1. Introduction
Sound localization is one of the major tasks of the auditory system. In animals, it is differently solved depending on which components of the sound wave, pressure or particle velocity are being detected [
1]. Hearing systems based on pressure receivers (tympanal ears) use binaural cues such as time and level differences to localize sounds [
2]. However, the amplitude difference at the two ears, as well as the timing difference may be very small even in mammals [
3], not to mention tinier creatures such as insects. To solve this problem, two ears often become mechanically or acoustically connected, thus creating a pressure-difference receiver, which enables amplification of tiny acoustic cues into larger interaural differences, which can be detected by the nervous system [
4,
5,
6].
On the other hand, particle velocity receivers, such as antennae of mosquitoes, are inherently directional [
1,
7,
8]. The radially symmetrical structure of the Johnston’s organ (JO), a modified second segment of the insect antenna, means that only a fraction of the circular array of sensory cells are activated in response to a sound source. As the air particles move back and forth during the propagation of a sound wave, a single mosquito JO and other similarly designed auditory receivers should not be able to distinguish sounds that come from the two opposite directions. One may assume that each antenna is functionally omnidirectional and only the inter-antennal amplitude differences (IADs) are used to provide the localization cues, thus converging the functionality of particle velocity receivers to that of pressure receivers. Although the modelling experiments demonstrated that IADs in mosquito may be sufficient for the task of the sound localization [
9], the main evidence in favour of a more complex mechanism of auditory directionality in mosquitoes is an extremely large number of sensory neurons in the JO [
10], compared to insect tympanal ears [
11].
Ablation of one antenna led to a reduced ability to locate a female [
12], however, it did not completely prevent the auditory behavior. One may assume that the above-mentioned directional limitations of a single mosquito antenna could be overcome by the movement of a mosquito relative to the sound source. However, due to competition in a swarm, male mosquitoes must rapidly detect fast-flying females, which would require a mechanism of instant directionality.
A second antenna at a short distance from the first one and held parallel to it would provide little additional data on directionality. However, insects, and mosquitoes in particular, rarely hold their antennae in parallel, which opens up the possibility for additional interaural cues. The most obvious approach to instantly determine the direction to the sound source from the two opposite ones, provided by each of the two antennae, would be to compare the responses originating in the left and the right JOs based on the directional mapping in each of them. However, the binaural comparison may be rather complicated, as sound fields are usually strongly divergent close to small sound sources (such as a female mosquito), and bilateral particle velocity receivers may experience vastly different vector fields depending on the distance from the sound source [
1].
Although the JOs are morphologically symmetrical, the responses of auditory neurons may not be physiologically symmetrical. In
Culex male mosquitoes, it was demonstrated that two of the four quadrants in the JO contained more responding auditory cells [
13]. Such asymmetry may speak in favor of the presence of ‘areas of interest’ in the three-dimensional auditory space of a mosquito and may indirectly point to the mechanisms of interaural comparison. However, functional asymmetry in the JO could arise from a possible experimental bias when recording the auditory responses from only a fraction of the antennal nerve. Such a bias is hard to avoid in experiments on male mosquitoes with their large extended antennae, which limit the direction of an electrode insertion.
The limitation can be overcome by recording from the JO of a female mosquito. The antennal fibrillae of female mosquitoes are smaller and allow for the insertion of a recording electrode from multiple directions, which would significantly decrease the possible bias caused by the systematic selection of recording sites within the antennal nerve.
The question ‘What do female mosquitoes hear?’ demands much attention by itself. The auditory system of biting female mosquitoes is poorly studied compared to that of the conspecific males, the outstanding listeners. Since mosquitoes spread dangerous diseases such as West Nile fever, the understanding of their hearing system is of crucial importance. Our very limited knowledge on such a practically important subject can be at least partially explained by the fact that for a long time it was difficult to demonstrate any behavioral auditory responses in female mosquitoes, although they also possess a sophisticated auditory system, only surpassed by their male counterparts.
Pre-copulatory acoustic interaction, demonstrated for a number of mosquito species [
14,
15,
16,
17,
18], suggests that a female mosquito hears the flight tone of a male. Such hearing is possible if a female detects the distortion products of the nonlinear mixing of its own flight tone with that of a male [
15,
19,
20,
21,
22]. However, there is evidence that previously reported “harmonic convergence” events are only a random by-product of the mosquitoes’ flight tone variance and not a signature of acoustic interaction between males and females [
23].
Among bloodsucking dipterans, there are examples of distant attraction to the communication sounds of frogs in midges [
24,
25,
26] and mosquitoes [
27,
28,
29]. A study by Menda et al. [
30] suggests that the
Aedes female mosquitoes can be attracted to sound frequencies similar to those of human speech. Although the results on the female mosquito audition are far from conclusive, from the existing behavioral studies one can reason that the bloodsucking females of some dipterans, and mosquitoes in particular, can perceive the direction towards the sound source.
