
Prediction of the Overseas Migration of the Fall Armyworm, Spodoptera frugiperda, to Japan


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Predictoin Method
2.2. Evaluation Method
3. Results
4. Discussion
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, X.-X.; Hu, C.-X.; Jia, H.-R.; Wu, Q.-L.; Shen, X.-J.; Zhao, S.-Y.; Jiang, Y.-Y.; Wu, K.-M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading in China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Wu, Q.-L.; He, L.-M.; Shen, X.-J.; Jiang, Y.-Y.; Liu, J.; Hu, G.; Wu, K.-M. Estimation of the potential infestation area of newly invaded fall armyworm Spodoptera frugiperda in the Yangtze River Valley of China. Insects 2019, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-F.; Qi, G.-J.; Chen, H.; Ma, J.; Liu, J.; Jiang, Y.-Y.; Lee, G.-S.; Otuka, A.; Hu, G. Overseas immigration of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), invading Korea and Japan in 2019. Insect Sci. 2022, 29, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-M.; Song, Y.-F.; Sun, X.-X.; Shen, X.-J.; Wu, Q.-L.; Zhang, H.-W.; Zhang, D.-D.; Zhao, S.-Y.; Liang, G.-M.; Wu, K.-M. Population occurrence of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in the winter season of China. J. Integr. Agric. 2021, 20, 772–782. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.-L.; Zhang, H.-W.; Wu, K.-M. Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Watanabe, T.; Seino, H.; Kitamura, C.; Hirai, Y. A computer program, LLJET, utilizing an 850 mb weather chart to forecast long-distance rice planthopper migration. Bull. Kyushu Natl. Agric. Exp. Stn. 1990, 26, 233–260. (In Japanese) [Google Scholar]
- Otuka, A.; Watanabe, T.; Suzuki, Y.; Matsumura, M.; Furuno, A.; Chino, M. Real-time prediction system for migration of rice planthoppers Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Appl. Entomol. Zool. 2005, 40, 221–229. [Google Scholar] [CrossRef]
- Otuka, A.; Zhou, Y.; Lee, G.-S.; Matsumura, M.; Zhu, Y.; Park, H.-H.; Liu, Z.; Sanada-Morimura, S. Prediction of overseas migration of the small brown planthopper, Laodelphax striatellus (Hemiptera: Delphacidae) in East Asia. Appl. Entomol. Zool. 2012, 47, 379–388. [Google Scholar] [CrossRef]
- Otuka, A. JPP-NET migration prediction system for the small brown planthopper in East Asia. Plant Prot. 2016, 70, 116–120. (In Japanese) [Google Scholar]
- Wu, Q.-L.; Shen, X.-J.; He, L.-M.; Jiang, Y.-Y.; Liu, J.; Hu, G.; Wu, K.-M. Windborne migration routes of newly-emerged fall armyworm from Qinling Mountains-Huaihe River region, China. J. Integr. Agric. 2020, 19, 2–14. [Google Scholar] [CrossRef]
- Wu, Q.-L.; Jiang, Y.-Y.; Liu, J.; Hu, G.; Wu, K.-M. Trajectory modeling revealed a southwest-northeast migration corridor for fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) emerging from the North China. Insect Sci. 2021, 28, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-Y.; Wu, Q.-L.; Jia, H.-R.; Wu, K.-M. Searchlight trapping reveals seasonal cross-ocean migration of fall armyworm over the South China Sea. J. Integr. Agric. 2021, 20, 673–684. [Google Scholar] [CrossRef]
- Westbrook, J.; Fleischer, S.; Jairam, S.; Meagher, R.; Nagoshi, R. Multigenerational migration of fall armyworm, a pest insect. Ecosphere 2019, 10, e02919. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Drake, V.A.; Reynolds, D.R. Radar Entomology: Observing Insect Flight and Migration; CABI: Oxfordshire, UK, 2012. [Google Scholar]
- Chapman, J.W.; Nesbit, R.L.; Burgin, L.E.; Reynolds, D.R.; Smith, A.D.; Middleton, D.R.; Hill, J.K. Flight Orientation Behaviors Promote Optimal Migration Trajectories in High-Flying Insects. Science 2010, 327, 682–685. [Google Scholar] [CrossRef]
- Skamarock, W.C.; Klemp, J.B.; Dudhia, J.; Gill, D.O.; Liu, Z.; Berner, J.; Wang, W.; Powers, J.G.; Duda, M.G.; Barker, D.; et al. A Description of the Advanced Research WRF Model Version 4.3; NCAR: Boulder, CO, USA, 2021. [Google Scholar] [CrossRef]
- Wang, W. An Introduction to the WRF Modelling System; MMM, NCAR: Boulder, CO, USA, 2021; Available online: https://www2.mmm.ucar.edu/wrf/users/tutorial/presentation_pdfs/202101/wang_intro.pdf (accessed on 28 March 2023).
