Simple Summary
In Drosophila melanogaster, autophagy regulates development and stress responses. However, the mechanisms underlying how autophagy modulates innate immunity still need further investigation. In this study, we identified Atg2, an autophagy-related protein, as playing an important role in controlling innate immunity. We revealed that inhibiting Atg2 induced melanotic nodule formation, disrupted phagocytosis in vivo, altered the expression of AMP-encoding genes and impaired the ability to resist bacterial infections. This study will advance the understanding of the relationship between autophagy and innate immunity.
Abstract
Autophagy is a process that promotes the lysosomal degradation of cytoplasmic proteins and is highly conserved in eukaryotic organisms. Autophagy maintains homeostasis in organisms and regulates multiple developmental processes, and autophagy disruption is related to human diseases. However, the functional roles of autophagy in mediating innate immune responses are largely unknown. In this study, we sought to understand how Atg2, an autophagy-related gene, functions in the innate immunity of Drosophila melanogaster. The results showed that a large number of melanotic nodules were produced upon inhibition of Atg2. In addition, inhibiting Atg2 suppressed the phagocytosis of latex beads, Staphylococcus aureus and Escherichia coli; the proportion of Nimrod C1 (one of the phagocytosis receptors)-positive hemocytes also decreased. Moreover, inhibiting Atg2 altered actin cytoskeleton patterns, showing longer filopodia but with decreased numbers of filopodia. The expression of AMP-encoding genes was altered by inhibiting Atg2. Drosomycin was upregulated, and the transcript levels of Attacin-A, Diptericin and Metchnikowin were decreased. Finally, the above alterations caused by the inhibition of Atg2 prevented flies from resisting invading pathogens, showing that flies with low expression of Atg2 were highly susceptible to Staphylococcus aureus and Erwinia carotovora carotovora 15 infections. In conclusion, Atg2 regulated both cellular and humoral innate immunity in Drosophila. We have identified Atg2 as a crucial regulator in mediating the homeostasis of immunity, which further established the interactions between autophagy and innate immunity.
1. Introduction
Despite the lack of adaptive immunity in Drosophila melanogaster, its innate immune system, acting as the first line of defense, shares similarities with mammals both in physiology and signaling pathways [1]. Thus, in recent decades, Drosophila has emerged as an ideal model for studying innate immunity [2]. The innate immune response of Drosophila is mainly composed of two parts: the cellular immune response and the humoral immune response [1,2]. The cellular immune response is executed by three types of hemocytes (blood cells of Drosophila), in which plasmatocytes constitute 95% of the total hemocytes and play an important role in phagocytosis [3,4,5]. Macrophage-like plasmatocytes are capable of engulfing pathogens and apoptotic debris produced during development [3,4,5]; the dynamics of the actin cytoskeleton and multiple phagocytosis receptors, including Nimrod C1 (NimC1), Eater, Draper, Croquemort, Down syndrome cell adhesion molecule 1 (Dscam1) and class C scavenger receptor I (dSr-CI), are required in this process [6,7,8]. Crystal cells, another type of hemocyte, can heal wounds via melanization responses [1,2]. In addition, although rare in healthy larvae, lamellocytes are largely induced upon immune challenge and are in charge of encapsulating large foreign bodies that cannot be phagocytosed [9,10]. As another aspect of innate immunity, the humoral immune response largely depends on antimicrobial peptides (AMPs). When encountered with immune challenge, fat bodies will secrete AMPs into the hemolymph to combat invading pathogens [1,2]. In Drosophila, the induction of AMPs is mainly regulated by two NF-κB-related signaling pathways, Toll and Immune deficiency (Imd). Moreover, the Toll pathway shows higher affinity for fungi and Gram-positive bacteria, whereas the Imd pathway is more likely to be involved in the response to Gram-negative bacteria [1,2].
Autophagy is a conserved process that promotes the intracellular degradation of cytoplasmic proteins or organelles with lysosomes in eukaryotic organisms [11,12]. Under normal conditions, the activity of autophagy is maintained at a relatively low level [13]. However, upon starvation or other stresses, autophagy is strongly induced [14,15]. Thus, autophagy plays a critical role in maintaining the homeostasis of cells and organisms, and autophagy has been shown to be versatile in regulating development and the stress response [16,17,18]. In addition, autophagy dysfunction is associated with human diseases, including neurodegeneration and cancer [19]. In Drosophila, autophagy activity is modulated by a series of autophagy-related (Atg) genes, which are highly conserved from yeast to mammals [20,21]. The biological functions of some Atg proteins are relatively well characterized; for instance, Atg1 regulates cell growth and apoptotic cell death and maintains germline stem cells by modulating mitochondrial dynamics [22,23]. Inhibiting Atg5 induces developmental delay and ataxia, while Atg8 is involved in larval midgut cell programmed cell death and determination of adult lifespan [24,25,26]. However, the functional role of Atg2 is largely unknown. Moreover, although autophagy was shown to restrict bacterial infection and viral replication, the detailed mechanism still warrants further investigation.
In this study, we focused on Atg2, an Atg protein with a lesser-known biological function, and its role in regulating innate immunity. We showed that inhibiting Atg2 in hemocytes induced the formation of melanotic nodules and disrupted phagocytosis. In addition, the proportion of NimC1-positive hemocytes decreased, and hemocytes displayed altered actin cytoskeleton distribution upon inhibition of Atg2. Flies with low Atg2 expression showed altered AMP-encoding gene expression and succumbed to bacterial infection. In summary, we identified Atg2 as a crucial regulator of innate immunity, which established the connection between autophagy and innate immune responses.
