


Influence of Soldiers on Exploratory Foraging Behavior in the Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Termites
2.2. Foraging Arena Setup
2.3. Foraging Behavior Assay
2.4. Data Analysis
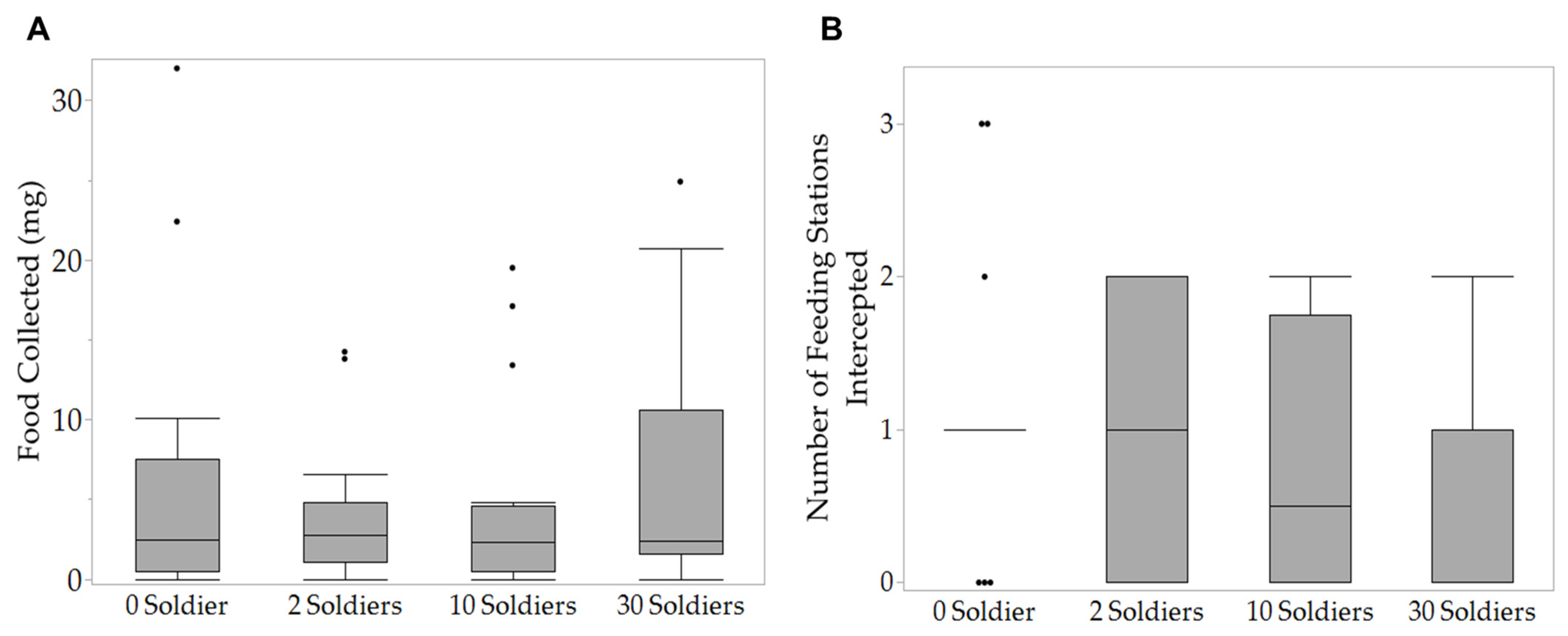
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bignell, D.E.; Roisin, Y.; Lo, N. Biology of Termites: A Modern Synthesis, 2nd ed.; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar]
- Tian, L.; Zhou, X. The soldiers in societies: Defense, regulation, and evolution. Int. J. Biol. Sci. 2014, 10, 296–308. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Grace, J.K. Effect of termite soldiers on the foraging behavior of Coptotermes formosanus (Isoptera: Rhinotermitidae) in the presence of predatory ants. Sociobiology 1997, 29, 247–253. [Google Scholar]
- Matsuura, K. Colony-level stabilization of soldier head width for head-plug defense in the termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Behav. Ecol. Sociobiol. 2002, 51, 172–179. [Google Scholar] [CrossRef]
- Delinge, J.; Quennedey, A.; Blum, M.S. The Enemies and Defense Mechanisms of Termites. In Social Insects; Academic Press: Athens, GA, USA, 1981; Volume 2. [Google Scholar]
- Henderson, G. Primer Pheromones and Possible Soldier Caste Influence on the Evolution of Sociality in Lower Termites. In Pheromone Communication in Social Insects; Westview Press: Boulder, CO, USA, 1998; pp. 314–330. [Google Scholar]
- Haverty, M. The proportion of soldiers in termite colonies: A list and a bibliography (Isoptera). Sociobiology 1977, 2, 199–216. [Google Scholar]
- Su, N.-Y.; LaFage, J.P. Effects of starvation on survival and maintenance of soldier proportion in laboratory groups of the Formosan subterranean termite, Coptotermes formosanus (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 1986, 79, 312–316. [Google Scholar] [CrossRef]
- Waller, D.A.; La Fage, J.P. Environmental influence on soldier differentiation in Coptotermes formosanus Shiraki (Rhinotermitidae). Insectes Soc. 1988, 35, 144–152. [Google Scholar] [CrossRef]
- Bardunias, P.M.; Su, N.-Y. Queue size determines the width of tunnels in the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Insect Behav. 2010, 23, 189–204. [Google Scholar] [CrossRef]
- Cao, R.; Su, N.-Y. Temperature preferences of four subterranean termite species (Isoptera: Rhinotermitidae) and temperature-dependent survivorship and wood-consumption rate. Ann. Entomol. Soc. Am. 2016, 109, 64–71. [Google Scholar] [CrossRef]
- Hedlund, J.C.; Henderson, G. Effect of available food size on search tunnel formation by the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1999, 92, 610–616. [Google Scholar] [CrossRef]
- Haverty, M.I.; LaFage, J.P.; Nutting, W.L. Seasonal activity and environmental control of foraging of the subterranean termite, Heterotermes aureus (Snyder), in a desert grassland. Life Sci. 1974, 15, 1091–1101. [Google Scholar] [CrossRef]
