Multiplex PCR for the Detection of African Cassava Mosaic Virus and East African Cassava Mosaic Cameroon Virus in Cassava. J. Virol. Methods 2008, 154, 111–120. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Whitefly Genotype | Number of Isolates | Nucleotide Identity Similarity % | Accession Numbers | Country | |
|---|---|---|---|---|---|
| SSA1 | SG1 | 20 | 99.87–100 | MW683075 MW683083 | Uganda |
| SG3 | 2 | 98.55–99.09 | MK360177 | Uganda | |
| SG5 | 101 | 98.15–100 | KM377922 KM377929 | Nigeria | |
| SSA1 Subtotal | 123 | ||||
| SSA2 | 2 | 99.45–99.64 | MK903492 | Uganda | |
| SSA3 | 19 | 99.34–100 | KM377923 KM377924 KM377926 | Nigeria | |
| MED | 1 | 100 | MH205754 | Uganda |
| Whitefly Genotype | SSA1-SG1 (%) | SSA1-SG5 (%) | SSA2 (%) | SSA3 (%) |
|---|---|---|---|---|
| SSA1-SG1 (Range) | 0.15 (0.00–0.52) | |||
| SSA1-SG5 (Range) | 1.66 (1.28–3.79) | 0.33 (0.00–3.27) | ||
| SSA2 (Range) | 7.20 (7.15–7.37) | 7.88 (7.24–9.67) | a | |
| SSA3 (Range) | 6.71 (6.51–7.24) | 6.97 (6.28–8.5) | 5.67 (5.49–6.00) | 0.53 (0.00–0.99) |
| Whitefly Genotype | n | h | S | K | Hd | π | Fu’s Fs | Tajima’s D | p Value |
|---|---|---|---|---|---|---|---|---|---|
| SG1 | 20 | 4 | 5 | 0.589 | 0.363 | 0.00078 | −1.197 | −1.78003 | 0.10 > p > 0.05 |
| SG3 | 2 | 2 | 31 | 31.000 | 1.000 | 0.03985 | 3.434 | - | - |
| SG5 | 101 | 5 | 4 | 0.079 | 0.078 | 0.00018 | −7.421 | −1.77619 | 0.10 > p > 0.05 |
| SSA1 subtotal | 123 | 8 | 24 | 2.288 | 0.354 | 0.00512 | 1.354 | −1.47570 | p > 0.10 |
| SSA2 | 2 | 2 | 2 | 2.000 | 1.000 | 0.00261 | 0.693 | - | - |
| SSA3 | 19 | 5 | 7 | 2.503 | 0.6491 | 0.00412 | 1.243 | 0.83356 | p > 0.10 |
| MED | 1 | 1 | - | - | - | - | - | - | - |
| Total | 145 | 15 | 96 | 10.645 | 0.530 | 0.02381 | 8.918 | −1.46119 | - |
| Zone | State (n) | SSA1-SG1 % | SSA1-SG3 % | SSA1-SG5 % | SSA2 % | SSA3 % | MED-ASL % |
|---|---|---|---|---|---|---|---|
| North Central | Benue (12) | 0.00 | 8.33 | 8.33 | 0.00 | 83.33 | 0.00 |
| Kogi (8) | 12.50 | 0.00 | 12.50 | 0.00 | 75.00 | 0.00 | |
| Kwara (4) | 25.00 | 0.00 | 75.00 | 0.00 | 0.00 | 0.00 | |
| Nasarawa (3) | 0.00 | 0.00 | 0.00 | 0.00 | 100.00 | 0.00 | |
| Niger (3) | 33.33 | 0.00 | 33.33 | 33.33 | 0.00 | 0.00 | |
| Plateau (1) | 0.00 | 0.00 | 0.00 | 100.00 | 0.00 | 0.00 | |
| Subtotal (31) | 9.68 | 3.23 | 19.35 | 6.45 | 61.29 | 0.00 | |
| South West | Ekiti (9) | 55.56 | 0.00 | 44.44 | 0.00 | 0.00 | 0.00 |
| Lagos (10) | 10.00 | 0.00 | 90.00 | 0.00 | 0.00 | 0.00 | |
| Ogun (40) | 0.00 | 0.00 | 97.50 | 0.00 | 0.00 | 2.50 | |
| Ondo (26) | 23.08 | 3.85 | 73.08 | 0.00 | 0.00 | 0.00 | |
| Osun (8) | 12.50 | 0.00 | 87.50 | 0.00 | 0.00 | 0.00 | |
| Oyo (21) | 19.05 | 0.00 | 80.95 | 0.00 | 0.00 | 0.00 | |
| Subtotal (114) | 14.91 | 0.88 | 83.33 | 0.00 | 0.00 | 0.88 | |
| Grand Total (145) | 13.79 | 1.38 | 69.66 | 1.38 | 13.10 | 0.69 | |
| Whitefly Genotype | ACMV | EACMV |
|---|---|---|
| SSA1-SG1 | 15% (3/20) | 0 |
| SSA1-SG3 | 0 | 0 |
| SSA1-SG5 | 10.9% (11/101) | 1% (1/101) |
| SSA2 | 0 | 0 |
| SSA3 | 21.1% (4/20) | 0 |
| MED | 0 | 0 |
| Total | 12.4% (18/145) | 0.7% (1/145) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Efekemo, O.P.; Onile-ere, O.A.; Abegunde, I.O.; Otitolaye, F.T.; Pita, J.S.; Alicai, T.; Eni, A.O. Molecular Diversity and Distribution of Whiteflies (Bemisia tabaci) in Cassava Fields Across South West and North Central, Nigeria. Insects 2024, 15, 906. https://doi.org/10.3390/insects15110906
Efekemo OP, Onile-ere OA, Abegunde IO, Otitolaye FT, Pita JS, Alicai T, Eni AO. Molecular Diversity and Distribution of Whiteflies (Bemisia tabaci) in Cassava Fields Across South West and North Central, Nigeria. Insects. 2024; 15(11):906. https://doi.org/10.3390/insects15110906
Chicago/Turabian StyleEfekemo, Oghenevwairhe P., Olabode A. Onile-ere, Isaac O. Abegunde, Folashade T. Otitolaye, Justin S. Pita, Titus Alicai, and Angela O. Eni. 2024. "Molecular Diversity and Distribution of Whiteflies (Bemisia tabaci) in Cassava Fields Across South West and North Central, Nigeria" Insects 15, no. 11: 906. https://doi.org/10.3390/insects15110906
APA StyleEfekemo, O. P., Onile-ere, O. A., Abegunde, I. O., Otitolaye, F. T., Pita, J. S., Alicai, T., & Eni, A. O. (2024). Molecular Diversity and Distribution of Whiteflies (Bemisia tabaci) in Cassava Fields Across South West and North Central, Nigeria. Insects, 15(11), 906. https://doi.org/10.3390/insects15110906