
Arsenophonus: A Double-Edged Sword of Aphid Defense against Parasitoids

 , , ,
, , ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
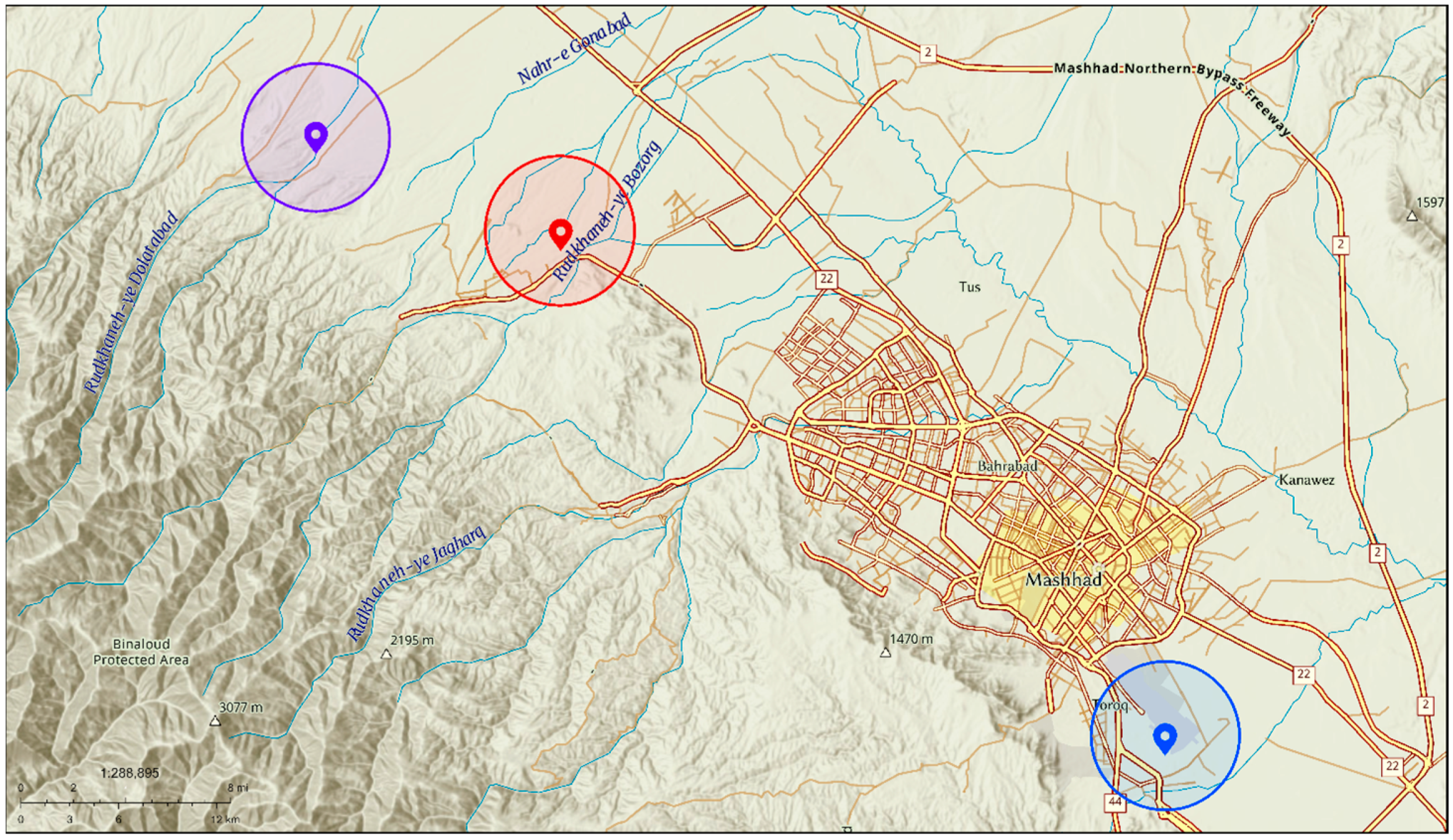
2.1. Sampling Sites
2.2. Rearing Specimens
2.2.1. Aphids
2.2.2. Rearing of BCAs from the Colonies II–VIII
2.2.3. Parasitoids
2.3. Species Identification
2.4. Images
2.5. Arsenophonus-Reduced Infection (AR)
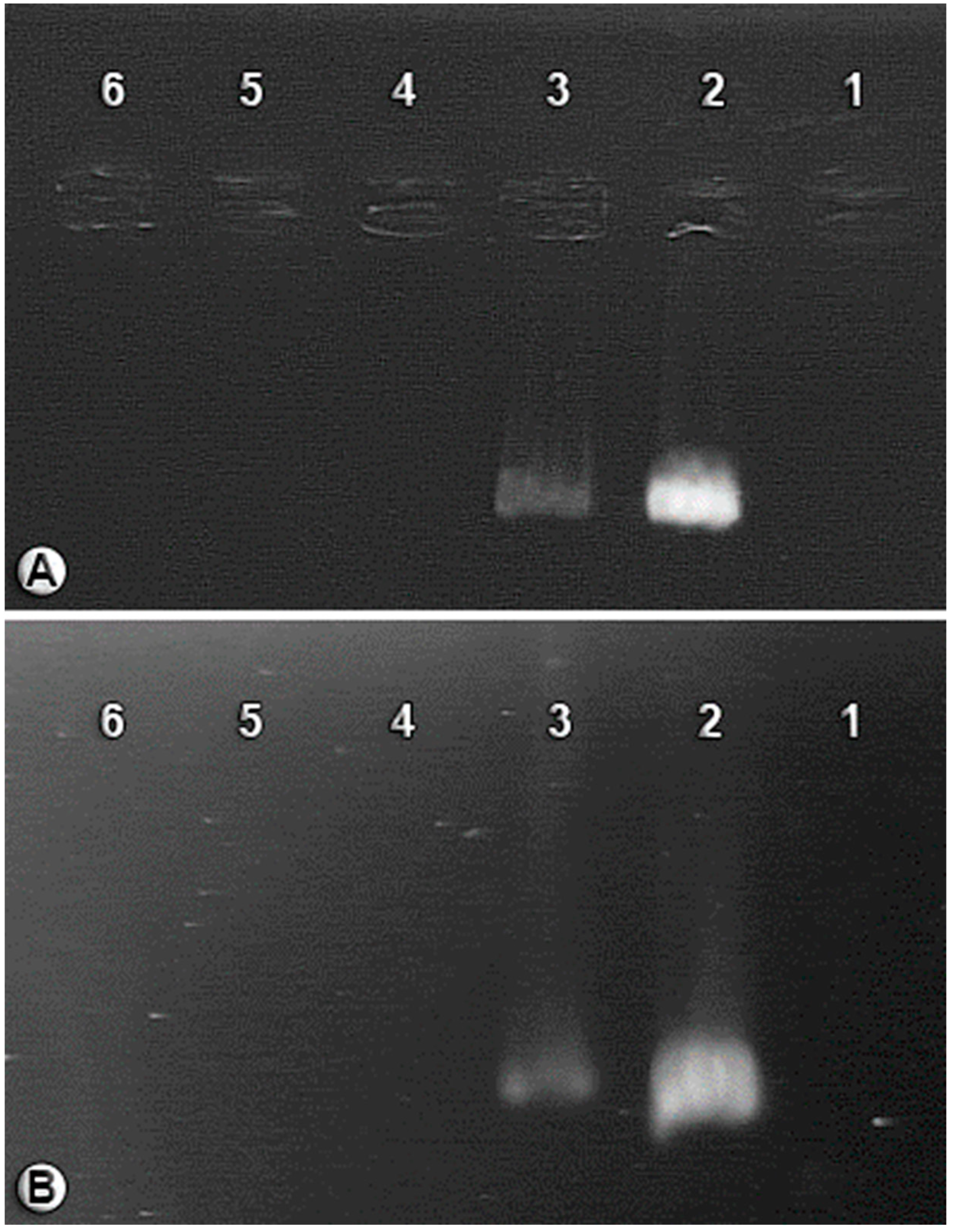
2.6. Molecular Methods
2.7. Parasitism Assay
2.8. Analysis
3. Results
3.1. BCAs Treated with an Antibiotic Mixture (Cured Aphids)
3.2. Parasitism Assays
3.2.1. Number of Mummified BCAs (Parasitism Rate) (Figure 3A and Figure 4A)
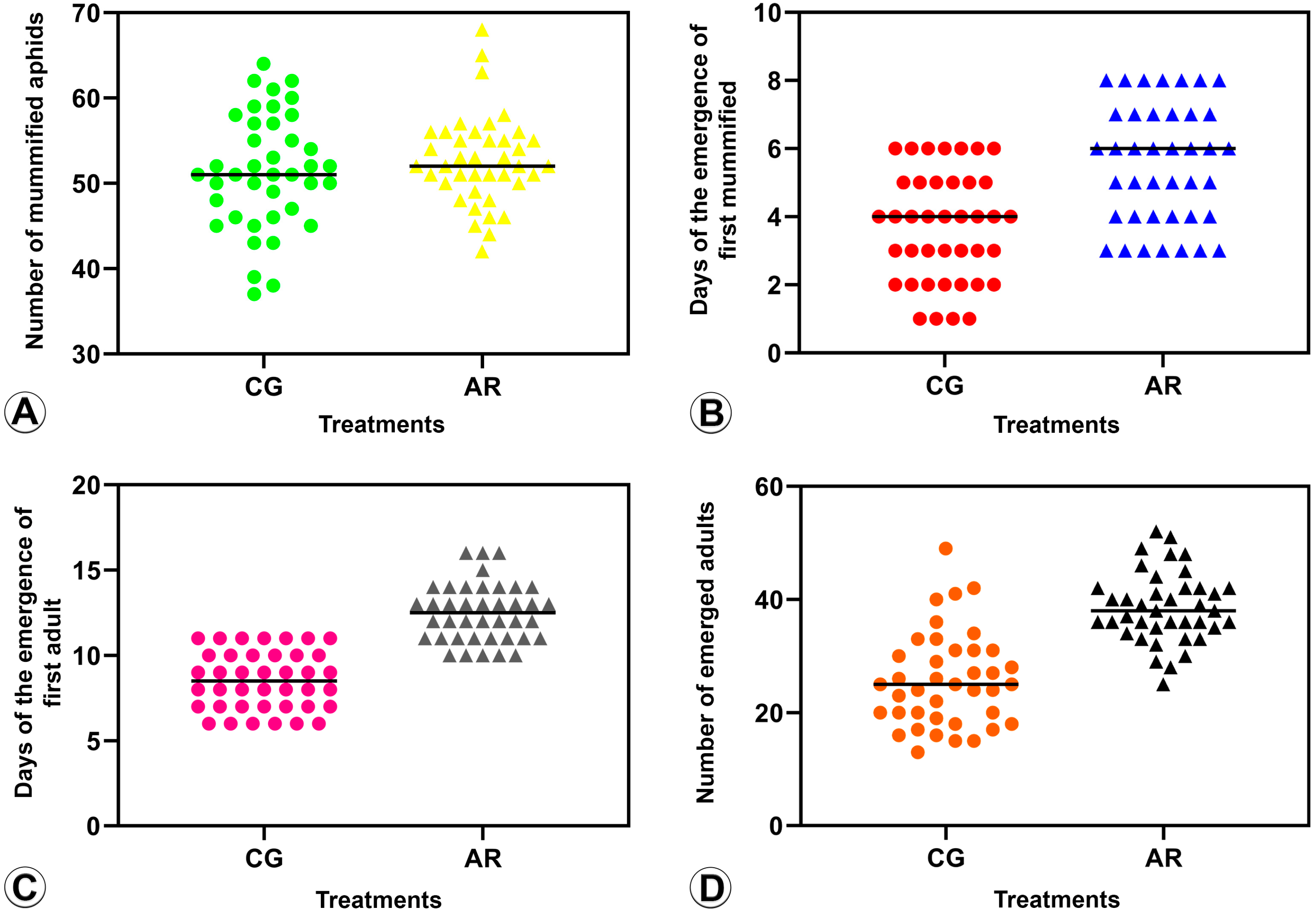
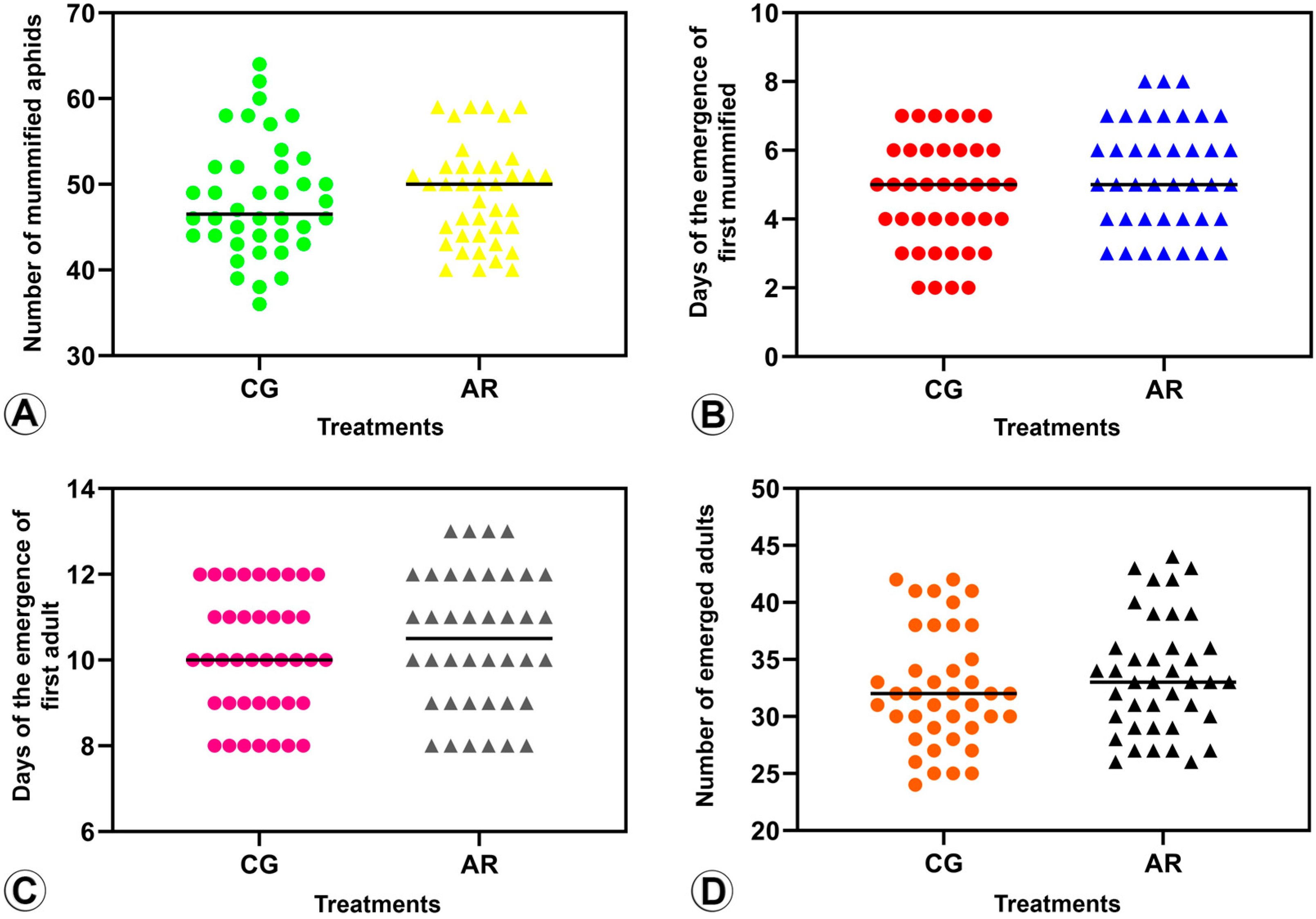
3.2.2. Pre-Imaginal Period (Figure 3B and Figure 4B)
3.2.3. Period of Mummification to Adult Emergence (Figure 3C and Figure 4C)
3.2.4. Emergence Rate (Figure 3D and Figure 4D)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hrcek, J.; McLean, A.H.; Godfray, H.C. Symbionts modify interactions between insects and natural enemies in the field. J. Anim. Ecol. 2016, 85, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.; Kaltenpoth, M. Beetle–bacterial symbioses: Endless forms most functional. Annu. Rev. Entomol. 2022, 67, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Fronk, D.C.; Sachs, J.L. Symbiotic organs: The nexus of host–microbe evolution. Trends Ecol. Evol. 2022, 37, 599–610. [Google Scholar] [CrossRef]
- Engelstadter, J.; Hurst, G.D. The ecology and evolution of microbes that manipulate host reproduction. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 127–149. [Google Scholar] [CrossRef]
- Gerardo, N.M.; Parker, B. Mechanisms of symbiont-conferred protection against natural enemies: An ecological and evolutionary framework. Curr. Opin. Insect Sci. 2014, 4, 8–14. [Google Scholar] [CrossRef]