The lack of direct physiological data on auditory directionality in female mosquitoes, together with the above-mentioned experimental convenience, formed the basis of this work. From the previous comparative physiological study [
31] we already knew that the auditory neurons in female mosquitoes are tuned to different frequencies. Accordingly, to make a correct measure of the auditory threshold of a given unit, first we measured its tuning frequency. Then, stimulated by the sound of that frequency, we quantified the directionality of the same unit by measuring the thresholds of its response to sounds coming from different directions. Such a procedure, repeated for a significant number of auditory units, gave us the overall directional characteristics of the JO and the estimate of the angular resolution of the auditory space. To test for the hypothesized irregularities in the directional distribution of the auditory units, we studied both left and right JOs.
On the next step, to analyze the auditory space of a mosquito possessing bilateral antennae, we complemented the individual auditory properties of each JO with the data on the relative in-flight orientation of the antennae. Towards this, we photographed flying mosquitoes, measured the orientations of their antennae in two planes and estimated the variation of these parameters. The combination of the antenna measurements with the dataset on the directionality of individual auditory units implied a three-dimensional representation of the data, which we achieved by designing a graphical computer model to present the results in a simplified and easy-to-grasp form.
In three dimensions, the neuronal irregularities mirrored in the left and right JOs were found to fit well with the in-flight orientation of the antennae as to jointly provide a uniformly shaped auditory space. Here, using the mosquito JO as a model system, we describe a common methodical framework to assess directionality in similarly organized bilateral particle velocity receivers.
As the final goal of this study, we explored if there are regions in the female mosquito auditory space to which it pays special attention in terms of binaural hearing. We believe that the answer to this question will lead to the design of more adequate behavioral experiments on female mosquitoes.
2. Materials and Methods
2.1. Animals
Females of the Culex pipiens pipiens L. were captured in the wild in the Moscow region of the Russian Federation. Experiments were performed at the Kropotovo biological station (54° 51′ 2″ N; 38° 20′ 58″ E) from August to September 2016–2021.
2.2. Behavioral Experiments