- Alamu, O.T.; Ewete, F.K. Influence of seasonal changes, weather factors and soil depth on the foraging activities of subterranean termites in Eucalyptus plantations. Int. J. Trop. Insect Sci. 2021, 41, 1213–1221. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Osbrink, W.L.A. Effect of seasonal changes in soil temperature and moisture on wood consumption and foraging activity of Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2011, 104, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, N.; Gile, G.H.; Pratt, S.C. Behavioral rules for soil excavation by colony founders and workers in termites. Ann. Entomol. Soc. Am. 2020, 114, 654–661. [Google Scholar] [CrossRef]
- Lee, S.-H.; Bardunias, P.M.; Su, N.-Y. Optimal length distribution of termite tunnel branches for efficient food search and resource transportation. BioSystems 2007, 90, 802–807. [Google Scholar] [CrossRef]
- Bordereau, C.; Pasteels, J.M. Pheromones and Chemical Ecology of Dispersal and Foraging in Termites. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 279–320. [Google Scholar]
- Traniello, J.F.A.; Leuthold, R.H. Behavior and Ecology of Foraging in Termites. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 141–168. [Google Scholar]
- Cao, R.; Su, N.-Y. Tunneling and food transportation activity of four subterranean termite species (Isoptera: Rhinotermitidae) at various temperatures. Ann. Entomol. Soc. Am. 2014, 107, 696–701. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Tuma, J.; Eggleton, P.; Fayle, T.M. Ant-termite interactions: An important but under-explored ecological linkage. Biol. Rev. 2020, 95, 555–572. [Google Scholar] [CrossRef]
- Modlmeier, A.P.; Keiser, C.N.; Watters, J.V.; Sih, A.; Pruitt, J.N. The keystone individual concept: An ecological and evolutionary overview. Anim. Behav. 2014, 89, 53–62. [Google Scholar] [CrossRef]
- Tian, L.; Preisser, E.L.; Haynes, K.F.; Zhou, X. Social buffering in a eusocial invertebrate: Termite soldiers reduce the lethal impact of competitor cues on workers. Ecology 2017, 98, 952–960. [Google Scholar] [CrossRef]
- Janowiecki, M.A.; Vargo, E.L. Effect of soldiers on collective tunneling behavior in three species of Reticulitermes (Blattodea: Rhinotermitidae). Insect Soc. 2022, 69, 237–245. [Google Scholar] [CrossRef]
- Traniello, J.F.A. Enemy deterrence in the recruitment strategy of a termite: Soldier-organized foraging in Nasutitermes costalis. Proc. Natl. Acad. Sci. USA 1981, 78, 1976–1979. [Google Scholar] [CrossRef]
- Traniello, J.F.A.; Busher, C. Chemical regulation of polyethism during foraging in the neotropical termite Nasutitermes costalis. J. Chem. Ecol. 1985, 11, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Gazai, V.; Bailez, O.; Viana-Bailez, A.M. Mechanisms of trail following by the arboreal termite Nasutiters corniger (Isoptera: Termitidae). Zool. Sci. 2014, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Casarin, F.E.; Costa-Leonardo, A.M.; Arab, A. Soldiers initiate foraging activities in the subterranean termite, Heterotermes tenuis. J. Insect Sci. 2008, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Olugbemi, B.O. Exploratory and recruitment phases in soldier-mediated foraging activities in the termite, Coptotermes intermedius Silvestri (Rhinotermitidae: Coptotermitinae). Bull. Entomol. Res. 2011, 101, 423–427. [Google Scholar] [CrossRef]