- NCAR. User’s Guide for the Advanced Research WRF (ARW) Modeling System Version 4.3; NCAR: Boulder, CO, USA, 2021; Available online: https://www2.mmm.ucar.edu/wrf/users/docs/user_guide_v4/v4.3/contents.html (accessed on 28 March 2023).
- Terada, H.; Nagai, H.; Furuno, A.; Kakefuda, T.; Harayama, T.; Chino, M. Development of Worldwide Version of System for Prediction of Environmental Emergency Dose Information: WSPEEDI 2nd Version. Trans. At. Energy Soc. Jpn. 2008, 7, 257–267. (In Japanese) [Google Scholar] [CrossRef]
- Terada, H.; Chino, M. Development of an atmospheric dispersion model for accidental discharge of radionuclides with the function of simultaneous prediction for multiple domains and its evaluation by application to the Chernobyl nuclear accident. J. Nucl. Sci. Technol. 2008, 45, 920–931. [Google Scholar] [CrossRef]
- JAEA. User’s Manual for WSPEEDI-II (Ver. 2.2.0); Research Group for Environmental Science, JAEA Nuclear Science and Engineering Center: Tokai, Japan, 2020; p. 24. [Google Scholar]
- Furuno, A.; Chino, M.; Yamazawa, H. Worldwide Version of System for Prediction of Environmental Emergency Dose Information (WSPEEDI) Model Code (II) –Three-Dimensional Atmospheric Dispersion Model for Synoptic Scale: GEARN–; JAERI: Tokai, Japan, 1999; JAERI-Data/Code 99-044; p. 68. (In Japanese)
- Furuno, A.; Chino, M.; Otuka, A.; Watanabe, T.; Matsumura, M.; Suzuki, Y. Development of a numerical simulation model for long-range migration of rice planthoppers. Agric. For. Meteorol. 2005, 133, 197–209. [Google Scholar] [CrossRef]
- Terada, H.; Chino, M.; Furuno, A. Improvement of worldwide version of system for prediction of environmental emergency dose information (WSPEEDI), (I); New combination of models, atmospheric dynamic model MM5 and particle random walk model GEARN-new. J. Nucl. Sci. Technol. 2004, 41, 632–640. [Google Scholar] [CrossRef]
- Wainwright, C.; Reynolds, D.R.; Reynolds, A.M. Linking small-scale flight manoeuvers and density profiles to the vertical movement of insects in the nocturnal stable boundary layer. Sci. Rep. 2020, 10, 1019. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Huang, L.; Xu, C.-F.; Li, J.-H.; Wang, F.-Y.; Cheng, W.; Gao, B.-Y.; Chapman, J.W.; Hu, G. Flight Capability and the Low Temperature Threshold of a Chinese Field Population of the Fall Armyworm Spodoptera frugiperda. Insects 2022, 13, 422. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Huang, L.; Liu, J.; Zhang, H.-B.; Qiu, K.; Lu, F.; Hu, G. Migration Dynamics of Fall Armyworm Spodoptera frugiperda (Smith) in the Yangtze River Delta. Insects 2023, 14, 127. [Google Scholar] [CrossRef] [PubMed]
