

Selection and Comparative Gene Expression of Midgut-Specific Targets for Drosophila suzukii


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Isolation of Total RNA
2.3. Illumina Sequencing
2.4. Anlaysis of up- or down-Regulated Genes
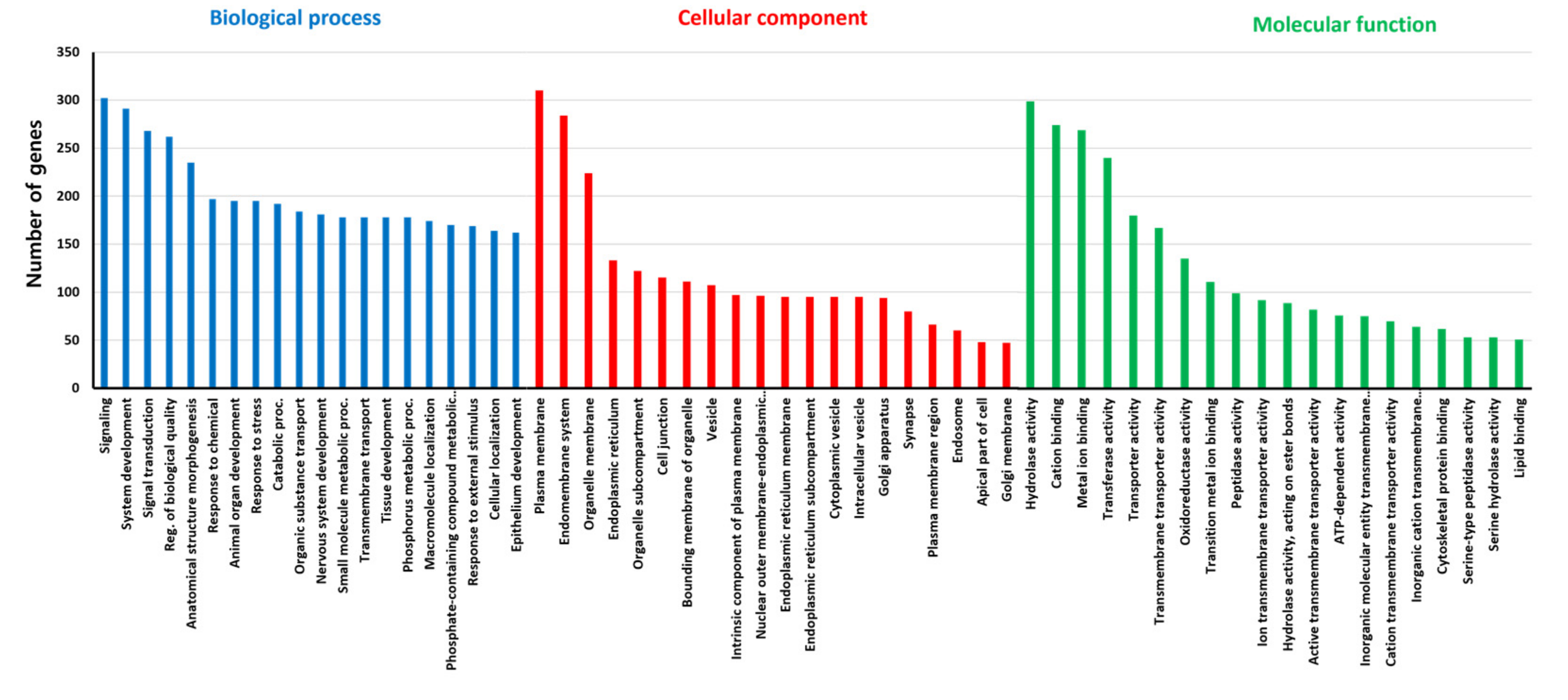
2.5. Functional Annotation and Gene Ontology Analysis
2.6. Manual Annotation of the Midgut Transcriptome
2.7. Quantitative Real-Time PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results and Discussion
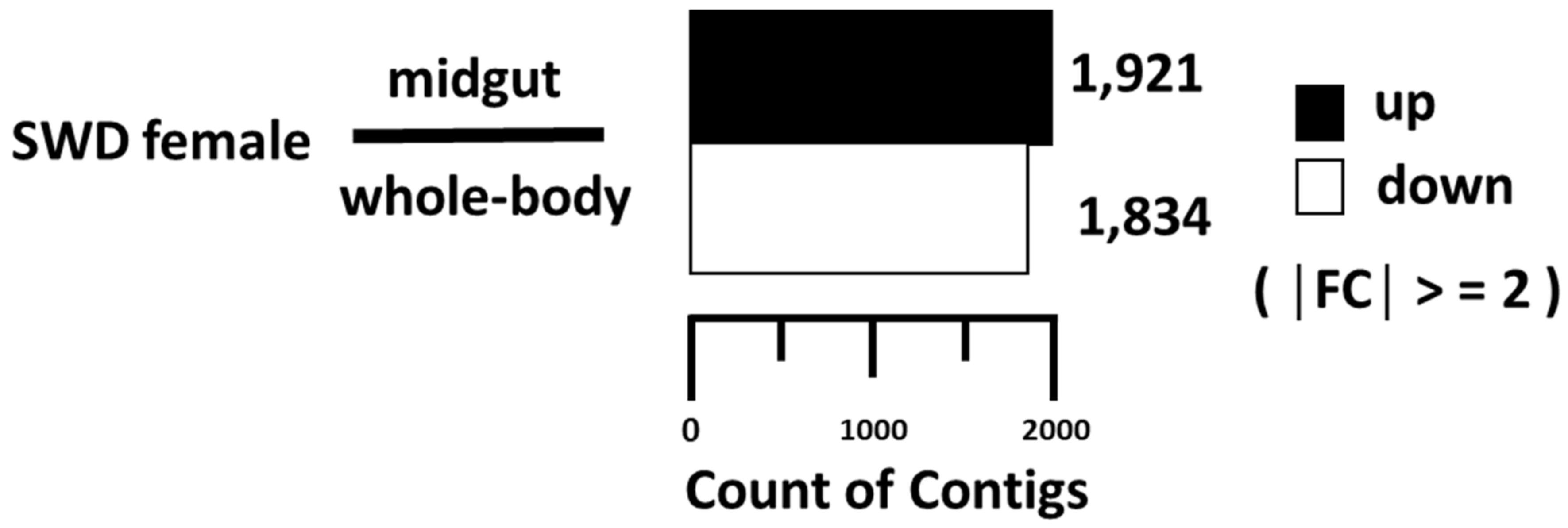
3.1. Midgut Transcriptome Analysis
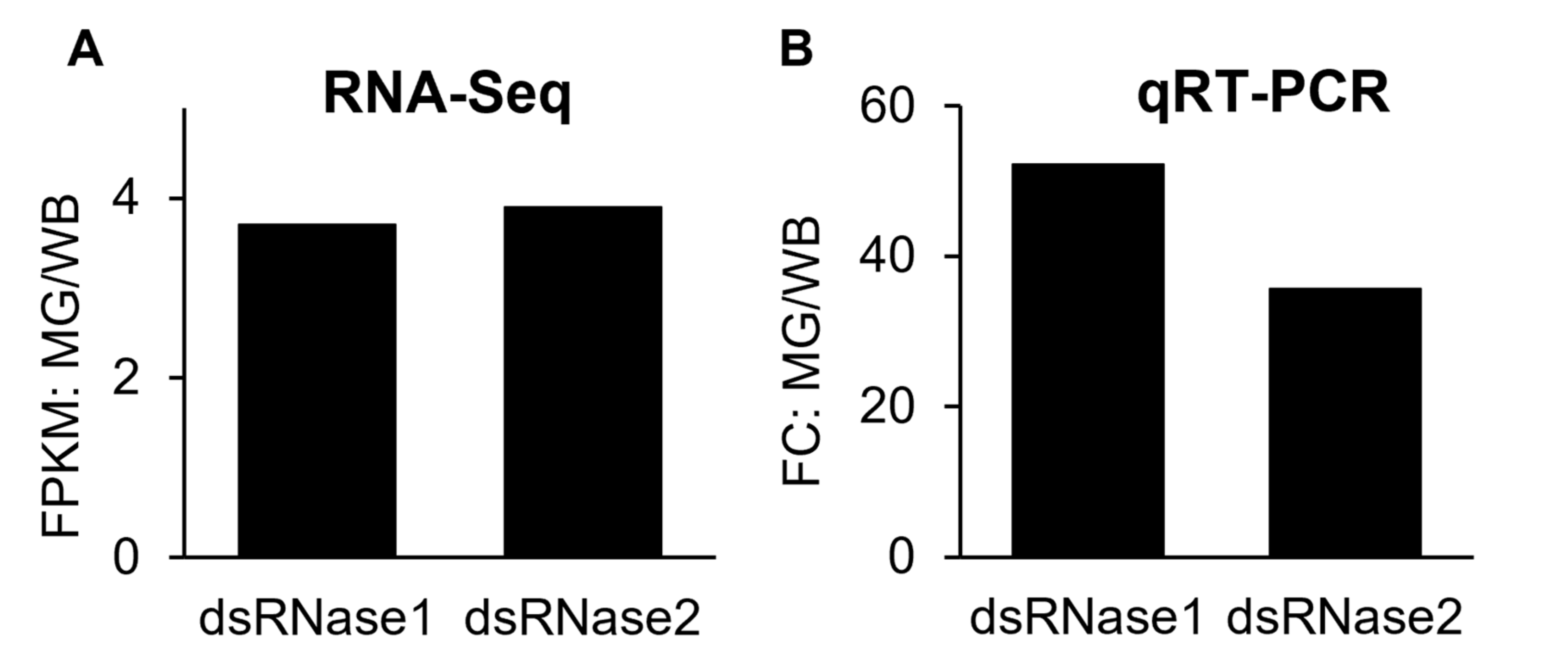
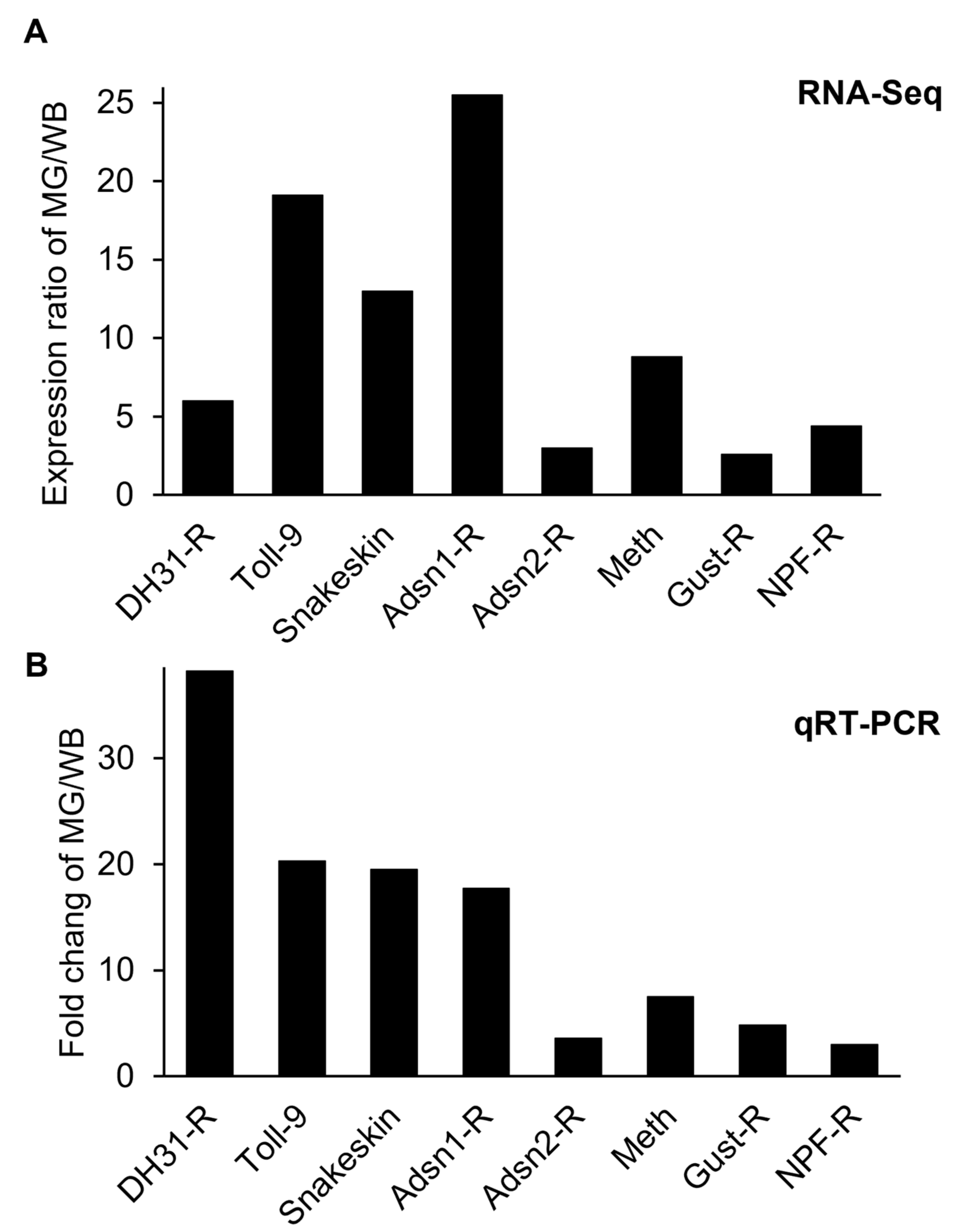
3.2. Validation of the Midgut-Specifically Upregulated Genes
3.3. Midgut-Specifically Upregulated Membrane Proteins and GPCRs
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. Biol. Sci. 2014, 281, 20132840. [Google Scholar] [CrossRef] [PubMed]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.A.; Mendes, M.F.; Kruger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R. Global potential distribution of Drosophila suzukii (Diptera, Drosophilidae). PLoS ONE 2017, 12, e0174318. [Google Scholar] [CrossRef] [PubMed]
- Klick, J.; Yang, W.Q.; Lee, J.C.; Bruck, D.J. Reduced spray programs for Drosophila suzukii management in berry crops. Int. J. Pest Manag. 2016, 62, 368–377. [Google Scholar] [CrossRef]
- Rice, K.B.; Short, B.D.; Leskey, T.C. Development of an Attract-and-Kill Strategy for Drosophila suzukii (Diptera: Drosophilidae): Evaluation of Attracticidal Spheres Under Laboratory and Field Conditions. J. Econ. Entomol. 2017, 110, 535–542. [Google Scholar] [CrossRef]