- Li, H.-F.; Ye, W.; Su, N.-Y.; Kanzaki, N. Phylogeography of Coptotermes gestroi and Coptotermes formosanus (Isoptera: Rhinotermitidae) in Taiwan. Ann. Entomol. Soc. Am. 2009, 102, 684–693. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- King, E.G., Jr.; Spink, W.T. Foraging galleries of the Formosan subterranean termite, Coptotermes formosanus, in Louisiana. Ann. Entomol. Soc. Am. 1969, 62, 536–542. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Saxton, A.M.; La Fage, J.P. Foraging phenology of the Formosan subterranean termite (Isoptera: Rhinotermitidae) in Louisiana. Am. Midl. Nat. 1991, 125, 222–230. [Google Scholar] [CrossRef]
- Su, N.-Y.; Fage, J.P. Forager proportions and caste compositions of colonies of the Formosan subterranean termite (Isoptera: Rhinotermitidae) restricted to cypress trees in the Calcasieu River, Lake Charles, Louisiana. Sociobiology 1999, 33, 185–193. [Google Scholar]
- Waller, D.A.; La Fage, J.P. Unpalatability as a passive defense of Coptotermes formosanus Shiraki soldiers against ant predation. J. Appl. Entomol. 1987, 103, 148–153. [Google Scholar] [CrossRef]
- Gautam, B.K.; Henderson, G. Effects of sand moisture level on food consumption and distribution of Formosan subterranean termites (Isoptera: Rhinotermitidae) with different soldier proportions. J. Entomol. Sci. 2011, 46, 1–13. [Google Scholar] [CrossRef]
- Chouvenc, T.; Bardunias, P.; Li, H.-F.; Elliott, M.L.; Su, N.-Y. Planar arenas for use in laboratory bioassay studies of subterranean termites (Rhinotermitidae). Fla. Entomol. 2011, 94, 817–826. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa—Residual Diagnostics for HierARchical Models. GitHub—Florianhartig/DHARMa: Diagnostics for HierArchical Regession Models. Available online: https://github.com/florianhartig/DHARMa (accessed on 16 January 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R companion To Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- R Core Team. R. A language and environment for statistical computing. MSOR Connect. 2014, 1, 1–3872. [Google Scholar]
- Su, N.-Y.; Scheffrahn, R.H. Foraging population and territory of the Formosan subterranean termite (Isoptera: Rhinotermitidae) in an urban environment. Sociobiology 1988, 14, 353–360. [Google Scholar]
- Su, N.Y.; Ban, P.M.; Scheffrahn, R.H. Foraging populations and territories of the eastern subterranean termite (Isoptera: Rhinotermitidae) in southeastern Florida. Environ. Entomol. 1993, 22, 1113–1117. [Google Scholar] [CrossRef]
- Suiter, D.R.; Jones, S.C.; Forschler, B.T. Biology of Subterranean Termites in the Eastern United States; University of Georgia Bulletin: Athens, GA, USA, 2009; Volume 1209. [Google Scholar]
- Grace, J.K.; Aihara-Sasaki, M.; Yates, J.R. Differences in tunneling behavior of Coptotermes vastator and Coptotermes formosanus (Isoptera: Rhinotermitidae). Sociobiology 2004, 43, 153–158. [Google Scholar]
- Puche, H.; Su, N. Tunnel formation by Reticulitermes flavipes and Coptotermes formosanus (Isoptera: Rhinotermitidae) in response to wood in sand. J. Econ. Entomol. 2001, 94, 1398–1404. [Google Scholar] [CrossRef]
