
Paving Behavior in Ants and Its Potential Application in Monitoring Two Urban Pest Ants, Solenopsis invicta and Tapinoma melanocephalum



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Efficiency of Adhesive Tapes to Detect Solenopsis invicta and Tapinoma melanocephalum
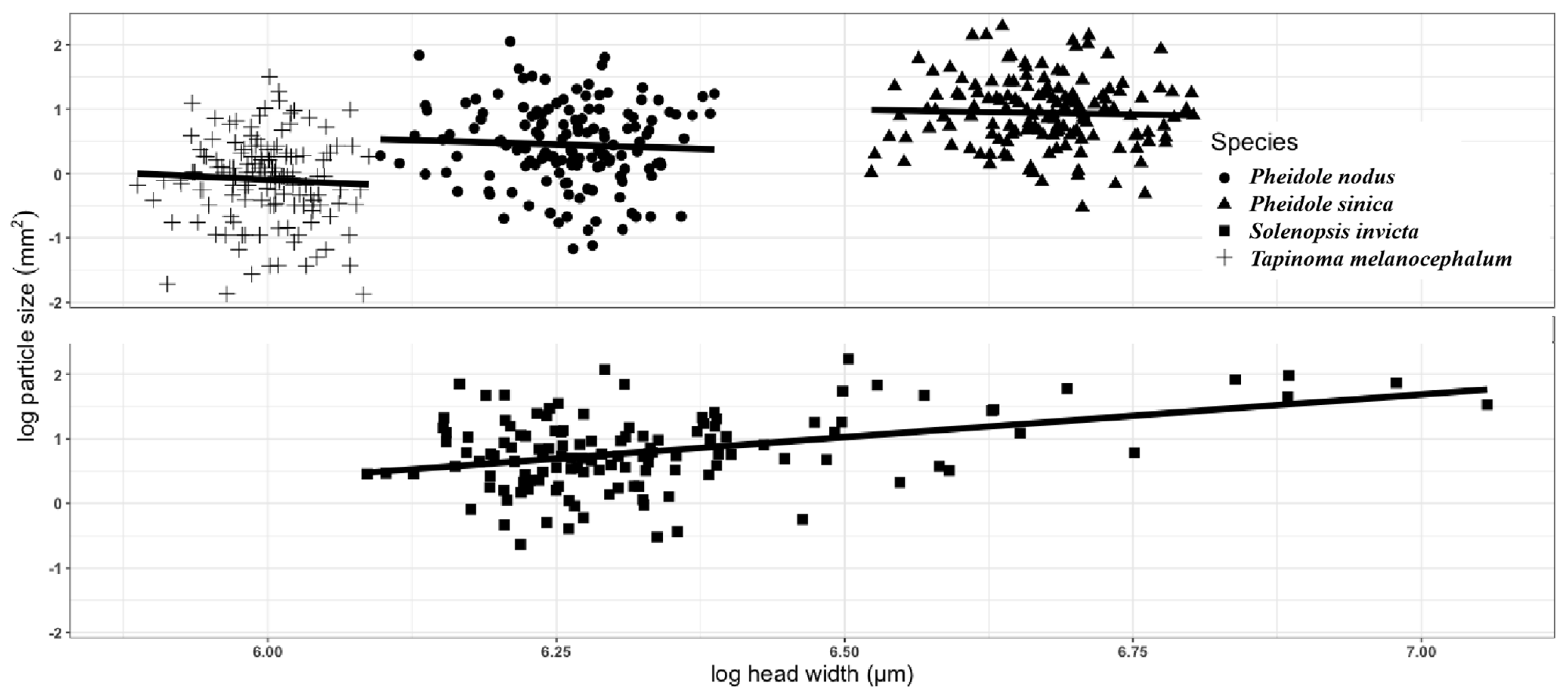
2.2. Correlation between Head Width and Particle Area of Active Tape-Paving Ant Species
2.3. Data Analyses
3. Results
3.1. Efficiency of Adhesive Tapes to Detect Solenopsis invicta and Tapinoma melanocephalum
3.2. Distinguishment of Solenopsis invicta, Tapinoma melanocephalum, and Non-Target Ants Performing the Tape-Paving Behavior
3.3. Correlation between Head Width and Particle Area of Active Tape-Paving Ant Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, S.F.; DeShazo, R.D.; Moffitt, J.E.; Williams, D.F.; Buhner, W.A., II. Expanding habitat of the imported fire ant (Solenopsis invicta): A public health concern. J. Allergy Clin. Immunol. 2000, 105, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zeng, L.; Xu, Y.; Lu, Y. Prevalence and management of Solenopsis invicta in China. NeoBiota 2020, 54, 89–124. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, J.; Zhou, A.; Zeng, L. Prevalence of Solenopsis invicta (Hymenoptera: Formicidae) venom allergic reactions in mainland China. Fla. Entomol. 2012, 95, 961–965. [Google Scholar] [CrossRef]
- Wang, L.; Lu, Y.; Li, R.; Zeng, L.; Du, J.; Huang, X.; Xu, Y. Mental health effects caused by red imported fire ant attacks (Solenopsis invicta). PLoS ONE 2018, 13, e0199424. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Lin, S.; Chen, X.; Chen, J.; Wang, L.; Xiong, H.; Xie, Q.; Sun, Z.; Wen, X.; Wang, C. Food transport of red imported fire ants (Hymenoptera: Formicidae) on vertical surfaces. Sci. Rep. 2019, 9, 3283. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Xiong, H.; Wen, Y.; Wen, X.; Wang, H.; Fang, Y.; Ma, T.; Sun, Z.; Chen, X.; Wang, C. Laboratory and field evaluation of the repellency of six preservatives to red imported fire ants (Hymenoptera: Formicidae). J. Asia-Pac. Entomol. 2017, 20, 535–540. [Google Scholar] [CrossRef]
- Valles, S.M.; Wylie, R.; Burwell, C.J.; McNaught, M.K.; Horlock, C. Evaluation of a lateral flow immunoassay for field identification of Solenopsis invicta (Hymenoptera: Formicidae) in Australia. Austral. Entomol. 2019, 58, 324–328. [Google Scholar] [CrossRef]
- Nakajima, N.; Sakamoto, Y.; Goka, K. Rapid detection of the red fire ant Solenopsis invicta (Hymenoptera: Formicidae) by loop-mediated isothermal amplification. Appl. Entomol. Zool. 2019, 54, 319–322. [Google Scholar] [CrossRef]