- Hamilton, W.A.; Garretson, O.; Kerne, A. Streaming on twitch: Fostering participatory communities of play within live mixed media. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Toronto, ON, Canada, 26 April–1 May 2014; pp. 1315–1324. [Google Scholar]
- Heyworth, E.R.; Ferrari, J. A facultative endosymbiont in aphids can provide diverse ecological benefits. J. Evol. Biol. 2015, 28, 1753–1760. [Google Scholar] [CrossRef]
- Douglas, A.E. The microbiome of insects: Diversity in structure and function. Adv. Insect Phys. 2021, 60, 25–62. [Google Scholar]
- Oliver, K.M.; Martinez, A.J. How resident microbiota and host genetics shape the adaptive landscape of animal hosts. Front. Microbiol. 2020, 11, 767. [Google Scholar]
- Renoz, F.; Pons, I.; Vanderpoorten, A.; Bataille, G.; Noël, C.; Foray, V.; Pierson, V.; Hance, T. Evidence for gut-associated Serratia symbiotica in wild aphids and ants provides new perspectives on the evolution of bacterial mutualism in insects. Microb. Ecol. 2019, 78, 159–169. [Google Scholar] [CrossRef]
- Ferrari, J.; Via, S.; Godfray, H.C. Defense and counter defense in the plant–aphid–parasitoid tri-trophic system. Annu. Rev. Entomol. 2021, 66, 15–34. [Google Scholar]
- Oliver, K.M.; Smith, A.H.; Russell, J.A. Defensive symbiosis in the real world–advancing ecological studies of heritable, protective bacteria in aphids and beyond. Funct. Ecol. 2014, 28, 341–355. [Google Scholar] [CrossRef]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Coolen, S.; Rogowska-van der Molen, M.; Welte, C.U. The secret life of insect-associated microbes and how they shape insect–plant interactions. FEMS Microbiol. Ecol. 2022, 98, fiac083. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Jimenez, P.; Griffin, J.S.; Davies, L.; Frost, C.L.; Marcello, M.; Hurst, G.D. Genetic manipulation allows in vivo tracking of the life cycle of the son-killer symbiont, Arsenophonus nasoniae, and reveals patterns of host invasion, tropism and pathology. Environ. Microbiol. 2019, 21, 3172–3182. [Google Scholar] [CrossRef] [PubMed]
- Wulff, J.A.; Buckman, K.A.; Wu, K.; Heimpel, G.E.; White, J.A. The endosymbiont Arsenophonus is widespread in soybean aphid, Aphis glycines, but does not provide protection from parasitoids or a fungal pathogen. PLoS ONE 2013, 8, e62145. [Google Scholar] [CrossRef]
- Brady, C.M.; White, J.A. Cowpea aphid (Aphis craccivora) associated with different host plants has different facultative endosymbionts. Ecol. Entomol. 2013, 38, 433–437. [Google Scholar] [CrossRef]
- Pervez, A.; Harsur, M.M. Coccinellids on crops: Nature’s gift for farmers. In Innovative Pest Management Approaches for the 21st Century: Harnessing Automated Unmanned Technologies; Springer: Singapore, 2020; pp. 429–460. [Google Scholar]
- Descamps, L.R.; Sanchez-Chopa, C.; Bizet-Turovsky, J. Resistance in alfalfa to Aphis craccivora Koch. Chil. J. Agric. Res. 2015, 75, 451–456. [Google Scholar] [CrossRef]
- Heidari Latibari, M.; Moravej, G.; Ghafouri Moghaddam, M.; Barahoei, H.; Hanley, G.A. The novel host associations for the aphid parasitoid, Pauesia hazratbalensis (Hymenoptera: Braconidae: Aphidiinae). Orient. Insects 2020, 54, 88–95. [Google Scholar] [CrossRef]
- Lenhart, P.A.; White, J.A. Endosymbionts facilitate rapid evolution in a polyphagous herbivore. J. Evol. Biol. 2020, 33, 1507–1511. [Google Scholar] [CrossRef]