- Hapukotuwa, N.K.; Grace, J.K. Do tunnel patterns of Coptotermes formosanus and Coptotermes gestroi (Blattodea: Rhinotermitidae) reflect different foraging strategies? Sociobiology 2012, 59, 189–202. [Google Scholar] [CrossRef]
- Wells, J.D.; Henderson, G. Fire ant predation on native and introduced subterranean termites in the laboratory: Effect of high soldier number in Coptotermes formosanus. Ecol. Entomol. 1993, 18, 270–274. [Google Scholar] [CrossRef]
- Cornelius, M.; Grace, J.K. Behavioral responses of the Formosan subterranean termite (Isoptera: Rhinotermitidae) to semiochemicals of seven ant species. Phys. Chem. Ecol. 1994, 23, 1524–1528. [Google Scholar] [CrossRef]
- Konishi, T.; Matsuura, K. Royal presence promotes worker and soldier aggression against non-nestmates in termites. Insectes Soc. 2021, 68, 15–21. [Google Scholar] [CrossRef]
- Blumenfeld, A.J.; Eyer, P.A.; Husseneder, C.; Mo, J.; Johnson, L.N.; Wang, C.; Grace, J.K.; Wang, T.C.S.; Vargo, E.L. Bridgehead effect and multiple introductions shape the global invasion history of a termite. Commun. Biol. 2021, 4, 196. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P.; Williams, P.H.; Gaston, K.J. Explaining global termite diversity: Productivity or history? Biodivers. Conserv. 1994, 3, 318–330. [Google Scholar] [CrossRef]
- Guénard, B. Forum The global ant biodiversity informatics (GABI) database: Synthesizing data on the geographic distribution of ant species (Hymenoptera: Formicidae). Myrm. News 2017, 24, 83–89. [Google Scholar]
- Mitaka, Y.; Mori, N.; Matsuura, K. Multi-functional roles of a soldier-specific volatile as a worker arrestant, primer pheromone and an antimicrobial agent in a termite. Proc. R. Soc. B 2017, 284, 11–34. [Google Scholar] [CrossRef]
- Park, Y.I.; Raina, A.K. Juvenile hormone III titers and regulation of soldier caste in Coptotermes formosanus (Isoptera: Rhinotermitidae). J. Insect Phys. 2004, 50, 561–566. [Google Scholar] [CrossRef]
- Haverty, M.; Grace, J.K.; Nelson, J.L.; Yamamoto, R.T. Intercaste, intercolony, and temporal variation in cuticular hydrocarbons of Coptotermes formosanus Shiraki (Isoptera: Rhinotermitidae). J. Chem. Ecol. 1996, 22, 1813–1834. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCarthy, J.; Khadka, A.; Hakanoglu, H.; Sun, Q. Influence of Soldiers on Exploratory Foraging Behavior in the Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae). Insects 2023, 14, 198. https://doi.org/10.3390/insects14020198
McCarthy J, Khadka A, Hakanoglu H, Sun Q. Influence of Soldiers on Exploratory Foraging Behavior in the Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae). Insects. 2023; 14(2):198. https://doi.org/10.3390/insects14020198
Chicago/Turabian StyleMcCarthy, Joseph, Arjun Khadka, Hasim Hakanoglu, and Qian Sun. 2023. "Influence of Soldiers on Exploratory Foraging Behavior in the Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae)" Insects 14, no. 2: 198. https://doi.org/10.3390/insects14020198
APA StyleMcCarthy, J., Khadka, A., Hakanoglu, H., & Sun, Q. (2023). Influence of Soldiers on Exploratory Foraging Behavior in the Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae). Insects, 14(2), 198. https://doi.org/10.3390/insects14020198