2. Materials and Methods
2.1. Drosophila Strains and Culture Conditions
w1118 (BL5905, used as the wild type), Atg2EP3697/TM6B, Tb (BL17156) and Cg-Gal4 (BL7011) were obtained from the Bloomington Drosophila Stock Center (BDSC). UAS-Atg2RNAi (THU3698) was obtained from the TsingHua Fly Center. UAS-ImdRNAi (V9253) were obtained from the Vienna Drosophila Resource Center. Pxn-Gal4; UAS-GFP was a gift from Professor Norbert Perrimon. UAS-DifRNAi was a gift from Professor Lei Xue. The flies were fed standard cornmeal-yeast medium and maintained at 25 °C in an incubator. To activate the UAS/Gal4 system, the conditions were shifted to 29 °C for at least 48 h.
2.2. In Vivo Phagocytosis
Ten to fifteen wandering 3rd instar larvae from each group were injected with 200 nl of latex beads (10%, F8821 and F8823, Invitrogen, Waltham, MA, USA), pHrodo-Escherichia coli (E. coli) (1 mg/mL, P35361, Thermo Fisher Scientific, Waltham, MA, USA) or pHrodo-Staphylococcus aureus (S. aureus) (1 mg/mL, A10010, Thermo Scientific) using an injector (PICOSPRITZER III, Parker Hannifin, Mayfield Heights, OH, USA). Next, injected larvae were placed in a humid chamber at room temperature for 1 h. To obtain circulating hemocytes, the larval cuticle was ripped gently with fine forceps (Dumont, Montignez, Switzerland) when submerging larvae in 10 μL of phosphate-buffered saline (PBS) and then incubated in a dark chamber at room temperature for 30 min on an adhesive glass slide. After fixation with 4% paraformaldehyde (PFA) for 10 min and several careful washes with PBS containing 0.1% Tween 20 (PBST), hemocytes were mounted with 70% glycerol and observed under a fluorescence microscope (Axio Scope A1, Zeiss, Germany). The images were captured randomly to avoid bias. To calculate the average phagocytosed particles per hemocyte (phagocytic index), the total number of particles (or the total fluorescence intensities) and the number of hemocytes of each image were determined with ImageJ software. Phagocytic index = Total number of particles (or total fluorescence intensities)/The total number of hemocytes. Engulfing cell (%) = The number of hemocytes with phagocytosed particles/The total number of hemocytes. Eight larvae from each group were employed in the bleeding. For each group, at least 500 hemocytes were analyzed.
2.3. Immunostaining of Melanotic Nodules
Melanotic nodules were directly dissected from wandering 3rd instar larvae and fixed with 4% PFA for 30 min. Following 3 washes (5 min/wash) with PBST, melanotic nodules were blocked with PBST containing 5% goat serum for 30 min and incubated with anti-L1 antibodies (a gift from Professor István Andó) at 4 °C overnight. After several rinses in PBST, melanotic nodules were incubated with Alexa Fluor 568-conjugated secondary antibodies (A-21124, Thermo Fisher Scientific). Following washing with PBST 3 times, dissected tissues were stained with Hoechst dye (H1399, Invitrogen) to visualize cell nuclei, mounted with Slowfade (S36967, Invitrogen), and observed under a microscope (Axio Scope A1, Zeiss, Germany). For each group, at least 15 larvae were analyzed.
2.4. Hemocyte Assays
P1 staining: eight wandering 3rd instar larvae from each group were bled in 10 μL of PBS, and then hemocytes were transferred to an adhesive glass slide. After incubation at room temperature for 30 min, hemocytes were fixed with 4% PFA, blocked with PBST containing 5% goat serum and incubated with anti-P1 (a gift from Professor István Andó) at 4 °C overnight. The next day, hemocytes were incubated with Alexa Fluor 568-conjugated secondary antibodies (A-11004, Invitrogen) and Hoechst dye for 2 h and 10 min, respectively, and mounted with Slowfade.
F-actin staining: eight wandering 3rd instar larvae were bled in 10 μL of PBS, incubated at room temperature for 30 min, and fixed with 4% PFA for 10 min. After 3 washes, hemocytes were incubated with Alexa Fluor 568-conjugated phalloidin (A12380, Invitrogen) and Hoechst dye for 30 min and 10 min, respectively. Then, hemocytes were mounted with Slowfade.
TUNEL and 7-AAD staining: To examine cell apoptosis and death, hemocytes were incubated with a TUNEL kit (11684795910, Roche, Basel, Switzerland) and 7-AAD (5 μg/mL, A1310, Invitrogen), respectively, according to the manufacturer’s instructions.
All stained sections were analyzed under a fluorescence microscope (Axio Scope A1, Zeiss, Germany).
2.5. Quantitative Real-Time PCR (qRT—PCR)
Eight adult flies from each group were collected in TRIzol reagent (Invitrogen) and used to isolate total RNA. After synthesizing cDNA using reverse transcriptase (Promega), qRT—PCR was performed with SYBR Green I Master Mix (Roche) on a Roche LightCycler 480 real-time PCR system. The transcript levels of AMP-encoding genes were normalized to ribosomal protein 49 (rp49), the primer sequences used are shown in Table 1. The fold change was calculated with the 2−∆∆Ct method [27]. The experiments were executed with three independent replications.