- Rakhshani, E.; Talebi, A.A.; Kavallieratos, N.; Fathipour, Y. Host Stage Preference, Juvenile Mortality and Functional Response of Trioxys pallidus (Haliday) (Hymenoptera: Braconidae: Aphidiinae). Biologia 2004, 59, 197–203. [Google Scholar]
- Rakhshani, H.; Rezwani, A.; Manzari, S.; Talebi, A.A.; Rakhshani, E. An investigation on alfalfa aphids and their parasitoids in different parts of Iran, with a key to the parasitoids (Hemiptera: Aphididae; Hymenoptera: Braconidae: Aphidiinae). J. Entomol. Soc. Iran. 2006, 25, 1–14. [Google Scholar]
- Yin, X.; Gurr, G.M. Biological control of aphids by parasitoids. Annu. Rev. Entomol. 2022, 67, 1–19. [Google Scholar]
- Ferracini, C.; Lozzia, G.C.; Tavella, L.; Alma, A. Exploring the potential of Aphidiinae parasitoids for the control of aphid pests in temperate horticultural crops. Biol. Control 2021, 155, 104546. [Google Scholar]
- Kahramanoglu, I.; Usanmaz, S.; Alas, T. Advances in breeding and cultivation of pomegranate. In Achieving Sustainable Cultivation of Tropical Fruits; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 569–596. [Google Scholar]
- Rakhshani, E.; Barahoei, H.; Ahmad, Z.; Starý, P.; Ghafouri-Moghaddam, M.; Mehrparvar, M.; Kavallieratos, N.G.; Čkrkić, J.; Tomanović, Ž. Review of Aphidiinae parasitoids (Hymenoptera: Braconidae) of the Middle East and North Africa: Key to species and host associations. Eur. J. Taxon. 2019, 552, 1–132. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Mitrović, M.; Petrović, A.; Kavallieratos, N.G.; Zikić, V.; Ivanović, A.; Rakhshani, E.; Starý, P.; Vorburger, C. Revision of the European Lysiphlebus species (Hymenoptera: Braconidae: Aphidiinae) on the basis of COI and 28SD2 molecular markers and morphology. Arthropod Syst. Phylogeny 2018, 76, 179–213. [Google Scholar] [CrossRef]