- Wen, C.; Chen, J.; Qin, W.Q.; Chen, X.; Cai, J.C.; Wen, J.B.; Wen, X.J.; Wang, C. Red imported fire ants (Hymenoptera: Formicidae) cover inaccessible surfaces with particles to facilitate food search and transportation. Insect Sci. 2021, 28, 1816–1828. [Google Scholar] [CrossRef]
- Choe, H.C.; Lim, J.O.; Lee, S. Tapinoma melanocephalum (Fabricius), a new exotic pest in Korea. J. Asia-Pac. Entomol. 2009, 12, 67–69. [Google Scholar] [CrossRef]
- MacGown, J.A.; Hill, J.G. Tapinoma melanocephalum (Hymenoptera: Formicidae), a new exotic ant in Mississippi. J. Miss. Acad. Sci. 2009, 54, 172–174. [Google Scholar]
- Wetterer, J.K. Worldwide spread of the ghost ant, Tapinoma melanocephalum (Hymenoptera: Formicidae). Myrmecol. News 2009, 12, 2333. [Google Scholar]
- Zima, J.; Lebrasseur, O.; Borovanska, M.; Janda, M. Identification of microsatellite markers for a worldwide distributed, highly invasive ant species Tapinoma melanocephalum (Hymenoptera: Formicidae). Eur. J. Entomol. 2016, 113, 409–414. [Google Scholar] [CrossRef]
- Zhou, A.M.; Kuang, B.; Gao, Y.; Liang, G.W. Density-dependent benefits in ant-hemipteran mutualism? The case of the ghost ant Tapinoma melanocephalum (Hymenoptera: Formicidae) and the invasive mealybug Phenacoccus solenopsis (Hemiptera: Pseudococcidae). PLoS ONE 2015, 10, e0123885. [Google Scholar] [CrossRef]
- Zhou, A.M.; Liang, G.W.; Zeng, L.; Lu, Y.Y.; Xu, Y.J. Interactions between ghost ants and invasive mealybugs: The case of Tapinoma melanocephalum (Hymenoptera: Formicidae) and Phenacoccus solenopsis (Hemiptera: Pseudococcidae). Fla. Entomol. 2014, 97, 1474–1480. [Google Scholar] [CrossRef]
- Feng, D.D.; Michaud, J.P.; Li, P.; Zhou, Z.S.; Xu, Z.F. The native ant, Tapinoma melanocephalum, improves the survival of an invasive mealybug, Phenacoccus solenopsis, by defending it from parasitoids. Sci. Rep. 2015, 5, 15691. [Google Scholar] [CrossRef]
- Zheng, C.; Yang, F.; Zeng, L.; Vargo, E.L.; Xu, Y. Genetic diversity and colony structure of Tapinoma melanocephalum on the islands and mainland of South China. Ecol. Evol. 2018, 8, 5427–5440. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, Y.; Zeng, L.; Liang, G. Foraging behavior and recruitment of red imported fire ant Solenopsis invicta Buren in typical habitats of South China. Acta Ecol. Sin. 2007, 27, 855–860. [Google Scholar]
- Wilder, S.M.; Holway, D.A.; Suarez, A.V.; LeBrun, E.G.; Eubanks, M.D. Intercontinental differences in resource use reveal the importance of mutualisms in fire ant invasions. Proc. Natl. Acad. Sci. USA 2011, 108, 20639–20644. [Google Scholar] [CrossRef]
- Pembury Smith, M.Q.R.; Ruxton, G.D. Effective use of the McNemar test. Behav. Ecol. Sociobiol. 2020, 74, 133. [Google Scholar] [CrossRef]