Imaging of freshly captured mosquitoes was performed in the field, as we noticed that mosquitoes kept in the laboratory for one or two hours flew continuously for significantly shorter periods. We placed individual mosquitoes into a circular plastic container, diameter 85 mm, depth 26 mm. Its flat walls were left transparent, while the curved side walls were covered with non-transparent film. The container was positioned either vertically (viewed from a side) or horizontally (viewed from above). In the former position, it was lit either by sunlight or by white LED array, intensity ca. 2000 lm, both through a light-scattering paper to provide a uniform bright background for a brightfield imaging. The other flat side of the container was attached to a tube, which in turn was connected to a photo camera with a macro lens (Olympus OM-D E-M10 II, 60 mm f/2.8). The pitch and roll of the camera were minimized relative to the horizon by means of a tripod. When viewed from above, the container was lit by scattered sunlight, and the same camera mounted on a tripod and facing down was used for imaging. Lateral and dorsal imaging for the same mosquito was done sequentially, one series after another. Experiments were performed at the end of August, from 1 p.m. to 6 p.m., substantially before the swarming hours of Culex pipiens. The mating status of the female mosquitoes was not checked.
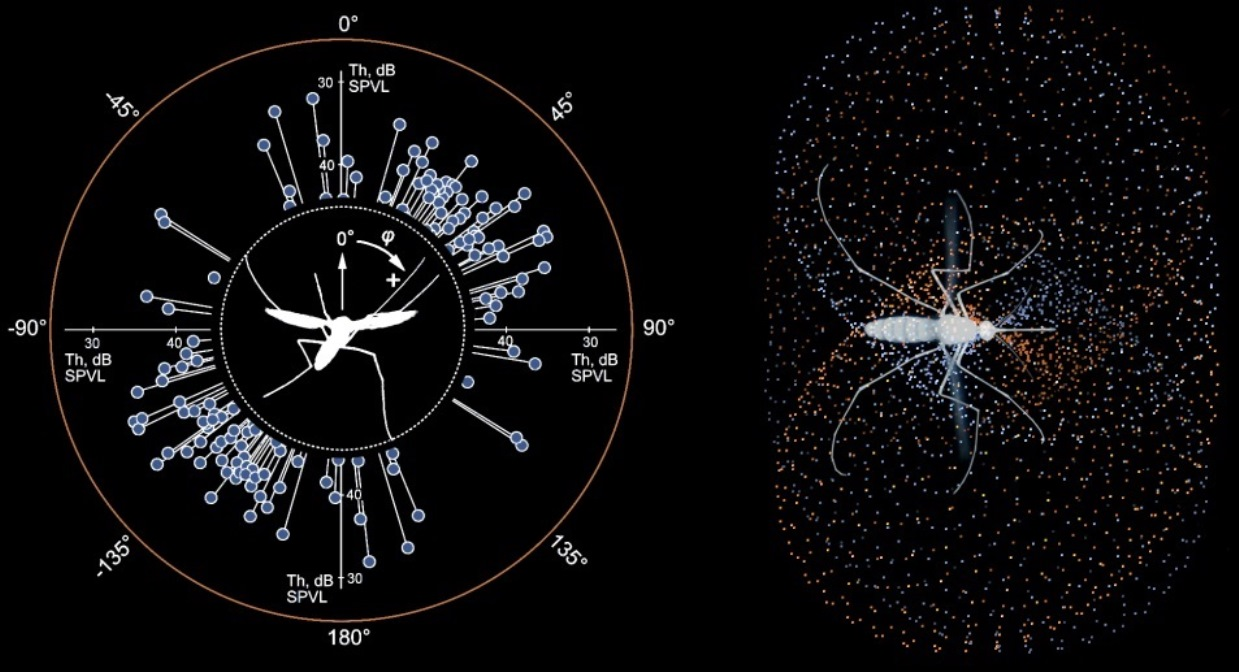
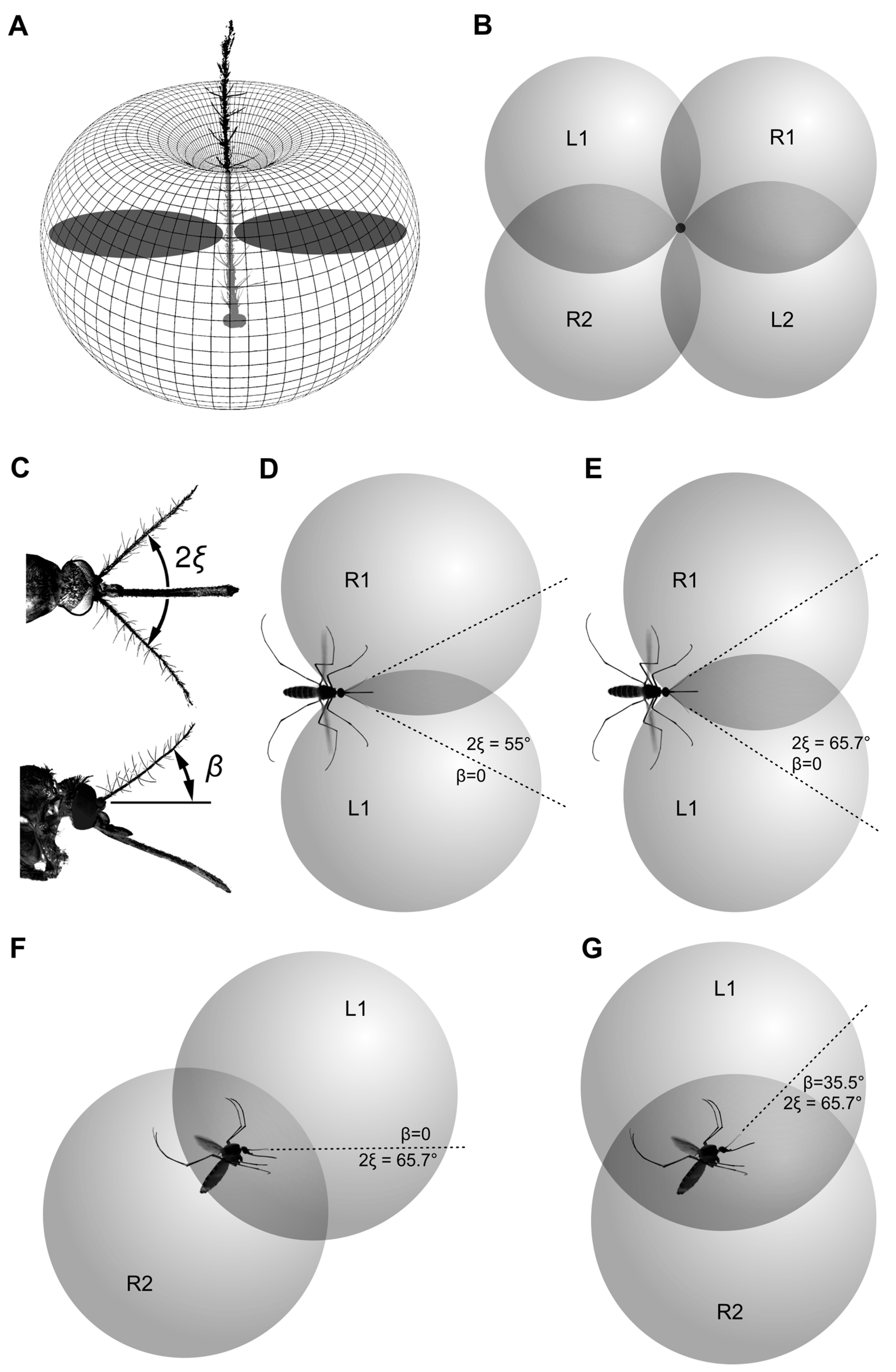
Mosquitoes (n = 22) were serially photographed during the periods of flight, either spontaneous or caused by a startle from a shadow. From the images acquired in lateral view, we selected those where a mosquito was viewed strictly laterally (the second antenna was either close to or was hidden behind the first one) and was more than 1 cm from each wall. The selected set of images (three per each mosquito) was cropped and then the following angles were manually measured using the FIJI software package, version 1.52p [
32]: antenna to horizon; abdomen to horizon; and between the two antennae (viewed from above, projection to the horizontal plane).
From the measurements in lateral view, we knew that the flying mosquito holds its antennae at an angle to the horizon. When viewed from above, this angle does not allow to directly measure the inter-antennal angle. One way to overcome this issue would be to photograph a mosquito in two projections simultaneously. However, as we required only the average value of the inter-antennal angle, we corrected the measured value using the averaged antenna-to-horizon angle according to the equation, derived from the geometry of the system:
where ξ is a half of the true inter-antennal angle, α is the measured inter-antennal angle, projected to the horizontal plane, and β is an angle between the antennal plane (a plane formed by the two antennae,
Figure 1A, dashed blue line) and the horizon.