- Lavrinienko, A.; Watts, P.C. The role of insect-symbiont interactions in biological control programs. Curr. Opin. Insect. Sci. 2020, 42, 31–36. [Google Scholar]
- Rakhshani, E.; Talebi, A.A.; Kavallieratos, N.G.; Rezwani, A.; Manzari, S.; Tomanović, Ž. Parasitoid complex (Hymenoptera, Braconidae, Aphidiinae) of Aphis craccivora Koch (Hemiptera: Aphidoidea) in Iran. J. Pest. Sci. 2005, 78, 193–198. [Google Scholar] [CrossRef]
- Heidari Latibari, M.; Moravvej, G.; Rakhshani, E.; Karimi, J.; Arias-Penna, D.C. A host record for a strictly specific aphid parasitoid Aphidius smithi (Braconidae: Aphidiinae): The food plant-host aphid-parasitoid association puzzle acquires a new piece. Biocontrol Sci. Technol. 2020, 32, 1389–1402. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Luo, J.Y.; Wang, C.Y.; Lv, L.M.; Cui, J.J. Bacterial communities of the cotton aphid Aphis gossypii associated with Bt cotton in northern China. Sci. Rep. 2016, 6, 22958. [Google Scholar] [CrossRef]
- Duron, O. Arsenophonus insect symbionts are commonly infected with APSE, a bacteriophage involved in protective symbiosis. FEMS Microbiol. Ecol. 2014, 90, 184–194. [Google Scholar] [CrossRef]
- Starý, P.V. Key to the Genera and Species (♀♀). In Aphid Parasites (Hymenoptera, Aphidiidae) of the Central Asian Area; Springer: Dordrecht, The Netherlands, 1979. [Google Scholar]
- Lesins, K.A.; Lesins, I. Genus Medicago (Leguminosae): A Taxogenetic Study; Springer Science & Business Media: Berlin, Germany, 1979. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Plants: An Online Identification and Information Guide. 2017. Available online: www.aphidinwordsplants.info/ (accessed on 18 November 2019).