- Chiu, J.C.; Jiang, X.; Zhao, L.; Hamm, C.A.; Cridland, J.M.; Saelao, P.; Hamby, K.A.; Lee, E.K.; Kwok, R.S.; Zhang, G.; et al. Genome of Drosophila suzukii, the spotted wing drosophila. G3 2013, 3, 2257–2271. [Google Scholar] [CrossRef]
- Crava, C.M.; Sassu, F.; Tait, G.; Becher, P.G.; Anfora, G. Functional transcriptome analyses of Drosophila suzukii antennae reveal mating-dependent olfaction plasticity in females. Insect Biochem. Mol. Biol. 2019, 105, 51–59. [Google Scholar] [CrossRef]
- Ahn, S.J.; Oh, H.W.; Corcoran, J.; Kim, J.A.; Park, K.C.; Park, C.G.; Choi, M.Y. Sex-biased gene expression in antennae of Drosophila suzukii. Arch. Insect Biochem. Physiol. 2020, 104, e21660. [Google Scholar] [CrossRef]
- Deng, D.; Xing, S.; Liu, X.; Ji, Q.; Zhai, Z.; Peng, W. Transcriptome analysis of sex-biased gene expression in the spotted-wing Drosophila, Drosophila suzukii (Matsumura). G3 2022, 12, jkac127. [Google Scholar] [CrossRef]
- Paris, M.; Boyer, R.; Jaenichen, R.; Wolf, J.; Karageorgi, M.; Green, J.; Cagnon, M.; Parinello, H.; Estoup, A.; Gautier, M.; et al. Near-chromosome level genome assembly of the fruit pest Drosophila suzukii using long-read sequencing. Sci. Rep. 2020, 10, 11227. [Google Scholar] [CrossRef]
- Tait, G.; Mermer, S.; Stockton, D.; Lee, J.; Avosani, S.; Abrieux, A.; Anfora, G.; Beers, E.; Biondi, A.; Burrack, H.; et al. Drosophila suzukii (Diptera: Drosophilidae): A decade of research towards a sustainable integrated pest management program. J. Econ. Entomol. 2021, 114, 1950–1974. [Google Scholar] [CrossRef]