Table 1.
Primer sequences used qRT—PCR.
2.6. Survival Rates upon Bacterial Infection
Three- to five-day-old adult flies (15 males and 15 females/group) were injected with 50 nl of S. aureus (OD = 0.0005) or Erwinia carotovora carotovora 15 (Ecc-15, OD = 0.4). After injection, flies were incubated at 29 °C, and surviving flies were counted every 24 h. In addition, injected flies were shifted to a new vial every 48 h. The experiment was repeated independently three times.
2.7. Statistical Analysis
All microscopic images were analyzed with ImageJ software. The statistical analysis was performed with GraphPad Prism 6.0, and p values were determined with unpaired Student’s t test or log-rank (Mantel—Cox) test (survival assay). The thresholds for statistical significance were * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001. All experiments were repeated independently at least three times. The error bars in all column diagrams indicate the means ± SDs.
3. Results
3.1. Inhibiting Atg2 Induced Massive Melanotic Nodule Formation
To examine the functional role of Atg2 in the innate immunity of Drosophila, we first knocked down Atg2 in the blood system with the hemocyte-specific Gal4 driver Pxn-Gal4; UAS-GFP. Surprisingly, massive melanotic nodules emerged in Pxn > GFP/Atg2RNAi larvae, while no melanotic nodules were observed in the control group (Figure 1A,B). To confirm this result, we utilized Cg-Gal4, an immune system-specific driver (hemocytes and fat bodies), to suppress Atg2 and observed a large number of melanotic nodules in Cg > Atg2RNAi larvae (Figure 1C,D). Consistent with the above data, loss of Atg2 by using mutant Atg2EP3697/Tb induced melanotic nodule formation (Figure 1E,F). Moreover, after immunostaining with anti-L1 antibodies that label lamellocytes, melanotic nodules showed GFP-positive (signals from Pxn-Gal4; UAS-GFP) and L1-positive signals, indicating that plasmatocytes and lamellocytes were deposited on the melanotic nodules (Figure 1G–K). To exclude non-specific fluorescence signals during the immunostaining, we stained melanotic nodules without using anti-L1 antibodies but using Alexa Fluor-conjugated secondary antibodies; no red fluorescence was observed, indicating that the fluorescence signals were specific (Figure S1A–E). However, TUNEL assay and 7-AAD staining showed that no significant cell apoptosis or cell death occurred in circulating hemocytes of Pxn > GFP/Atg2RNAi larvae (Figure S2A–F). Collectively, inhibition of Atg2 significantly induced melanotic nodule formation.
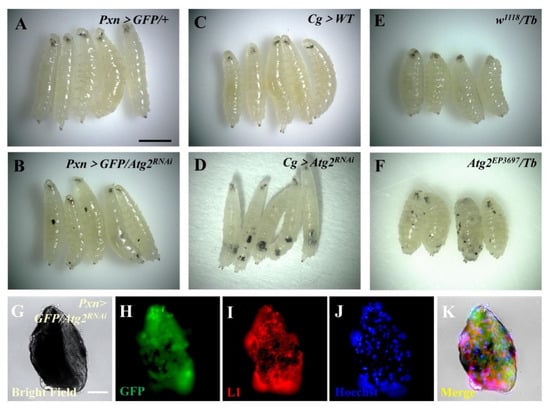
Figure 1.
Inhibiting Atg2 led to massive melanotic nodule formation. (A–F) The formation of melanotic nodules was observed in Pxn > GFP/Atg2RNAi, Cg > Atg2RNAi and Atg2EP3697/Tb larvae. (G–K) Immunostaining showed that plasmatocytes (GFP-positive cells, green) and lamellocytes (L1-positive cells, red) emerged in melanotic nodules from Pxn > GFP/Atg2RNAi larvae. Scale bars: 500 μm (larvae) and 50 μm (melanotic nodules).
3.2. Inhibiting Atg2 Impaired In Vivo Phagocytosis
Next, we sought to determine the effects of inhibiting Atg2 on hemocyte function by examining in vivo phagocytosis. First, latex beads, nonpathogenic particles with red fluorescence, were incorporated, and the results showed that compared with the control group, the proportion of hemocytes with phagocytosed beads (engulfing hemocytes) significantly decreased in Pxn > GFP/Atg2RNAi hemocytes; the average number of phagocytosed beads per hemocyte (phagocytic index) also decreased (Figure 2A–D). In addition, the phagocytosis of the Gram-positive bacterium S. aureus was disrupted upon inhibition of Atg2 (Figure 2E–H). Given that different bacteria might activate different phagocytosis receptors, the Gram-negative bacterium E. coli was utilized. Similarly, the proportion of hemocytes with phagocytosed pHrodo-E.coli and the phagocytic index decreased in Pxn > GFP/Atg2RNAi, showing that in vivo phagocytosis was disrupted by inhibiting Atg2 (Figure 2I–L). Moreover, these results were confirmed in Cg > Atg2RNAi and Atg2EP3697/Tb larvae (Figure S3), indicating that Atg2 facilitates in vivo phagocytosis.
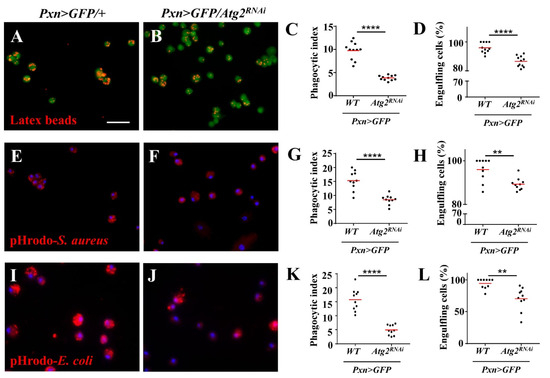
Figure 2.