2.3. Microelectrode Recordings
The method for measuring the directional properties of the JO sensory units was reported in detail in our previous study on male mosquitoes [
13].
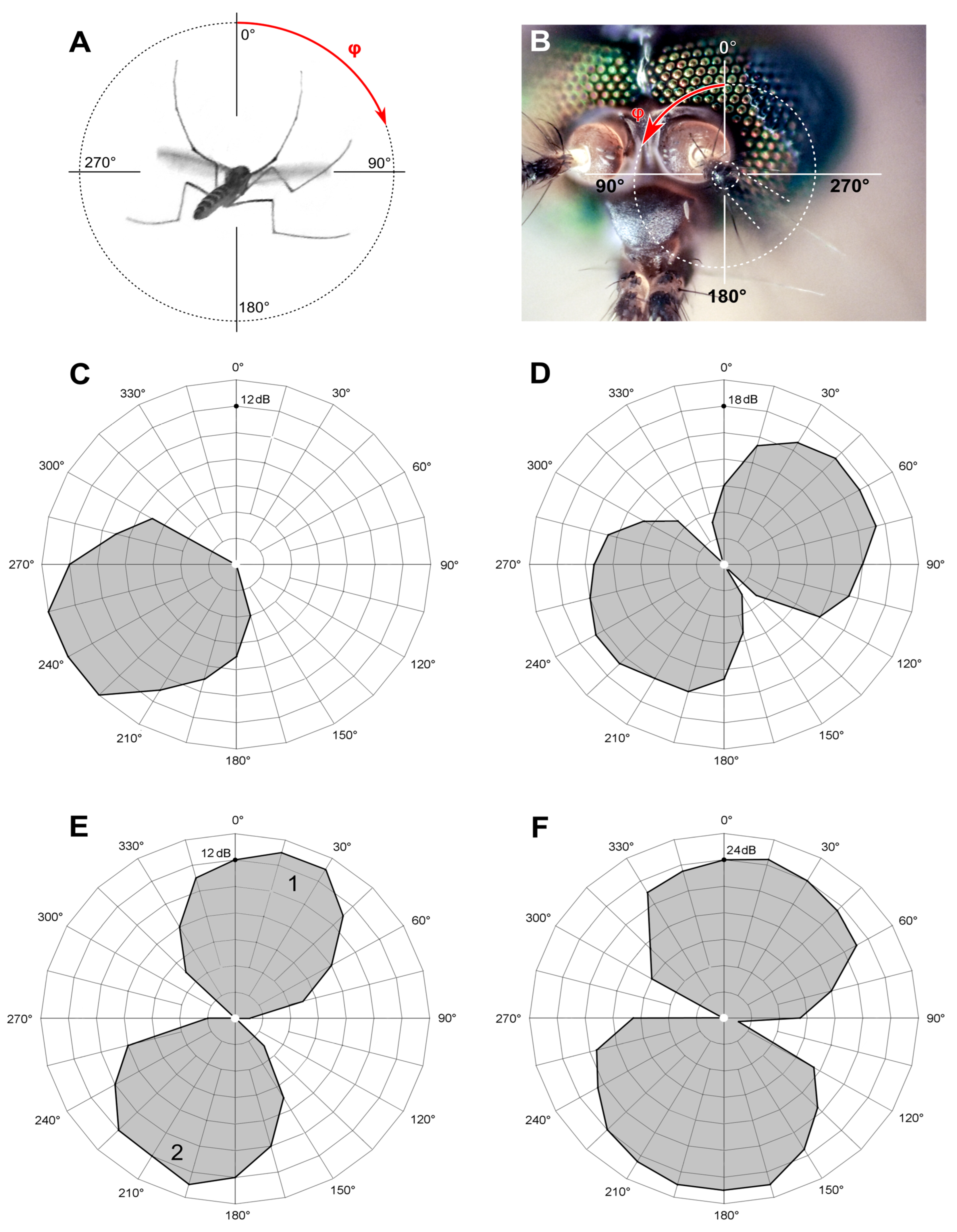
Experiments were conducted in laboratory conditions with air temperature 17–24 °C. Focal extracellular recordings from the axons of the antennal nerve were made with glass microelectrodes (1B100F–4, WPI Inc., Sarasota, FL, USA) filled with 0.15 M sodium chloride and inserted at the scape–pedicel joint (
Figure 2A). After the penetration of the cuticle, the electrodes had a resistance of 10–40 MΩ.
While penetrating the antennal nerve by the electrode, the preparation was continuously stimulated with tonal pulses (amplitude 60 dB SVPL, duration 80 ms, period 600 ms and dorso-ventral direction of acoustic vector (0°)). During this searching procedure the groups of the JO neurons situated orthogonal to the antenna oscillation could be overlooked, so the vector of the acoustic wave was periodically changed by 90°. The searching frequency was 100–130 Hz in the main series of experiments and 70 Hz during the search for low-frequency-tuned units (see below). We considered the recording site acceptable when the amplitude of response increased above 0.5 mV (peak to peak,
Figure 2C).