- Brassard, F.; Leong, C.M.; Chan, H.H.; Guénard, B. High diversity in urban areas: How comprehensive sampling reveals high ant species richness within one of the most urbanized regions of the world. Diversity 2021, 13, 358. [Google Scholar] [CrossRef]
- Chan, K.H.; Guénard, B. Ecological and socio-economic impacts of the red import fire ant, Solenopsis invicta (Hymenoptera: Formicidae), on urban agricultural ecosystems. Urban Ecosyst. 2020, 23, 112. [Google Scholar] [CrossRef]
- Beck, B.B. Animal Tool Behavior; Garland STPM Press: New York, NY, USA, 1980. [Google Scholar]
- Wang, C.; Chen, X.; Hooper-Bùi, L.M.; Strecker, R.; Wen, Y.Z.; Qin, W.Q.; Ma, T.; Zhao, H.S.; Chen, X.Y.; Wen, X.J. How do groups of red imported fire ants (Hymenoptera: Formicidae) feed on a droplet of sugar water? Insect Sci. 2018, 25, 499–507. [Google Scholar] [CrossRef]
- Zhou, A.M.; Du, Y.; Chen, J. Ants adjust their tool use strategy in response to foraging risk. Funct. Ecol. 2020, 34, 2524–2535. [Google Scholar] [CrossRef]
- Zhou, A.M.; Du, Y.; Chen, J. Surfactant affects the tool use behavior of foraging ants. Ecoto. Environ. Safe 2022, 238, 113592. [Google Scholar] [CrossRef]
- Maák, I.; Lőrinczi, G.; Le Quinquis, P.; Módra, G.; Bovet, D.; Call, J.; d’Ettorre, P. Tool selection during foraging in two species of funnel ants. Anim. Behav. 2017, 123, 207–216. [Google Scholar] [CrossRef]
- Lőrinczi, G.; Módra, G.; Juhász, O.; Maák, I. Which tools to use? Choice optimization in the tool-using ant, Aphaenogaster subterranea. Behav. Ecol. 2018, 29, 1444–1452. [Google Scholar] [CrossRef]
- Módra, G.; Maák, I.; Lőrincz, Á.; Lőrinczi, G. Comparison of foraging tool use in two species of myrmicine ants (Hymenoptera: Formicidae). Insect Soc. 2022, 69, 512. [Google Scholar] [CrossRef]
- Pirk, G.I.; Lopez de Casenave, J. Influence of seed size on feeding preferences and diet composition of three sympatric harvester ants in the central Monte Desert, Argentina. Ecol. Res. 2010, 25, 439–445. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Nais, J.; Linsenmair, K. Worker size and seed size selection in ‘seed’-collecting ant ensembles (Hymenoptera: Formicidae) in primary rain forests on Borneo. J. Trop. Ecol. 2006, 22, 685–693. [Google Scholar] [CrossRef]
- Arnan, X.; Retana, J.; Rodrigo, A.; Cerdá, X. Foraging behaviour of harvesting ants determines seed removal and dispersal. Insect Soc. 2010, 57, 421–430. [Google Scholar] [CrossRef]
- Ipser, R.M.; Gardner, W.A. Particle size preference of six ant species (Hymenoptera: Formicidae). J. Entomol. Sci. 2019, 54, 370–377. [Google Scholar] [CrossRef]