- Tojo, S.; Ryuda, M.; Fukuda, T.; Matsunaga, T.; Choi, D.-R.; Otuka, A. Overseas migration of the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae), from May to mid-July in East Asia. Appl. Entomol. Zool. 2013, 48, 131–140. [Google Scholar] [CrossRef]
- JMA (Japan Meteorological Agency). Climatological Normal of the Hitting Ratio of Rainfall Forecast. 2023. Available online: https://www.data.jma.go.jp/fcd/yoho/kensho/reinen.html (accessed on 12 May 2023). (In Japanese)
- Otuka, A.; Watanabe, T.; Suzuki, Y.; Matsumura, M.; Furuno, A.; Chino, M.; Kondo, T.; Kamimuro, T. A migration analysis of Sogatella furcifera (Horváth) (Homoptera: Delphacidae) using hourly catches and a three-dimensional simulation model. Agric. For. Entomol. 2005, 8, 35–47. [Google Scholar] [CrossRef]
- Chapman, J.W.; Reynolds, D.R.; Mouritsen, H.; Hill, J.K.; Riley, J.R.; Sivell, D.; Smith, A.D.; Woiwod, I.P. Wind selection and drift compensation optimize migratory pathways in a high-flying moth. Curr. Biol. 2008, 18, 514–518. [Google Scholar] [CrossRef]
- Wolf, W.W.; Westbrook, J.K.; Raulston, J.R.; Pair, S.D.; Lingren, P.D. Radar observations of orientation of noctuids migrating from corn fields in the Lower Rio Grande Valley. Southwest. Entomol. Suppl. 1995, 18, 45–61. [Google Scholar]
- Drake, V.A. Radar observations of moths migrating in a nocturnal low-level jet. Ecol. Entomol. 1985, 10, 259–265. [Google Scholar] [CrossRef]
- Houston Center Weather Service Unit. The Low Level Jet. 2023. Available online: https://www.weather.gov/source/zhu/ZHU_Training_Page/Miscellaneous/lowleveljet/lowleveljet.html (accessed on 24 September 2023).
- Sato, K.; Manda, A.; Moteki, Q.; Komatsu, K.; Ogata, K.; Nishikawa, H.; Oshika, M.; Otomi, Y.; Kunoki, S.; Kanehara, H.; et al. Influence of the Kuroshio on Mesoscale Convective Systems in the Baiu Frontal Zone over the East China Sea. Mon. Weather Rev. 2016, 144, 1017–1033. [Google Scholar] [CrossRef]
- Koyama, J.; Matsumura, M. Ecology and control of armyworm, Mythimna separata (Lepidoptera: Noctuidae) in Japan, with special reference to outbreak and migration. Jpn. J. Appl. Entomol. Zool. 2019, 63, 39–56. (In Japanese) [Google Scholar] [CrossRef]