In this study, a number of axons of primary sensory neurons (PSNs) contributed to the extracellular recording. Although we could not estimate this number in each recording, our previous studies [
31,
33], which included the intracellular recordings from the axons of PSNs, suggested that the number of simultaneously recorded axons was not large. In our experience, the stability of intracellular recording from the axon of the JO PSN is hard to maintain during the time required for the measurement of thresholds at different frequencies or directions of sound. For the sake of the stability of the recording, in this study we used only extracellular recording of neuronal responses. We use the terms ‘unit’ or ‘sensory unit’ in the sense of one or several axons belonging to the PSNs of the JO, closely located within the antennal nerve and sharing indistinguishable frequency and phasic properties, thus representing a single functional unit.
2.4. Acoustic Stimulation
We used two orthogonally oriented Scandinavia 75 (DLS, Örebro, Sweden) stationary speakers to create a vector superposition of acoustic waves at the point of mosquito antenna (
Figure 2A,B), as described in detail in [
13]. The mosquito was positioned at the crossing of the axes of two speakers in such a way that the antenna′s flagellum was perpendicular to the directions of sound waves originating from each of the two speakers (
Figure 2B). This approach enabled us to set the desired direction of the acoustic vector relative to the antenna flagellum.
The moving parts of the speakers had a low resonant frequency (90 Hz). Due to the considerable response lag of the dome of the speaker and its support, the emission phase delay increased with the signal frequency up to the point of inversion. To stabilize the phase delay, a phase correction depending on the stimulation frequency was included in the speaker control circuit. Both speakers were covered by a metallic mesh to screen the recording electrode from the electrical signals that drove the speakers.
The sinusoidal stimuli were generated by the digital-to-analog converter LA-DACn10m1 (Rudnev-Shilyaev, Moscow, Russian Federation). Acoustic calibration of the stimulating device was performed with an NR-231-58-000 differential capacitor microphone (Knowles Electronics, USA) attached to a micropositioner with an axial rotation feature and set in the position of the mosquito. The same microphone placed 2 cm from the mosquito was used to record the stimulation signals during the experiments.
The speakers were powered by the home-made amplifier via a passive Sin–Cos (SC) transducer which produced two derived signals with the amplitudes
where
A1 and
A2 are the amplitudes of the signals for the first and the second speaker, respectively; U is the alternating voltage at the input of the SC transducer; φ is the angle between the dorso-ventral axis passing through the mosquito′s head and the vector of the vibrational velocity of the air particles. An increase in φ corresponds to the clockwise rotation of the velocity vector when viewed from the mosquito′s head along the antenna.
The resulting direction of the air vibration velocity in the stimulating system was determined by the vector superposition of the signals from both speakers. Changes in the sound wave direction relative to the mosquito in 15° steps were accomplished by coordinated switching of voltage dividers in the SC transducer. For those angles at which the values of the functions sin(φ + 45) or cos(φ + 45) were negative, the signal polarity was inverted by switching the terminals of the speakers.
The differential microphone together with its amplifier was previously calibrated in the far field using a B&K 2253 sound level meter with a B&K 4135 microphone (Brüel & Kjær, Nærum, Denmark). All sound level data in this study are given in the logarithmic scale in dB RMS SPVL (root mean square sound particle velocity level), with a reference level of 0 dB being equal to 4.85 × 10−5 mm/s, which corresponds in the far field to the standard reference sound pressure of 20 µPa.