- Kaspari, M. Worker size and seed size selection by harvester ants in a neotropical forest. Oecologia 1996, 105, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Hooper-Bùi, L.M.; Appel, A.G.; Rust, M.K. Preference of food particle size among several urban ant species. J. Econ. Entomol. 2002, 95, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Neff, R.; Puckett, R.T.; Gold, R.E. Particle size and bait preference of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Sociobiology 2011, 58, 473–488. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | No. of Sites | Number of Sites Detecting Solenopsis invicta | |||
|---|---|---|---|---|---|
| Baiting | Taping | Pitfall Trapping | |||
| Determined by Ant Infestation on the Sausage | Determined by Tape Paving | ||||
| 1 | 224 | 211 a | 215 a | 214 a | - |
| 2 | 224 | 0 a | 0 a | 0 a | - |
| 3 | 199 | 74 b | 107 a | 106 a | 86 b |
| 4 | 200 | 181 a | 184 a | 184 a | - |
| 5 | 199 | 190 b | 197 a | 193 ab | - |
| 6 | 197 | 73 b | 86 a | 86 a | - |
| 7 | 200 | 145 ab | 154 a | 138 b | - |
| 8 | 200 | 134 a | 143 a | 141 a | - |
| 9 | 199 | 134 a | 131 a | 129 a | - |
| 10 | 200 | 50 a | 46 a | 44 a | - |
| 11 | 199 | 109 a | 100 b | 100 b | - |
| 12 | 200 | 60 a | 58 a | 54 a | - |
| 13 | 200 | 69 a | 67 a | 65 a | 70 a |
| 14 | 200 | 75 a | 74 a | 72 a | - |
| 15 | 195 | 62 a | 60 a | 52 b | - |
| 16 | 198 | 66 b | 78 a | 77 a | - |
| 17 | 196 | 0 a | 0 a | 0 a | - |
| 18 | 196 | 2 a | 2 a | 2 a | - |
| 19 | 191 | 93 a | 99 a | 99 a | - |
| 20 | 181 | 95 a | 99 a | 97 a | - |
| No. | No. of Sites | Number of Sites Detecting Tapinoma melanocephalum | |||
|---|---|---|---|---|---|
| Baiting | Taping | Pitfall Trapping | |||
| Determined by Ant Infestation on the Sausage | Determined by Tape Paving | ||||
| 1 | 224 | 6 a | 2 a | 2 a | - |
| 2 | 224 | 92 a | 82 a | 82 a | - |
| 3 | 199 | 98 a | 85 b | 85 b | 97 a |
| 4 | 200 | 0 a | 0 a | 0 a | - |
| 5 | 199 | 0 a | 0 a | 0 a | - |
| 6 | 197 | 19 a | 18 a | 18 a | - |
| 7 | 200 | 2 a | 2 a | 2 a | - |
| 8 | 200 | 14 a | 16 a | 16 a | - |
| 9 | 199 | 19 a | 17 a | 17 a | - |
| 10 | 200 | 0 a | 0 a | 0 a | - |
| 11 | 199 | 33 a | 38 a | 37 a | - |
| 12 | 200 | 2 a | 1 a | 1 a | - |
| 13 | 200 | 8 a | 6 a | 6 a | 5 a |
| 14 | 200 | 17 a | 15 a | 15 a | - |
| 15 | 195 | 7 a | 7 a | 7 a | - |
| 16 | 198 | 17 a | 6 b | 5 b | - |
| 17 | 196 | 0 a | 0 a | 0 a | - |
| 18 | 196 | 9 a | 9 a | 9 a | - |
| 19 | 191 | 2 a | 2 a | 2 a | - |
| 20 | 181 | 5 a | 5 a | 4 a | - |
| No. | No. of Sites | Number of Sites Detecting Non-Target Ants | |||
|---|---|---|---|---|---|
| Baiting | Taping | Pitfall Trapping | |||
| Determined by Ant Infestation on the Sausage | Determined by Tape Paving | ||||
| 1 | 224 | 4 ab | 6 a | 0 b | - |
| 2 | 224 | 124 a | 120 a | 5 b | - |