- SPPS (Shimane Prefectural Plant Protection Station). Pest Occurrence Forecast Technical Note No. 4, Occurrence of the Oriental Armyworm [2017-06-29]; Shimane Prefecture: Izumo, Japan, 2017. (In Japanese) [Google Scholar]
- TPPS (Tottori Prefectural Plant Protection Station). Advisory Information of Insect Pest Occurrence: Oriental Armyworm [2017-06-30]; Tottori Prefecture: Tottori, Japan, 2017. (In Japanese) [Google Scholar]
- YPPS (Yamaguchi Prefectural Plant Protection Station). Pest Occurrence Forecast Technical Note No. 2, Outbreak of the Oriental Armyworm and Its Control [2017-07-04]; Yamaguchi Prefecture: Yamaguchi, Japan, 2017. (In Japanese) [Google Scholar]
- OPPS (Okayama Prefectural Plant Protection Station). Plant Protection Information No. 2, Occurrence of the Oriental Armyworm [2017-07-06]; Okayama Prefecture: Okayama, Japan, 2017. (In Japanese) [Google Scholar]
- TPG (Tottori Prefectural Government). Damage Caused by the Oriental Armyworm Outbreak in 2017 and Its Measure; Tottori Prefecture: Tottori, Japan, 2017. (In Japanese) [Google Scholar]
- Drake, A.V. The vertical distribution of macro-insects migrating in the nocturnal boundary layer: A radar study. Bound. Layer Meteorol. 1984, 28, 353–374. [Google Scholar] [CrossRef]
- Feng, H.-Q.; Zhao, X.-C.; Wu, X.-F.; Wu, B.; Wu, K.-M.; Cheng, D.-F.; Guo, Y.-Y. Autumn migration of Mythimna separata (Lepidoptera: Noctuidae) over the Bohai Sea in Northern China. Environ. Entomol. 2008, 37, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Otuka, A.; Matsumura, M.; Tokuda, M. Development of method to forecast soybean leaf damage by common cutworm using entomological radar and searchlight trap. J. Eng. 2019, 2019, 7531–7533. [Google Scholar] [CrossRef]
- Hammer, I.T.; Drake, V.A. Insect monitoring radar: Analytical time-domain algorithm for retrieving trajectory and target parameters, Comput. Electron. Agric. 2004, 43, 23–41. [Google Scholar] [CrossRef]
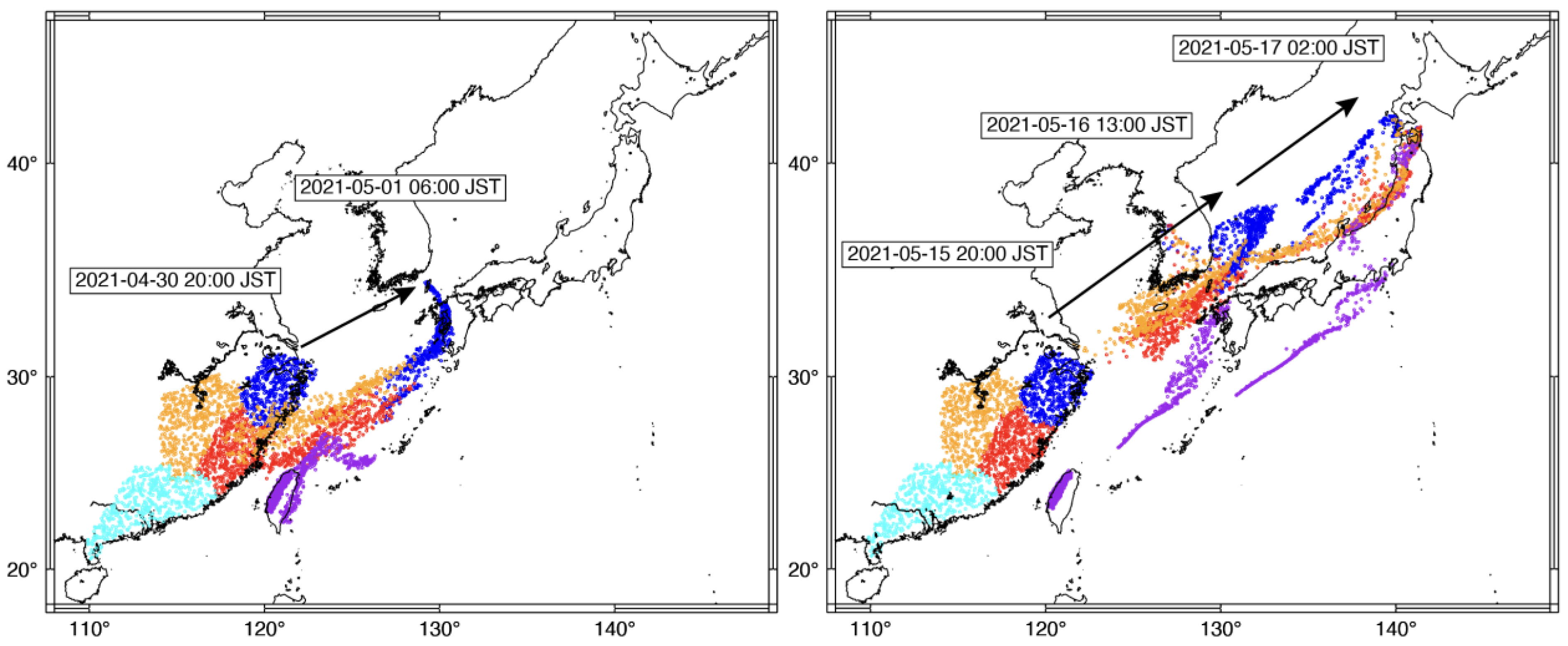

{kind=link}
| Parameter/Data | Values |
|---|---|
| WRF parameters: | |
| domain size | 151, 131 |
| number of domains | 1 |
| horizontal resolution (dx, dy) | 27 km |
| map projection | Mercator |
| projection reference latitude | 32.0 degree |