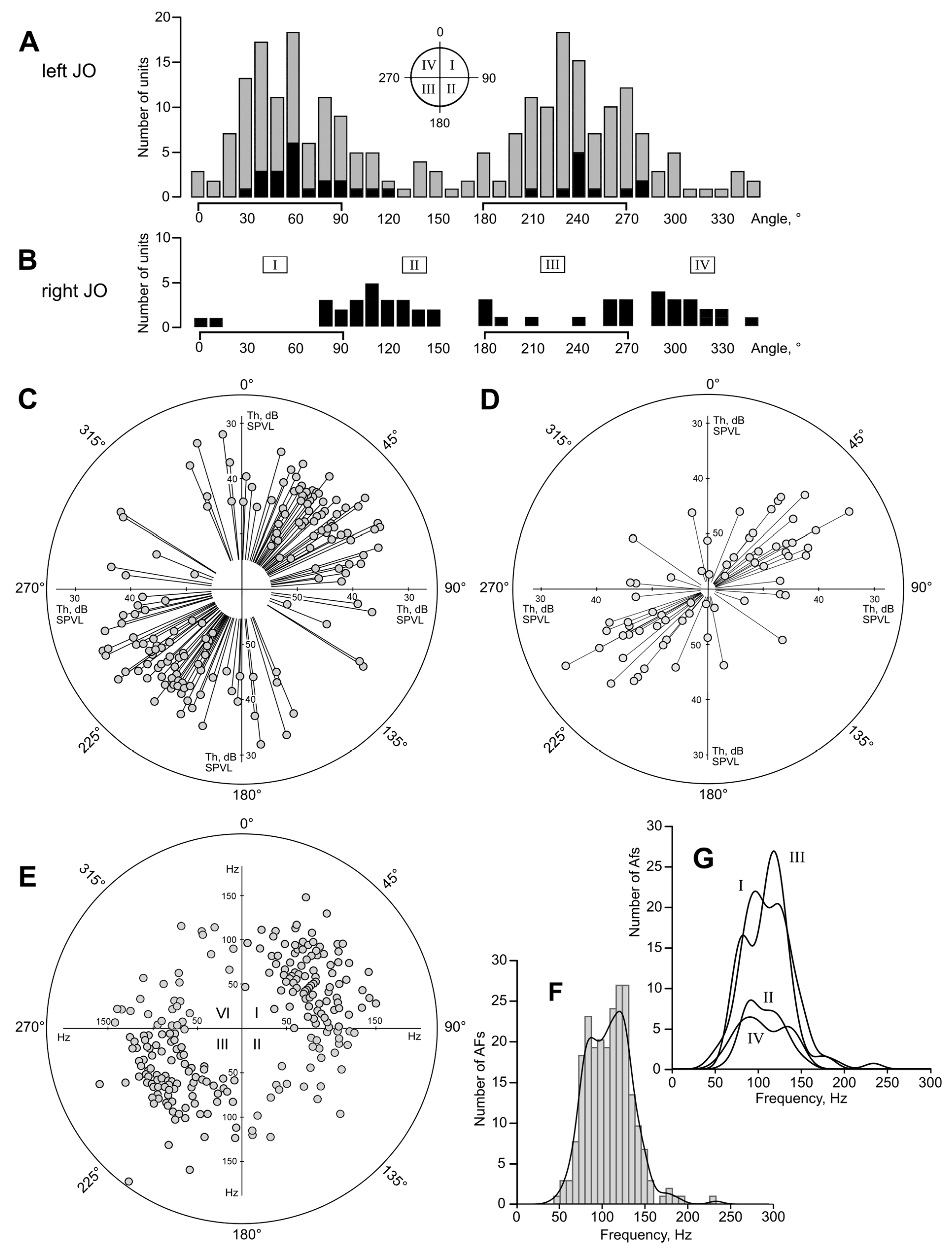
The stereotyped way of positioning the mosquito relative to the experimental setup, especially to the recording electrode, could lead to selective recording only from a certain part of the antennal nerve. To minimize the probability of such a bias, the mosquito was occasionally positioned ventral side up (180° rotation, n = 15) or rotated by ±45° (n = 16) or ±90° (n = 37) around the longitudinal axis of its body. In each case, the recording electrode was inserted through a different part of the scape–pedicel joint. The directional data measured in all such experiments were corrected accordingly. To make an additional control for the above-mentioned experimental bias, in a separate series of experiments (n = 24) we performed recordings from the right JO instead of the left one, keeping similar all other properties of the experimental setup. To avoid confusion due to the possible symmetrical nature of the two antennae, unless specifically stated, the presented data include measurements from only the left JO, while the data from the right JO serve only as a control.
2.5. Positive Feedback Stimulation
The essence of the positive feedback stimulation is a feedback loop established using the amplified in-phase response of a sensory unit as the signal to drive the stimulation loudspeaker. Applying such a stimulation to the sensory unit, we expect it to ‘sing’ at the frequency which is close to its intrinsic tuning frequency—an effect hereafter called ‘autoexcitation’. According to our view, several differently tuned PSNs near the electrode tip compete to set the autoexcitation frequency (AF). When one of them (in some cases two or, rarely, three) puts the system into the autoexcitation, other sensory units with different tuning frequencies become inhibited due to the general level adaptation of the JO. Many similarly tuned units oriented in line with the stimulation sound wave may be active during the autoexcitation, but the ability to determine the AF must be extremely sensitive to the amplitude of a unit’s recorded response, which in turn depends on the distance to the recording electrode.
The value of the AF was inevitably affected by the total phase shift in the feedback loop, arising primarily from the latency of the sensory cell response. As the phase shift was not constant along the frequency range, for each frequency measurement, a compensatory phase shift was introduced into the loop. The value of that phase shift was adjusted to provide the maximal amplitude and stability of autoexcitation at the same level of feedback.
The stimulation signal passed through a complex frequency filter to compensate for the influence of the loudspeaker mechanical resonance at lower frequencies. Additionally, the signal was low-pass-filtered with a slope of −6 dB per octave above 500 Hz to reduce the second and higher harmonic components of the sensory unit’s extracellular response. The stimulation signal was limited within an amplitude of 80 dB SPVL to prevent its uncontrolled rise due to positive feedback.
The complete feedback loop included the following elements: the JO sensory unit and its axon in the antennal nerve, recording microelectrode, amplifier, stimulation amplitude adjuster, frequency filter, phase adjuster, amplitude limiter, power amplifier, loudspeakers and, closing the loop, flagellum of a mosquito’s antenna that passed vibrations to the JO.
The method of positive feedback stimulation may be considered a functional sharpening of the electrode, allowing for the recording of only a single or a few axons of the PSNs lying in immediate proximity of the electrode tip. To our knowledge, this method currently allows the best spatial resolution of recording from multiple tightly packed sensory neurons.
Before making any tests of a unit’s directionality, we ensured the quality of recording according to the following criteria: low levels of feedback required to start the autoexcitation and the presence of not more than three different units responding in a narrow angular range.