- Nassel, D.R.; Zandawala, M. Recent advances in neuropeptide signaling in Drosophila, from genes to physiology and behavior. Prog. Neurobiol. 2019, 179, 101607. [Google Scholar] [CrossRef]
- Nassel, D.R.; Zandawala, M. Endocrine cybernetics: Neuropeptides as molecular switches in behavioural decisions. Open Biol. 2022, 12, 220174. [Google Scholar] [CrossRef]
- Audsley, N.; Down, R.E. G protein coupled receptors as targets for next generation pesticides. Insect Biochem. Mol. Biol. 2015, 67, 27–37. [Google Scholar] [CrossRef]
- Caers, J.; Verlinden, H.; Zels, S.; Vandersmissen, H.P.; Vuerinckx, K.; Schoofs, L. More than two decades of research on insect neuropeptide GPCRs: An overview. Front. Endocrinol. 2012, 3, 151. [Google Scholar] [CrossRef]
- Yeoh, J.G.C.; Pandit, A.A.; Zandawala, M.; Nassel, D.R.; Davies, S.A.; Dow, J.A.T. DINeR: Database for Insect Neuropeptide Research. Insect Biochem. Mol. Biol. 2017, 86, 9–19. [Google Scholar] [CrossRef]
- Murphy, K.A.; Tabuloc, C.A.; Cervantes, K.R.; Chiu, J.C. Ingestion of genetically modified yeast symbiont reduces fitness of an insect pest via RNA interference. Sci. Rep. 2016, 6, 22587. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Christiaens, O.; Berkvens, N.; Casteels, H.; Maes, M.; Smagghe, G. Oral RNAi to control Drosophila suzukii: Laboratory testing against larval and adult stages. J. Pest Sci. 2016, 89, 803–814. [Google Scholar] [CrossRef]
- Ahn, S.J.; Donahue, K.; Koh, Y.; Martin, R.R.; Choi, M.Y. Microbial-Based Double-Stranded RNA Production to Develop Cost-Effective RNA Interference Application for Insect Pest Management. Int. J. Insect Sci. 2019, 11, 1179543319840323. [Google Scholar] [CrossRef]