Knocking down Atg2 in hemocytes disrupted the phagocytosis of latex beads and bacteria. (A–L) Pxn > GFP/+ and Pxn > GFP/Atg2RNAi larvae were injected with latex beads (A,B), pHrodo-S. aureus (E,F) and pHrodo-E. coli (I,J). The quantification analysis showed that the average phagocytosed particles per hemocyte (phagocytic index) (C,G,K) and the percentage of engulfing hemocytes (hemocytes with phagocytosed particles) (D,H,L) significantly decreased when Atg2 was knocked down in hemocytes. ** p < 0.01, **** p < 0.0001; scale bar: 25 μm.
3.3. Inhibiting Atg2 Altered the Proportion of NimC1-Positive Hemocytes and the Actin Cytoskeleton
To further explore the mechanism of impaired phagocytosis upon inhibition of Atg2, we examined the expression of NimC1, a phagocytosis receptor and a marker for mature plasmatocytes, with anti-P1 antibodies. A total of 95.14% of the hemocytes from the control group were P1-positive, while only 41.16% of hemocytes were labeled with P1 in Pxn > GFP/Atg2RNAi hemocytes, indicating that inhibiting Atg2 decreased the proportion of NimC1-positive hemocytes (Figure 3A,B); this was also observed in Cg > Atg2RNAi and Atg2EP3697/Tb hemocytes upon latex bead injection (Figure 3D–F,H–J). In addition, we compared the phagocytosis capacity between P1-positive and P1-negative hemocytes and showed that P1-positive hemocytes significantly phagocytosed more latex beads (Figure 3G,K). We stained hemocytes from w1118 larvae (with or without being injected with latex beads) only with Alexa Fluor 568-conjugated secondary antibodies, and no red fluorescence was observed (Figure S1F,G); this result suggested that the red fluorescence in Figure 3 was specific.
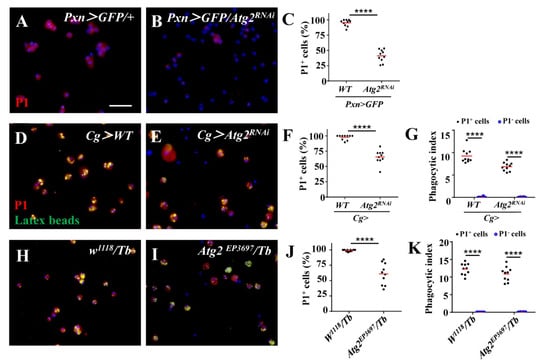
Figure 3.
The proportion of NimC1-positive hemocytes decreased upon inhibition of Atg2. (A–C) The proportion of NimC1-positive hemocytes displayed by P1 staining significantly decreased in Pxn > GFP/Atg2RNAi hemocytes. (D–K) When injected with latex beads (green), the percentage of P1-positive hemocytes also decreased in Cg > Atg2RNAi and Atg2EP3697/Tb larvae. In addition, P1-positive hemocytes phagocytosed more latex beads than P1-negative hemocytes. **** p < 0.0001; scale bar: 25 μm.
Next, we labeled the F-actin of hemocytes with phalloidin to examine whether inhibiting Atg2 affected the actin cytoskeleton. Although the average length of filopodia was longer in Pxn > GFP/Atg2RNAi hemocytes than in the control group, the average number of filopodia decreased (Figure 4A–D); this result suggested that hemocytes displayed aberrant F-actin distribution. We confirmed this result in Cg > Atg2RNAi and Atg2EP3697/Tb larvae (Figure 4E–L). In summary, Atg2 plays an important role in maintaining normal proportion of NimC1-positive hemocytes and the actin cytoskeleton.
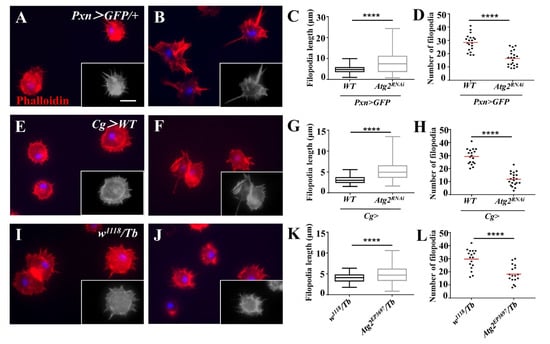
Figure 4.
Inhibiting Atg2 affected actin cytoskeleton distribution. (A–L) The hemocytes from Pxn > GFP/+, Pxn > GFP/Atg2RNAi, Cg > WT, Cg > Atg2RNAi, w1118/Tb and Atg2EP3697/Tb larvae were stained with phalloidin. The average filopodia length and filopodia number are shown in (C,D,G,H,K,L). To better identify filopodia, the grayscale images are also provided. The data showed that the hemocytes showed aberrant actin cytoskeleton distribution upon inhibition of Atg2. **** p < 0.0001; scale bar: 10 μm.