Positive feedback stimulation allows us to separate the antiphase responses of the axons of sensory units which are, in most cases, recorded together [
13,
30,
32] and are presumed to lie close to each other in the antennal nerve. Contrary to this, other kinds of acoustic stimulation (e.g., sinusoidal stimulation) do not allow such a separation, as each of the units respond to both phases of the acoustic vibration. On the other hand, feedback stimulation does not allow us to measure the true auditory threshold of a sensory unit, providing only a relative estimate of sensitivity. Thus, for each recording site, we used two kinds of acoustic stimulation: positive feedback and sinusoidal.
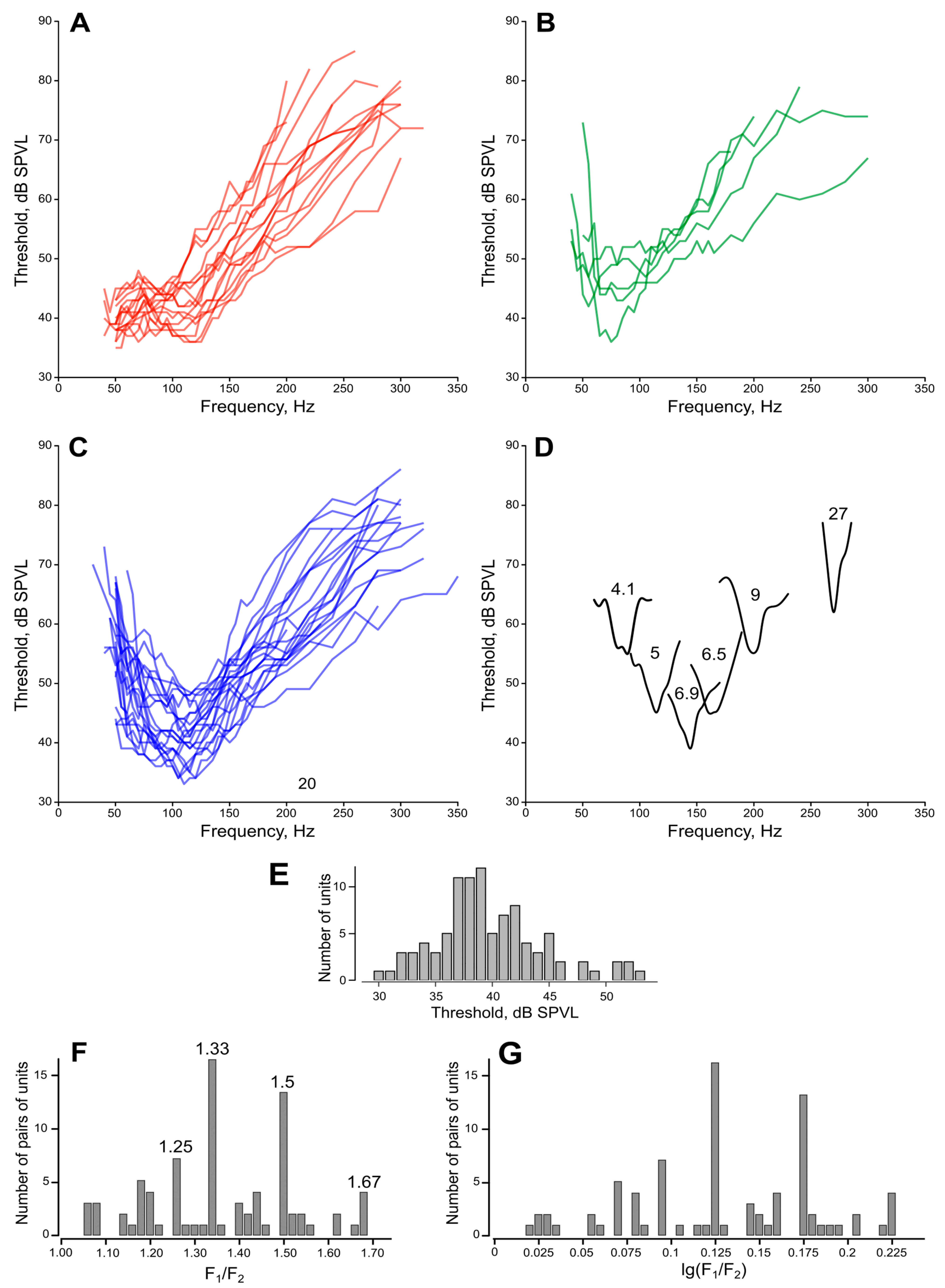
2.6. Measurements of Directionality
The successive stages of the electrophysiological experiment are summarized in
Figure 3.
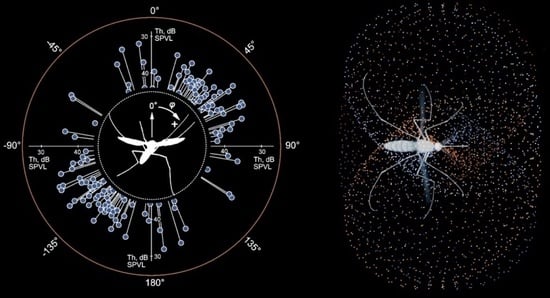
Two kinds of directional measurements were performed, depending on the type of stimulation. (1) The thresholds of the feedback stimulation were measured depending on the direction of a sound wave, producing an unipolar plot (hereafter ‘polar pattern’, example in
Figure 4C). (2) Using the measured AF value as the frequency of sinusoidal stimulation, the absolute auditory thresholds were measured depending on the direction of a sound wave, producing a bipolar plot (hereafter ‘directional characteristics’, example in
Figure 4D).