- Folmer, O.M.; Black, M.; Hoeh, W.; Luzt, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Darsouei, R.; Karimi, J.; Modarres Awal, M. First record of Aphelinus paramali Zehavi and Rosen 1989 (Hymenoptera, Aphelinidae), parasitoid of Aphis pomi de Geer (Hemiptera, Aphididae) in Iran, and its phylogenetic position based on sequence data of ITS2 and COI genes. Entomol. Res. 2011, 41, 194–200. [Google Scholar] [CrossRef]
- McLean, A.H.C.; Van Asch, M.; Ferrari, J.; Godfray, H.C.J. Effects of bacterial secondary symbionts on host plant use in pea aphids. Proc. R. Soc. B Biol. Sci. 2011, 278, 760–766. [Google Scholar] [CrossRef]
- McLean, A.H.C.; (University of Oxford, Oxford, UK). Personal communication, 2022.
- Jousselin, E.; Cœur d’Acier, A.; Vanlerberghe-Masutti, F.; Duron, O. Evolution and diversity of Arsenophonus endosymbionts in aphids. Mol. Ecol. 2013, 22, 260–270. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.R-project.org (accessed on 23 January 2022).
- Guo, J.; Hatt, S.; He, K.; Chen, J.; Francis, F.; Wang, Z. Nine facultative endosymbionts in aphids. A review. J. Asia-Pac. Entomol. 2017, 20, 794–801. [Google Scholar] [CrossRef]
- Doremus, M.R.; Stouthamer, C.M.; Kelly, S.E.; Schmitz-Esser, S.; Hunter, M.S. Cardinium localization during its parasitoid wasp host’s development provides insights into cytoplasmic incompatibility. Front. Microbiol. 2020, 11, 606399. [Google Scholar] [CrossRef] [PubMed]
- Vorburger, C.; Sandrock, C.; Ganesanandamoorthy, P. Secondary symbionts protect aphids from parasitism by destabilizing the aphid-parasitoid interaction network. Ecol. Lett. 2020, 23, 605–615. [Google Scholar]
- Khudorozhkova, Y.; Poirié, M.; Carton, Y. Parasitoid infection improves host egg and offspring survival against fungal competition. J. Evol. Biol. 2018, 31, 1044–1054. [Google Scholar]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef]