| projection reference longitude | 128.0 degree |
| forecast time | 72 h |
| time step | 120 s |
| microphysics | Thompson scheme |
| cumulus parameterization | Tiedtke scheme |
| longwave radiation | RRTMG scheme |
| shortwave radiation | RRTMG shortwave |
| planetary boundary layer | Mellor-Yamada-Janjic scheme |
| surface layer | Eta similarity |
| land surface | Noah Land Surface Model |
| analysis nudging | applied |
| Meteorological data: | |
| name | NOAA Global Forecast System (GFS) |
| time interval | 6 h |
| horizontal resolution | 0.25 degree |
| number of vertical layers | 23 layers up to p_top |
| pressure top (p_top) | 50 hPa |
| URL for downloading | https://registry.opendata.aws/noaa-gfs-bdp-pds/ (accessed on 1 August 2023) |
| Mapping software: | |
| name | The General Mapping Tools Ver. 6.3.0 |
| 2020 | 2021 |
|---|---|
| Minami-satsuma (31.48° N, 130.34° E) | Minami-satsuma (31.48° N, 130.34° E) |
| Koshi (32.88° N, 130.74° E) | Koshi (32.88° N, 130.74° E) |
| Isahaya (32.83° N, 130.02° E) | Isahaya (32.83° N, 130.02° E) |
| Nangoku (33.59° N, 133.64° E) | Nangoku (33.59° N, 133.64° E) |
| Izumo (35.33° N, 132.75° E) | Nagakute (35.16° N, 137.08° E) |
| Towada (40.62° N, 141.16° E) | Towada (40.62° N, 141.16° E) |
| Date of Collection/ Prediction | Trap Location: Koshi | Isahaya | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Prediction 1 (Source) | FT 2 (h) | Trap Catch | Eval. 3-Day 3 | Eval. 5-Day 3 | Prediction (Source) | FT (h) | Trap Catch 4 | Eval. 3-Day | Eval. 5-Day | |
| 1 May 2021 | Yes (ZJ) | 12 | 0 | H | H | Yes (ZJ) | 11 | 0 | F | F |
| 2 May 2021 | No | 34 | - | - | No | - | - | |||
| 3 May 2021 | No | 38 | - | - | No | - | - | |||
| 4 May 2021 | Yes (JX) | 38 | 18 | H | H | Yes (GD) | 35 | F | F | |
| 5 May 2021 | No | 9 | - | - | No | 0 | - | - | ||
| 6 May 2021 | Yes (TW) | 35 | 17 | H | H | Yes (TW) | 33 | F | F | |
| 7 May 2021 | Yes (ZJ) | 29 | 22 | H | H | Yes (ZJ) | 24 | 0 | F | F |
| 8 May 2021 | Yes (ZJ) | 23 | 3 | H | H | Yes (ZJ) | 21 | F | F | |
| 9 May 2021 | Yes (ZJ) | 37 | 9 | H | H | Yes (ZJ) | 32 | F | F | |
| 10 May 2021 | Yes (FJ) | 36 | 13 | H | H | Yes (FJ) | 34 | F | F | |
| 11 May 2021 | No | 0 | - | - | No | - | - | |||
| 12 May 2021 | Yes (JX) | 35 | 2 | H | H | Yes (ZJ) | 26 | 0 | F | H |
| 13 May 2021 | Yes (FJ) | 29 | 3 | H | H | Yes (FJ) | 27 | F | H | |