In 162 mosquitoes studied, of which in 24 the auditory responses were recorded from the right JO, a total of 290 (50 from the right JO) polar patters were obtained (
Table 1).
In the mode of feedback stimulation, we could only measure the relative threshold of auto-excitation for the entire system, including the mosquito and the stimulation setup. Such a relative threshold was defined as the signal level, which required one more incremental step (+1 dB) at the attenuator output for the system to enter a state of sustained autoexcitation (
Figure 2D). For each AF, we measured a polar pattern (relative threshold of autoexcitation as a function of orientation of the acoustic wave vector relative to the axis of the antenna, with a 15° step). Then, using sinusoidal stimulation at AF (rounded to 5 or 10 Hz), a directional characteristic (similar function for auditory threshold).
The criterion of the response threshold was set at 2 dB of sustained excess of the response amplitude above the average noise level in a given recording. At each combination of stimulation parameters, the threshold was measured consecutively at least twice. Such auditory thresholds, measured in response to sinusoidal stimulation and expressed in dB RMS SPVL, are hereafter called ‘absolute thresholds’ or just ‘thresholds’.
Similarly, after finding the best direction using the positive feedback stimulation, we measured a frequency-threshold curve in the range of 40−300 Hz using sinusoidal stimulation with a 5 Hz step below 170 Hz and a 10 Hz step otherwise. Below 40 Hz, the measurements were limited by a substantial decrease in the efficiency of the speakers.
The best frequency and best direction of a given unit, obtained by means of measuring the auditory thresholds, also served as a control check for the AF and best direction, obtained through a positive feedback stimulation.
2.7. Search for Low-Frequency Units
Based on the preliminary results, we realized that the true responses of the low-frequency sensory units are often difficult to distinguish from the combination harmonics that appear when two or more high-frequency units respond simultaneously at different frequencies (example of two responses in
Figure 2D, traces at 69 and 114 Hz). This effect could lead to the under-estimation of true low-frequency-tuned units. Moreover, the positive feedback stimulation method was found to be rather ineffective at the lower frequencies, most probably due to the high competition from the more sensitive units tuned to higher frequencies, which often captured the autoexcitation frequency. Accordingly, we designed an additional series of experiments to compensate for these biases. In these experiments, the searching frequency was set to 70 Hz, and during the choice of the recording site we gave preference to the units that responded maximally at that or at lower frequencies. Moreover, the initial (searching) vector of the acoustic wave was set to φ = –45°, that is, to search for units in the II and IV quadrants, where, according to the preliminary analysis of the data, we expected to find less responding units.
2.8. Data Analysis
Directional plots. In an array of threshold data obtained from a single recording site, we determined the maximum threshold value (Th
max). Based on this, a set of derived values describing each unit’s directional characteristic or polar pattern was estimated using the equation
where Th
i is the threshold at any given direction of the sound wave. In the plots based on these data, the sectors of the highest sensitivity corresponded to the lowest recorded thresholds, and the central zero point corresponded to Th
max. The angles at which no response at the best frequency was observed were given the value A
i = 0.
The angular sensitivity range of a unit (the width of its directional characteristic) was determined at −6 dB of its maximum sensitivity (in case of bipolar directional characteristics the values from the two independently measured symmetrical plots were averaged). The best direction of a given unit was determined as the bisector of this range.
Sonograms, from which the AF data were extracted, were plotted and analyzed in Sound Forge Pro 10 (Sony).
Correction for temperature. As the experiments were performed in a range of ambient temperatures, all frequency data were normalized to 20 °C according to the previously established equation:
where t is a temperature for a given experiment and k = 0.02 is a relative shift of tuning frequency with a change of frequency by 1 °C [
34].
Statistics. Statistical analysis was performed in PAST [
35] and R [
36]. Measurements are given as mean ± standard deviation, with the distributions pre-tested for normality using the Shapiro–Wilk test.
5. Conclusions
According to our findings, female mosquitoes possess asymmetric distribution of auditory neurons within the Johnston’s organ, mirrored in the left and right. The neuronal asymmetry is coordinated with the in-flight position of antennae to maintain a uniformly shaped auditory space, symmetric relative to a mosquito body. This finding may reflect a common principle of directional hearing based on the particle velocity receivers, which in its turn may determine the ranges of angular position of antennae in different insects.
Within the mosquito auditory space, there are areas located above, below, and in front of a flying mosquito, which we called equisignal zones. Within these zones, directional cues should not depend on the absolute level of sound, and, accordingly, on the distance to the source. This feature may be critically important during swarming behavior, when the everchanging distance to other mosquitoes demands instant and precise directional estimates.
At the same time, each JO possesses units that duplicate the function of the contralateral one. Such redundancy increases the reliability of the entire system and potentially can provide acoustic orientation even if one antenna gets damaged.
The auditory system of female mosquitoes has enough resolution to estimate the direction to the sound source, while its frequency range enables detection of sounds produced by the animal and human hosts.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}