- Monticelli, L.S.; Franco, J.C.; Zepeda-Paulo, F.A.; Dion, E.; Simon, J.C.; Salem, A.Z.M. Facultative symbiont infections and plant protection in aphids: A meta-analysis. Ecol. Evol. 2019, 9, 11262–11270. [Google Scholar]
- Ferrari, J.; Vavre, F. Bacterial symbionts in insects or the story of communities affecting communities. Philos. Trans. Royal Soc. 2011, 366, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Degnan, P.H.; Hunter, M.S.; Moran, N.A. Bacteriophages encode factors required for protection in a symbiotic mutualism. Science 2016, 325, 763. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Krch, M.; Almohamad, R. Role of bacterial symbionts in the ecology of aphids and their parasitoids. In Aphid-Plant Interactions; Springer: Cham, Switzerland, 2022; pp. 59–78. [Google Scholar]
- Wang, L.; Xu, S.; Zhang, Y.; Liu, T. Effect of secondary symbionts on the parasitism of Lipaphis erysimi (Hemiptera: Aphididae) by Diaeretiella rapae (Hymenoptera: Braconidae). J. Insect Sci. 2020, 20, 1–9. [Google Scholar]
- Hua, Z.; Hou, Y.; Meng, L.; Zhu, Y.; Cui, J.; Gao, X. Removing secondary symbionts increases the susceptibility of Aphis gossypii Glover to parasitoid wasps. Insect Sci. 2021, 28, 698–708. [Google Scholar]
- Pineda, A.; Marcos-García, M.A. Microbial symbionts of aphids: Diversity, evolution and functional roles. Curr. Opin. Insect Sci. 2021, 43, 44–51. [Google Scholar]
- Huang, W.; Gao, X.; Lei, C. The role of secondary symbionts in the development of two aphid parasitoids, Aphidius gifuensis and Aphidius avenae. J. Insect Physiol. 2020, 124, 104042. [Google Scholar]
- Degnan, P.H.; Moran, N.A.; Dillman, A.R. Mutualism-defection dynamics in the Moran model for host–microbial symbiosis. J. Theor. Biol. 2016, 39, 61–83. [Google Scholar]