- Abrieux, A.; Chiu, J.C. Oral delivery of dsRNA by microbes: Beyond pest control. Commun. Integr. Biol. 2016, 9, e1236163. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Ahn, S.J.; Flinn, C.M.; Choi, M.Y. Identification and functional analysis of dsRNases in spotted-wing drosophila, Drosophila suzukii. Arch. Insect Biochem. Physiol. 2021, 107, e21822. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, R.; Heschuk, D.; Kim, B.; Whyard, S. A novel paperclip double-stranded RNA structure demonstrates clathrin-independent uptake in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2020, 127, 103492. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Miguel-Aliaga, I. The digestive tract of Drosophila melanogaster. Annu. Rev. Genet. 2013, 47, 377–404. [Google Scholar] [CrossRef]
- Bing, X.; Gerlach, J.; Loeb, G.; Buchon, N. Nutrient-Dependent Impact of Microbes on Drosophila suzukii Development. mBio 2018, 9, e02199-17. [Google Scholar] [CrossRef]
- Solomon, G.M.; Dodangoda, H.; McCarthy-Walker, T.; Ntim-Gyakari, R.; Newell, P.D. The microbiota of Drosophila suzukii influences the larval development of Drosophila melanogaster. PeerJ 2019, 7, e8097. [Google Scholar] [CrossRef]
- Woltz, J.M.; Donahue, K.M.; Bruck, D.J.; Lee, J.C. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 2015, 139, 759–770. [Google Scholar] [CrossRef]
- Hu, Y.; Comjean, A.; Perrimon, N.; Mohr, S.E. The Drosophila Gene Expression Tool (DGET) for expression analyses. BMC Bioinform. 2017, 18, 98. [Google Scholar] [CrossRef]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef]