3.4. Inhibiting Atg2 Altered AMP-Encoding Gene Expression
The above data showed that Atg2 participated in cellular immune responses. Thus, we sought to determine whether Atg2 also functions in humoral immune responses, another arm of Drosophila innate immunity. To verify this, we knocked down Atg2 in the immune system with Cg-Gal4 and checked the expression of a series of AMP-encoding genes, including Drosomycin (Drs), Attacin-A (AttA), Diptericin (Dpt), Metchnikowin (Mtk) and Defensin (Def) in larvae. Compared with the control group, the transcript level of Drs increased in Cg > Atg2RNAi larvae, whereas the expressions of Atta, Dpt and Mtk significantly decreased (Figure 5). This result showed that Atg2 had a role in maintaining the homeostasis of AMP production.
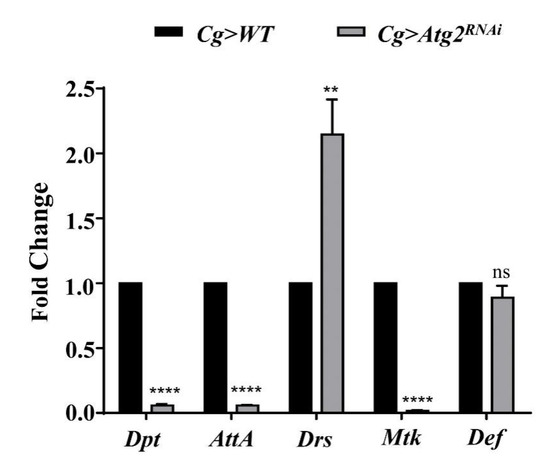
Figure 5.
Inhibiting Atg2 altered the expression of AMP-encoding genes. The transcript levels of AMP-encoding genes, including Dpt, AttA, Drs, Mtk and Def were determined by RT—qPCR, and the fold change was compared with the control group. The expression of Drs increased in Cg > Atg2RNAi larvae, while Dpt, AttA and Mtk were downregulated. ** p < 0.01, **** p < 0.0001, ns: not significant.
3.5. Flies Were Susceptible to Bacterial Infection upon Inhibition of Atg2
Finally, to examine whether altered innate immune responses caused by inhibiting Atg2 would affect the ability to resist infection, adult flies were septic-infected with S. aureus, and survival rates were determined; flies with decreased expression of Dif, a key regulator of the Toll pathway, were employed as the positive control. After infection for 7 days, 40.00% of control flies survived, while the survival rate decreased significantly in Pxn > GFP/Atg2RNAi and Pxn > GFP/DifRNAi flies, showing that only 8.31 and 1.90% flies survived, respectively (Figure 6A). Consistent with this, Cg > Atg2RNAi and Cg > DifRNAi flies succumbed to S. aureus infection (Figure 6B). In addition, Ecc-15 was incorporated to infect flies, and flies upon inhibition of Imd were used as the positive control. Similar to S. aureus infection, flies showed significantly lower survival rates in the Pxn > GFP/Atg2RNAi, Pxn > GFP/ImdRNAi, Cg > Atg2RNAi and Cg > ImdRNAi groups (Figure 6C,D). Collectively, these results indicated that Atg2 had a critical role in resisting bacterial infection.
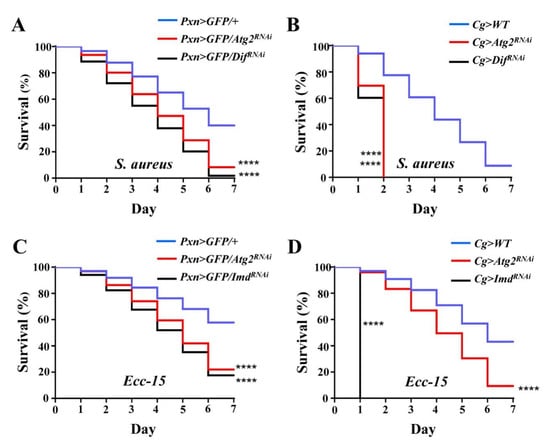
Figure 6.
Flies are more susceptible to bacterial infection upon inhibition of Atg2. (A–D) Upon S. aureus and Ecc-15 infections, Pxn > GFP > Atg2RNAi and Cg > Atg2RNAi flies showed significantly lower survival rates. Pxn > GFP/DifRNAi, Cg > DifRNAi, Pxn > GFP/ImdRNAi and Cg > ImdRNAi flies served as the positive controls and succumbed to S. aureus and Ecc-15 infections. For each group, 15 males and 15 females were employed in the survival assay, and the experiment was repeated three times. The cumulative values of three replications are shown in the figures (A–D). **** p < 0.0001.
4. Discussion
Autophagy is a conserved and crucial intracellular process that maintains the homeostasis of organisms. Several studies have reported the role of autophagy in regulating immunity [28,29,30]. Upon binding to the peptidoglycan recognition receptor PGRP-LE, the peptidoglycan of Listeria monocytogenes (L. monocytogenes) induces autophagy, and the bacteria are eliminated through Atg8 [28]. The clearance of Mycobacterium marinum and Salmonella enterica also involves Atg8 [28]. In addition, autophagy was shown to play an important role in restricting the vesicular stomatitis virus [28,30]. However, pathogens can also take advantage of the functions or components of autophagy to enhance their replication and maintain intracellular survival, indicating that the relationship between autophagy and immunity is complicated and that different Atg proteins may have different roles [31].