| 3 | 199 | 9 b | 5 bc | 1 c | 24 a |
| 4 | 200 | 16 a | 14 a | 1 b | - |
| 5 | 199 | 2 a | 2 a | 0 a | - |
| 6 | 197 | 52 b | 64 a | 4 c | - |
| 7 | 200 | 2 a | 1 a | 0 a | - |
| 8 | 200 | 4 ab | 8 a | 0 b | - |
| 9 | 199 | 7 a | 11 a | 0 b | - |
| 10 | 200 | 114 a | 124 a | 2 b | - |
| 11 | 199 | 24 a | 31 a | 0 b | - |
| 12 | 200 | 43 a | 41 a | 0 b | - |
| 13 | 200 | 95 a | 104 a | 5 b | 99 a |
| 14 | 200 | 45 b | 53 a | 0 c | - |
| 15 | 195 | 78 a | 80 a | 0 b | - |
| 16 | 198 | 47 b | 70 a | 3 c | - |
| 17 | 196 | 182 a | 175 a | 148 b | - |
| 18 | 196 | 120 b | 130 a | 74 c | - |
| 19 | 191 | 19 a | 17 a | 0 b | - |
| 20 | 181 | 51 a | 45 a | 5 b | - |
| Subfamily | Species | Baiting | Taping (Determined by Ant Infestation on the Sausage) | Taping (Determined by Tape Paving) | Pitfall Trapping | ||||
|---|---|---|---|---|---|---|---|---|---|
| No. of Baits Detecting the Ant Species | Percentage of Baits Detecting the Ant Species (%) | No. of Tapes Detecting the Ant Species | Percentage of Tapes Detecting the Ant Species (%) | No. of Tapes Detecting the Ant Species | Percentage of Tapes Detecting the Ant Species (%) | No. of Pitfall Traps Detecting the Ant Species | Percentage of Pitfall Traps Detecting the Ant Species (%) | ||
| Myrmicinae | Solenopsis invicta * | 1823 | 45.60 | 1900 | 47.52 | 1853 | 46.35 | 156 | 39.10 |
| Dolichoderinae | Tapinoma melanocephalum * | 350 | 8.75 | 311 | 7.78 | 308 | 7.70 | 102 | 25.56 |
| Myrmicinae | Pheidole sinica * | 135 | 3.38 | 126 | 3.15 | 118 | 2.95 | 0 | 0 |
| Ponerinae | Odontoponera transversa * | 134 | 3.35 | 145 | 3.63 | 37 | 0.93 | 28 | 7.02 |
| Formicinae | Paratrechina longicornis | 133 | 3.33 | 104 | 2.60 | 0 | 0 | 7 | 1.75 |
| Myrmicinae | Pheidole parva * | 130 | 3.25 | 138 | 3.45 | 3 | 0.08 | 12 | 3.01 |
| Myrmicinae | Monomorium chinense | 104 | 2.60 | 108 | 2.70 | 0 | 0 | 21 | 5.26 |
| Myrmicinae | Pheidole nodus * | 87 | 2.18 | 95 | 2.38 | 58 | 1.45 | 7 | 1.75 |
| Formicinae | Camponotus nicobarensis * | 80 | 2.00 | 98 | 2.45 | 7 | 0.18 | 16 | 4.01 |
| Myrmicinae | Tetramorium caespitum | 48 | 1.20 | 58 | 1.45 | 0 | 0 | 7 | 1.75 |
| Myrmicinae | Crematogaster rogenhoferi | 57 | 1.43 | 64 | 1.60 | 0 | 0 | 4 | 1.00 |
| Ponerinae | Diacamma rugosum | 31 | 0.78 | 36 | 0.90 | 0 | 0 | 9 | 2.26 |
| Myrmicinae | Monomorium pharaonis | 25 | 0.63 | 21 | 0.53 | 0 | 0 | 0 | 0 |
| Myrmicinae | Pheidole pieli | 20 | 0.50 | 18 | 0.45 | 0 | 0 | 0 | 0 |
| Formicinae | Anoplolepis gracilipes | 18 | 0.50 | 34 | 0.94 | 0 | 0 | 0 | 0 |
| Formicinae | Polyrhachis dives | 15 | 0.41 | 18 | 0.50 | 0 | 0 | 0 | 0 |
| Myrmicinae | Carebara affinis * | 14 | 0.39 | 14 | 0.39 | 14 | 0.39 | 0 | 0 |