- Almeida Garcia, R.; Lima Pepino Macedo, L.; Cabral do Nascimento, D.; Gillet, F.X.; Moreira-Pinto, C.E.; Faheem, M.; Moreschi Basso, A.M.; Mattar Silva, M.C.; Grossi-de-Sa, M.F. Nucleases as a barrier to gene silencing in the cotton boll weevil, Anthonomus grandis. PLoS ONE 2017, 12, e0189600. [Google Scholar] [CrossRef]
- Cooper, A.M.W.; Song, H.; Shi, X.; Yu, Z.; Lorenzen, M.; Silver, K.; Zhang, J.; Zhu, K.Y. Molecular Characterizations of Double-Stranded RNA Degrading Nuclease Genes from Ostrinia nubilalis. Insects 2020, 11, 652. [Google Scholar] [CrossRef]
- Liu, J.; Swevers, L.; Iatrou, K.; Huvenne, H.; Smagghe, G. Bombyx mori DNA/RNA non-specific nuclease: Expression of isoforms in insect culture cells, subcellular localization and functional assays. J. Insect Physiol. 2012, 58, 1166–1176. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, K.; Chen, J.; Wang, J.; Zhang, H.; Ze, L.; Zhu, G.; Zhao, C.; Xiao, H.; Han, Z. Identification of a double-stranded RNA-degrading nuclease influencing both ingestion and injection RNA interference efficiency in the red flour beetle Tribolium castaneum. Insect Biochem. Mol. Biol. 2020, 125, 103440. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, K.; Zhu, G.; Han, Q.; Chen, J.; Elzaki, M.E.A.; Sheng, C.; Zhao, C.; Palli, S.R.; Han, Z. Identification and characterization of multiple dsRNases from a lepidopteran insect, the tobacco cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2020, 162, 86–95. [Google Scholar] [CrossRef]
- Hu, X.; Richtman, N.M.; Zhao, J.Z.; Duncan, K.E.; Niu, X.; Procyk, L.A.; Oneal, M.A.; Kernodle, B.M.; Steimel, J.P.; Crane, V.C.; et al. Discovery of midgut genes for the RNA interference control of corn rootworm. Sci. Rep. 2016, 6, 30542. [Google Scholar] [CrossRef]