Unlike Atg1, Atg5 or Atg8, the function of Atg2 is not well characterized. A previous study showed that Atg2 is related to lipid transport function [32]. In this study, we found that inhibiting Atg2 induced a large number of melanotic nodules (Figure 1A–F). In fact, melanotic nodules have been studied for more than half a century and are regarded as a model of cancer [10]. The Toll, JAK/STAT, JNK and Ras/EGFR pathways participate in the formation of melanotic nodules [33,34,35]. The mechanism of melanotic nodule formation is complex, one of which involves the aggregation of hemocytes [36]. Consistent with this, plasmatocytes and lamellocytes were observed in melanotic nodules (Figure 1G–K). In addition, we observed more hemocytes in Pxn > GFP/Atg2RNAi larvae (Figure S2B,E). However, lamellocytes are rare in healthy larvae, indicating that hematopoiesis is aberrant upon inhibition of Atg2. We will investigate how Atg2 controls hematopoiesis in further studies. Notably, compared to Pxn > GFP/Atg2RNAi larvae, when Atg2 was knocked down using Cg-Gal4, a hemocyte- and fat-body-specific driver, more melanotic nodules were induced (Figure 1A–F), suggesting that an interaction between hemocytes and fat bodies may exist.
To examine whether the function of hemocytes was impaired, we used latex beads and pHrodo-conjugated pathogens to determine phagocytosis. pHrodo dye is sensitive to pH and will fluoresce in phagosomes whose pH is lower [37]. Based on this, we observed weaker fluorescent signals upon inhibition of Atg2, indicating that phagocytosis was suppressed (Figure 2). This finding is consistent with a previous study showing that Atg2 restricts the replication of Mycobacterium marinum, which involves the regulation of lipid droplets [38]. In our study, the proportion of NimC1 (P1)-positive hemocytes decreased upon inhibition of Atg2 (Figure 3). In addition, we also showed that P1-positive hemocytes phagocytosed more latex beads than P1-negative hemocytes. Given that NimC1 is a marker for mature hemocytes and a typical phagocytosis receptor, this result suggested that the proportion of immature hemocytes increased, which led to disrupted in vivo phagocytosis. Different receptors are in charge of different types of pathogens. Whether Atg2 affects other receptors warrants further study. Given that the phagocytosis of nonpathogenic particle latex beads, the Gram-negative bacterium E. coli and Gram-positive bacterium S. aureus was disrupted, Atg2 played a broad role in controlling phagocytosis. Studies showed that filopodia act as “tentacles” to capture particles during phagocytosis [39,40]. In this study, the filopodia were altered by inhibiting Atg2 (Figure 4). Similarly, Atg1 was shown to maintain F-actin protrusions of hemocytes, indicating that autophagy participates in actin cytoskeleton reorganization [41]. Moreover, our previous study revealed that Jumu regulates phagocytosis through NimC1 and cytoskeleton reorganization [42].
The secretion of AMPs by the fat body, equivalent to mammalian livers, is the hallmark of humoral immune responses of Drosophila [1,2]. This aspect of immunity is also known as the systemic immune response, which is efficient in eliminating invading pathogens. In addition, excessive AMP production damages cells [43]. In our study, the expression of AMP-encoding genes was altered by inhibiting Atg2 (Figure 5); Drs was upregulated, and the transcript levels of AttA, Dpt and Mtk were decreased, indicating that the humoral immune response was disrupted. AMPs were shown to regulate aging and long-term memory beyond the role in mediating immune responses [2]. Moreover, in addition to the Toll and Imd pathways, JNK pathway transcription factor AP-1 also controls the expression of AMP-encoding genes [44]. These results suggested that both the role and the regulation of AMPs are complex. Thus, we speculated that Atg2 may regulate different AMPs in different manners. In line with the altered humoral immunity, flies were more susceptible to bacterial infections upon inhibition of Atg2 (Figure 6). Although the contribution of cellular immune responses was questioned due to the prominent role of humoral immunity upon immune challenge, a previous study showed that plasmatocytes are required to promote AMP Def induction in the fat body during bacterial infections [45,46,47]. Consistent with this finding, knocking down Atg2 only in hemocytes significantly decreased the survival rates after septic infection, suggesting that Atg2 may participate in the interactions between cellular and humoral responses.
Previous studies have shown that PGRP-LE activates autophagy in response to L. monocytogenes, indicating that PGRP-LE acts as an upstream regulator of autophagy [28]. However, in our study, Atg2 was upstream of the phagocytosis receptor and controlled its expression. Thus, the relationship between autophagy and the innate immune response is rather complex and requires further study. In conclusion, we have identified Atg2 as an important regulator of immune responses, which will advance the understanding of how autophagy functions in innate immunity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14080706/s1, Figure S1: The negative controls for L1 staining and P1 staining. Figure S2: Inhibiting Atg2 did not induce cell apoptosis or death; Figure S3: Phagocytosis was disrupted upon inhibition of Atg2.
Author Contributions
Conceptualization, L.H.J. and B.Q.; software, Q.C.; validation, B.Q. and S.Y.; formal analysis, S.Y. and Q.C.; investigation, B.Q. and Q.C.; data curation, Q.C.; interpretation of data, S.Y.; writing—original draft preparation, S.Y.; writing—review and editing, B.Q., S.Y.; visualization, B.Q.; supervision, project administration, funding acquisition, L.H.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32170484), the Fundamental Research Funds for the Central Universities (2572019AA22) and the China Postdoctoral Science Foundation (2023M730532).
Data Availability Statement
All data used in this paper are available within the text.