| Myrmicinae | Pheidole yeensis * | 10 | 0.28 | 9 | 0.25 | 4 | 0.11 | 12 | 3.01 |
| Tape-Paving Ant Species | Head Width of Workers (mm) | Head Width of Soldiers (mm) | χ2 | df | p-Value |
|---|---|---|---|---|---|
| Solenopsis invicta | 0.583 ± 0.150 d | - | - | - | - |
| Tapinoma melanocephalum | 0.402 ± 0.019 g | - | - | - | - |
| Pheidole parva | 0.450 ± 0.020 f | 0.783 ± 0.050 d | 106.78 | 1 | <0.0001 * |
| Pheidole yeensis | 0.597 ± 0.021 d | 1.720 ± 0.057 c | 91.14 | 1 | <0.0001 * |
| Pheidole nodus | 0.536 ± 0.028 e | 1.717 ± 0.041 c | 113.98 | 1 | <0.0001 * |
| Pheidole sinica | 0.799± 0.038 c | 2.971 ± 0.038 a | 57.51 | 1 | <0.0001 * |
| Carebara affinis | 0.536 ± 0.027 e | 1.888 ± 0.185 b | 105.28 | 1 | <0.0001 * |
| Odontoponera transversa | 1.966 ± 0.049 a | - | - | - | - |
| Camponotus nicobarensis | 1.896 ± 0.035 b | - | - | - | - |
| χ2 | 830.44 | 159.93 | |||
| df | 8 | 4 | |||
| p-value | <0.0001 | <0.0001 |
| Species | Estimate | SD | t-Value | p-Value | |
|---|---|---|---|---|---|
| Solenopsis invicta | Intercept | −7.583 | 1.746 | −4.344 | <0.001 |
| Head width | 1.325 | 0.276 | 4.807 | <0.001 | |
| Pheidole sinica | Intercept | 2.861 | 4.956 | 0.577 | 0.565 |
| Head width | −0.287 | 0.743 | −0.387 | 0.699 | |
| Pheidole nodus | Intercept | 3.786 | 5.735 | 0.660 | 0.510 |
| Head width | −0.534 | 0.916 | −0.583 | 0.561 | |
| Tapinoma melanocephalum | Intercept | 5.195 | 7.983 | 0.651 | 0.516 |
| Head width | −0.882 | 1.331 | −0.663 | 0.509 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Wen, C.; Chen, X.; Hua, Y.; Du, C.; Cai, J.; Wen, X.; Wang, L.; Wang, C. Paving Behavior in Ants and Its Potential Application in Monitoring Two Urban Pest Ants, Solenopsis invicta and Tapinoma melanocephalum. Insects 2023, 14, 219. https://doi.org/10.3390/insects14030219
Shen L, Wen C, Chen X, Hua Y, Du C, Cai J, Wen X, Wang L, Wang C. Paving Behavior in Ants and Its Potential Application in Monitoring Two Urban Pest Ants, Solenopsis invicta and Tapinoma melanocephalum. Insects. 2023; 14(3):219. https://doi.org/10.3390/insects14030219
Chicago/Turabian StyleShen, Liming, Chao Wen, Xuan Chen, Yan Hua, Chengju Du, Jiacheng Cai, Xiujun Wen, Lei Wang, and Cai Wang. 2023. "Paving Behavior in Ants and Its Potential Application in Monitoring Two Urban Pest Ants, Solenopsis invicta and Tapinoma melanocephalum" Insects 14, no. 3: 219. https://doi.org/10.3390/insects14030219
APA StyleShen, L., Wen, C., Chen, X., Hua, Y., Du, C., Cai, J., Wen, X., Wang, L., & Wang, C. (2023). Paving Behavior in Ants and Its Potential Application in Monitoring Two Urban Pest Ants, Solenopsis invicta and Tapinoma melanocephalum. Insects, 14(3), 219. https://doi.org/10.3390/insects14030219