- Lin, X.; Lu, Y.; Zhang, Q.; Wang, Q.; Cui, J. Effects of eliminating Buchnera aphidicola on the growth, development and fecundity of a parasitoid wasp Aphidius gifuensis. J. Asia-Pac. Entomol. 2021, 24, 1164–1169. [Google Scholar]
- Gao, X.; Niu, R.; Zhu, X.; Wang, L.; Ji, J.; Niu, L.; Wu, C.; Zhang, S.; Luo, J.; Cui, J. Characterization and comparison of the bacterial microbiota of Lysiphlebia japonica parasitioid wasps and their aphid host Aphis gosypii. Pest. Manag. Sci. 2021, 77, 2710–2718. [Google Scholar] [CrossRef]
- Vorburger, C.; Gehrer, L.; Rodriguez, P. A strain of the bacterial symbiont Regiella insecticola protects aphids against parasitoids. Biol. Lett. 2010, 6, 109–111. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidari Latibari, M.; Moravvej, G.; Rakhshani, E.; Karimi, J.; Arias-Penna, D.C.; Butcher, B.A. Arsenophonus: A Double-Edged Sword of Aphid Defense against Parasitoids. Insects 2023, 14, 763. https://doi.org/10.3390/insects14090763
Heidari Latibari M, Moravvej G, Rakhshani E, Karimi J, Arias-Penna DC, Butcher BA. Arsenophonus: A Double-Edged Sword of Aphid Defense against Parasitoids. Insects. 2023; 14(9):763. https://doi.org/10.3390/insects14090763
Chicago/Turabian StyleHeidari Latibari, Minoo, Gholamhossein Moravvej, Ehsan Rakhshani, Javad Karimi, Diana Carolina Arias-Penna, and Buntika A. Butcher. 2023. "Arsenophonus: A Double-Edged Sword of Aphid Defense against Parasitoids" Insects 14, no. 9: 763. https://doi.org/10.3390/insects14090763
APA StyleHeidari Latibari, M., Moravvej, G., Rakhshani, E., Karimi, J., Arias-Penna, D. C., & Butcher, B. A. (2023). Arsenophonus: A Double-Edged Sword of Aphid Defense against Parasitoids. Insects, 14(9), 763. https://doi.org/10.3390/insects14090763