Acknowledgments
We thank Norbert Perrimon and Lei Xue for gifting the fly stocks and István Andó for providing the antibodies. We also thank the Bloomington Drosophila Stock Center, Vienna Drosophila Resource Center and TsingHua Fly Center for providing the fly stocks.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Yu, S.; Luo, F.; Xu, Y.; Zhang, Y.; Jin, L.H. Drosophila innate immunity involves multiple signaling pathways and coordinated communication between different tissues. Front. Immunol. 2022, 13, 905370. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.J.; Hartenstein, V.; Banerjee, U. Thicker than blood: Conserved mechanisms in Drosophila and vertebrate hematopoiesis. Dev. Cell. 2003, 5, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Lanot, R.; Zachary, D.; Holder, F.; Meister, M. Postembryonic hematopoiesis in Drosophila. Dev. Biol. 2001, 230, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, U.; Girard, J.R.; Goins, L.M.; Spratford, C.M. Drosophila as a genetic model for hematopoiesis. Genetics 2019, 211, 367–417. [Google Scholar] [CrossRef]
- Melcarne, C.; Lemaitre, B.; Kurant, E. Phagocytosis in Drosophila: From molecules and cellular machinery to physiology. Insect Biochem. Mol. Biol. 2019, 109, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Krishnamoorthy, P.; Walker, E.C.; Joshi, H.; Morley, S.C. Expression of non-phosphorylatable S5A-L-plastin exerts phenotypes distinct from L-plastin deficiency during podosome formation and phagocytosis. Front. Cell Dev. Biol. 2023, 11, 1020091. [Google Scholar] [CrossRef]
- Ulvila, J.; Vanha-Aho, L.M.; Rämet, M. Drosophila phagocytosis—Still many unknowns under the surface. Apmis 2011, 119, 651–662. [Google Scholar] [CrossRef]
- Rizki, T.M.; Rizki, R.M. Lamellocyte differentiation in Drosophila larvae parasitized by Leptopilina. Dev. Comp. Immunol. 1992, 16, 103–110. [Google Scholar] [CrossRef]
- Yu, S.; Luo, F.; Jin, L.H. The Drosophila lymph gland is an ideal model for studying hematopoiesis. Dev. Comp. Immunol. 2018, 83, 60–69. [Google Scholar] [CrossRef]
- Levine, B. Eating oneself and uninvited guests: Autophagy-related pathways in cellular defense. Cell 2005, 120, 159–162. [Google Scholar] [PubMed]
- Das, G.; Shravage, B.V.; Baehrecke, E.H. Regulation and function of autophagy during cell survival and cell death. Cold Spring Harb. Perspect. Biol. 2012, 4, a008813. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, L.; Zhang, G.; Shangguan, G.; Hou, X.; Duan, W.; Xi, Y.; Xu, N.; Zhang, B.; Dong, J.; et al. A nonautophagic role of ATG5 in regulating cell growth by targeting c-Myc for proteasome-mediated degradation. iScience 2021, 24, 103296. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, J.; Zhao, J.; Ma, N.; Kim, S.W.; Qiao, S.; Ma, X. Autophagy: The last defense against cellular nutritional stress. Adv. Nutr. 2018, 9, 493–504. [Google Scholar] [PubMed]
- King, J.S.; Veltman, D.M.; Insall, R.H. The induction of autophagy by mechanical stress. Autophagy 2011, 7, 1490–1499. [Google Scholar] [CrossRef]
- Chun, Y.; Kim, J. Autophagy: An essential degradation program for cellular homeostasis and life. Cells 2018, 7, 278. [Google Scholar]
- Mariño, G.; Madeo, F.; Kroemer, G. Autophagy for tissue homeostasis and neuroprotection. Curr. Opin. Cell Biol. 2011, 23, 198–206. [Google Scholar] [CrossRef]
- Allen, E.A.; Baehrecke, E.H. Autophagy in animal development. Cell Death Differ. 2020, 27, 903–918. [Google Scholar] [CrossRef]
- Jiang, P.; Mizushima, N. Autophagy and human diseases. Cell Res. 2014, 24, 69–79. [Google Scholar] [CrossRef]
- Chang, Y.Y.; Neufeld, T.P. Autophagy takes flight in Drosophila. FEBS Lett. 2010, 584, 1342–1349. [Google Scholar] [CrossRef]
- McPhee, C.K.; Baehrecke, E.H. Autophagy in Drosophila melanogaster. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.C.; Juhász, G.; Neufeld, T.P. Direct induction of autophagy by Atg1 inhibits cell growth and induces apoptotic cell death. Curr. Biol. 2007, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ayachit, M.S.; Shravage, B.V. Atg1 modulates mitochondrial dynamics to promote germline stem cell maintenance in Drosophila. Biochem. Biophys. Res. Commun. 2023, 643, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Sandford, E.; Gatica, D.; Qiu, Y.; Liu, X.; Zheng, Y.; Schulman, B.A.; Xu, J.; Semple, I.; Ro, S.H.; et al. Mutation in ATG5 reduces autophagy and leads to ataxia with developmental delay. Elife 2016, 5, e12245. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Nicolson, S.; Denton, D.; Kumar, S. Distinct requirements of Autophagy-related genes in programmed cell death. Cell Death. Differ. 2015, 22, 1792–1802. [Google Scholar] [CrossRef]