- Birgul Iyison, N.; Shahraki, A.; Kahveci, K.; Duzgun, M.B.; Gun, G. Are insect GPCRs ideal next-generation pesticides: Opportunities and challenges. FEBS J 2021, 288, 2727–2745. [Google Scholar] [CrossRef]
- Choi, M.Y.; Vander Meer, R.K. GPCR-Based Bioactive Peptide Screening Using Phage-Displayed Peptides and an Insect Cell System for Insecticide Discovery. Biomolecules 2021, 11, 583. [Google Scholar] [CrossRef]
- Arnot, C.J.; Gay, N.J.; Gangloff, M. Molecular mechanism that Induces activation of Spatzle, the ligand for the Drosophila Toll receptor. J. Biol. Chem. 2010, 285, 19502–19509. [Google Scholar] [CrossRef]
- Liu, J.; Smagghe, G.; Swevers, L. Transcriptional response of BmToll9-1 and RNAi machinery genes to exogenous dsRNA in the midgut of Bombyx mori. J. Insect Physiol. 2013, 59, 646–654. [Google Scholar] [CrossRef]
- Johnson, E.C.; Shafer, O.T.; Trigg, J.S.; Park, J.; Schooley, D.A.; Dow, J.A.; Taghert, P.H. A novel diuretic hormone receptor in Drosophila: Evidence for conservation of CGRP signaling. J. Exp. Biol. 2005, 208, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Zandawala, M. Calcitonin-like diuretic hormones in insects. Insect Biochem. Mol. Biol. 2012, 42, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Seroude, L.; Benzer, S. Extended life-span and stress resistance in the Drosophila mutant methuselah. Science 1998, 282, 943–946s. [Google Scholar] [CrossRef]
- Cao, C.; Sun, L.; Du, H.; Moural, T.W.; Bai, H.; Liu, P.; Zhu, F. Physiological functions of a methuselah-like G protein coupled receptor in Lymantria dispar Linnaeus. Pestic. Biochem. Physiol. 2019, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef]