| 14 May 2021 | Yes (FJ) | 32 | 1 | F | H | Yes (FJ) | 29 | H | H | |
| 15 May 2021 | Yes (ZJ) | 20 | 0 | F | H | Yes (ZJ) | 17 | H | H | |
| 16 May 2021 | Yes (TW) | 20 | 0 | H | H | Yes (TW) | 18 | H | H | |
| 17 May 2021 | No | 0 | - | - | No | 7 | - | - | ||
| 18 May 2021 | No | 0 | - | - | No | 0 | - | - | ||
| 19 May 2021 | Yes (ZJ) | 30 | 1 | H | H | Yes (FJ) | 31 | 0 | H | H |
| 20 May 2021 | Yes (TW) | 20 | 0 | H | H | Yes (GD) | 32 | 2 | H | H |
| 21 May 2021 | No | 0 | - | - | No | 19 | - | - | ||
| 22 May 2021 | No | 3 | - | - | No | 3 | - | - | ||
| 23 May 2021 | Yes (TW) | 36 | 0 | H | H | Yes (ZJ) | 36 | 2 | H | H |
| 24 May 2021 | No | 1 | - | - | No | 3 | - | - | ||
| 25 May 2021 | No | 0 | - | - | No | 7 | - | - | ||
| 26 May 2021 | Yes (ZJ) | 11 | 0 | F | F | Yes (TW) | 30 | H | H | |
| 27 May 2021 | No | 0 | - | - | No | 2 | - | - | ||
| 28 May 2021 | Yes (ZJ) | 23 | 0 | H | H | Yes (ZJ) | 20 | 0 | H | H |
| 29 May 2021 | No | 0 | - | - | No | 3 | - | - | ||
| 30 May 2021 | No | 0 | - | - | No | 0 | - | - | ||
| 31 May 2021 | No | 1 | F | - | No | F | - | |||
| Location | Hitting Ratio | |
|---|---|---|
| 3-Day Period | 5-Day Period | |
| 2020: | ||
| Minami-satsuma | 0.67 | 0.88 |
| Koshi | 0.76 | 0.93 |
| Isahaya | 0.89 | 0.89 |
| Nangoku | 0.64 | 0.81 |
| Izumo | 0.83 | 0.76 |
| Towada | 0.96 | 0.95 |
| Yearly avg. | 0.79 | 0.87 |
| 2021: | ||
| Minami-satsuma | 0.83 | 1.00 |
| Koshi | 0.78 | 0.94 |
| Isahaya | 0.44 | 0.59 |
| Nangoku | 0.79 | 0.87 |
| Izumo | 0.84 | 0.75 |
| Towada | 0.91 | 0.95 |
| Yearly avg. | 0.77 | 0.85 |
| Total avg. | 0.78 | 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otuka, A. Prediction of the Overseas Migration of the Fall Armyworm, Spodoptera frugiperda, to Japan. Insects 2023, 14, 804. https://doi.org/10.3390/insects14100804
Otuka A. Prediction of the Overseas Migration of the Fall Armyworm, Spodoptera frugiperda, to Japan. Insects. 2023; 14(10):804. https://doi.org/10.3390/insects14100804
Chicago/Turabian StyleOtuka, Akira. 2023. "Prediction of the Overseas Migration of the Fall Armyworm, Spodoptera frugiperda, to Japan" Insects 14, no. 10: 804. https://doi.org/10.3390/insects14100804
APA StyleOtuka, A. (2023). Prediction of the Overseas Migration of the Fall Armyworm, Spodoptera frugiperda, to Japan. Insects, 14(10), 804. https://doi.org/10.3390/insects14100804