- Simonsen, A.; Cumming, R.C.; Lindmo, K.; Galaviz, V.; Cheng, S.; Rusten, T.E.; Finley, K.D. Genetic modifiers of the Drosophila blue cheese gene link defects in lysosomal transport with decreased life span and altered ubiquitinated-protein profiles. Genetics 2007, 176, 1283–1297. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kuo, C.J.; Hansen, M.; Troemel, E. Autophagy and innate immunity: Insights from invertebrate model organisms. Autophagy 2018, 14, 233–242. [Google Scholar] [CrossRef]
- Moy, R.H.; Cherry, S. Antimicrobial autophagy: A conserved innate immune response in Drosophila. J. Innate Immun. 2013, 5, 444–455. [Google Scholar] [CrossRef]
- Shelly, S.; Lukinova, N.; Bambina, S.; Berman, A.; Cherry, S. Autophagy is an essential component of Drosophila immunity against vesicular stomatitis virus. Immunity 2009, 30, 588–598. [Google Scholar] [CrossRef]
- Deretic, V.; Levine, B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe. 2009, 5, 527–549. [Google Scholar] [CrossRef] [PubMed]
- Laczkó-Dobos, H.; Maddali, A.K.; Jipa, A.; Bhattacharjee, A.; Végh, A.G.; Juhász, G. Lipid profiles of autophagic structures isolated from wild type and Atg2 mutant Drosophila. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158868. [Google Scholar] [CrossRef]
- Harrison, D.A.; Binari, R.; Nahreini, T.S.; Gilman, M.; Perrimon, N. Activation of a Drosophila Janus kinase (JAK) causes hematopoietic neoplasia and developmental defects. EMBO J. 1995, 14, 2857–2865. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Pan, P.C.; Govind, S. A role for the Drosophila Toll/Cactus pathway in larval hematopoiesis. Development 1998, 125, 1909–1920. [Google Scholar] [CrossRef]
- Zettervall, C.J.; Anderl, I.; Williams, M.J.; Palmer, R.; Kurucz, E.; Ando, I.; Hultmark, D. A directed screen for genes involved in Drosophila blood cell activation. Proc. Natl. Acad. Sci. USA 2004, 101, 14192–14197. [Google Scholar] [CrossRef] [PubMed]
- Minakhina, S.; Steward, R. Melanotic mutants in Drosophila: Pathways and phenotypes. Genetics 2006, 174, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Lindner, B.; Burkard, T.; Schuler, M. Phagocytosis assays with different pH-sensitive fluorescent particles and various readouts. Biotechniques 2020, 68, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Péan, C.B.; Schiebler, M.; Tan, S.W.; Sharrock, J.A.; Kierdorf, K.; Brown, K.P.; Maserumule, M.C.; Menezes, S.; Pilátová, M.; Bronda, K.; et al. Regulation of phagocyte triglyceride by a STAT-ATG2 pathway controls mycobacterial infection. Nat. Commun. 2017, 8, 14642. [Google Scholar] [CrossRef]
- Horsthemke, M.; Bachg, A.C.; Groll, K.; Moyzio, S.; Müther, B.; Hemkemeyer, S.A.; Wedlich-Söldner, R.; Sixt, M.; Tacke, S.; Bähler, M.; et al. Multiple roles of filopodial dynamics in particle capture and phagocytosis and phenotypes of Cdc42 and Myo10 deletion. J. Biol. Chem. 2017, 292, 7258–7273. [Google Scholar] [CrossRef]
- Kress, H.; Stelzer, E.H.; Holzer, D.; Buss, F.; Griffiths, G.; Rohrbach, A. Filopodia act as phagocytic tentacles and pull with discrete steps and a load-dependent velocity. Proc. Natl. Acad. Sci. USA 2007, 104, 11633–11638. [Google Scholar] [CrossRef]
- Kadandale, P.; Stender, J.D.; Glass, C.K.; Kiger, A.A. Conserved role for autophagy in Rho1-mediated cortical remodeling and blood cell recruitment. Proc. Natl. Acad. Sci. USA 2010, 107, 10502–10507. [Google Scholar] [CrossRef]
- Hao, Y.; Yu, S.; Luo, F.; Jin, L.H. Jumu is required for circulating hemocyte differentiation and phagocytosis in Drosophila. Cell Commun. Signal 2018, 16, 95. [Google Scholar]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster--from microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Kim, L.K.; Choi, U.Y.; Cho, H.S.; Lee, J.S.; Lee, W.B.; Kim, J.; Jeong, K.; Shim, J.; Kim-Ha, J.; Kim, Y.J. Down-regulation of NF-kappaB target genes by the AP-1 and STAT complex during the innate immune response in Drosophila. PLoS Biol. 2007, 5, e238. [Google Scholar] [CrossRef]
- Irving, P.; Ubeda, J.M.; Doucet, D.; Troxler, L.; Lagueux, M.; Zachary, D.; Hoffmann, J.A.; Hetru, C.; Meister, M. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol. 2005, 7, 335–350. [Google Scholar] [CrossRef]
- Charroux, B.; Royet, J. Elimination of plasmatocytes by targeted apoptosis reveals their role in multiple aspects of the Drosophila immune response. Proc. Natl. Acad. Sci. USA 2009, 106, 9797–9802. [Google Scholar] [CrossRef]
- Defaye, A.; Evans, I.; Crozatier, M.; Wood, W.; Lemaitre, B.; Leulier, F. Genetic ablation of Drosophila phagocytes reveals their contribution to both development and resistance to bacterial infection. J. Innate Immun. 2009, 1, 322–334. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).