- Wu, S.K.; Yap, A.S. Patterns in space: Coordinating adhesion and actomyosin contractility at E-cadherin junctions. Cell Commun. Adhes. 2013, 20, 201–212. [Google Scholar] [CrossRef]
- Garczynski, S.F.; Brown, M.R.; Shen, P.; Murray, T.F.; Crim, J.W. Characterization of a functional neuropeptide F receptor from Drosophila melanogaster. Peptides 2002, 23, 773–780. [Google Scholar] [CrossRef]
- Miyamoto, T.; Slone, J.; Song, X.; Amrein, H. A fructose receptor functions as a nutrient sensor in the Drosophila brain. Cell 2012, 151, 1113–1125. [Google Scholar] [CrossRef]
- Miyamoto, T.; Amrein, H. Diverse roles for the Drosophila fructose sensor Gr43a. Fly 2014, 8, 19–25. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.-S.; Ahn, S.-J.; Choi, M.-Y. Selection and Comparative Gene Expression of Midgut-Specific Targets for Drosophila suzukii. Insects 2023, 14, 76. https://doi.org/10.3390/insects14010076
Yoon J-S, Ahn S-J, Choi M-Y. Selection and Comparative Gene Expression of Midgut-Specific Targets for Drosophila suzukii. Insects. 2023; 14(1):76. https://doi.org/10.3390/insects14010076
Chicago/Turabian StyleYoon, June-Sun, Seung-Joon Ahn, and Man-Yeon Choi. 2023. "Selection and Comparative Gene Expression of Midgut-Specific Targets for Drosophila suzukii" Insects 14, no. 1: 76. https://doi.org/10.3390/insects14010076
APA StyleYoon, J.-S., Ahn, S.-J., & Choi, M.-Y. (2023). Selection and Comparative Gene Expression of Midgut-Specific Targets for Drosophila suzukii. Insects, 14(1), 76. https://doi.org/10.3390/insects14010076