

Developmental Differentiations of Major Maize Stemborers Due to Global Warming in Temperate and Tropical Climates


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
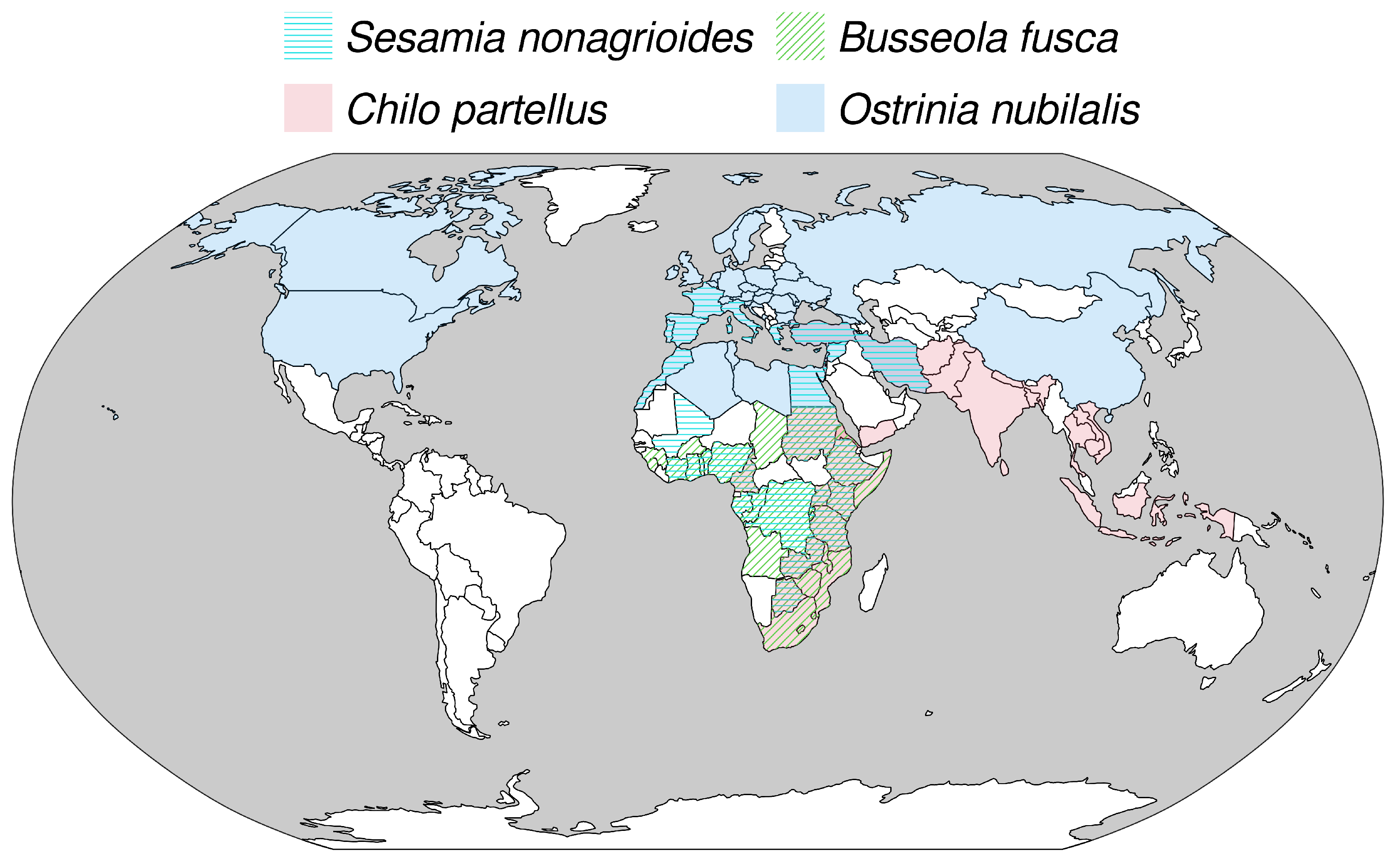
2.1. Biological Models
2.2. Thermal Performance Curves
2.3. Temperature Scenarios
2.4. Simulations of Development Times
3. Results
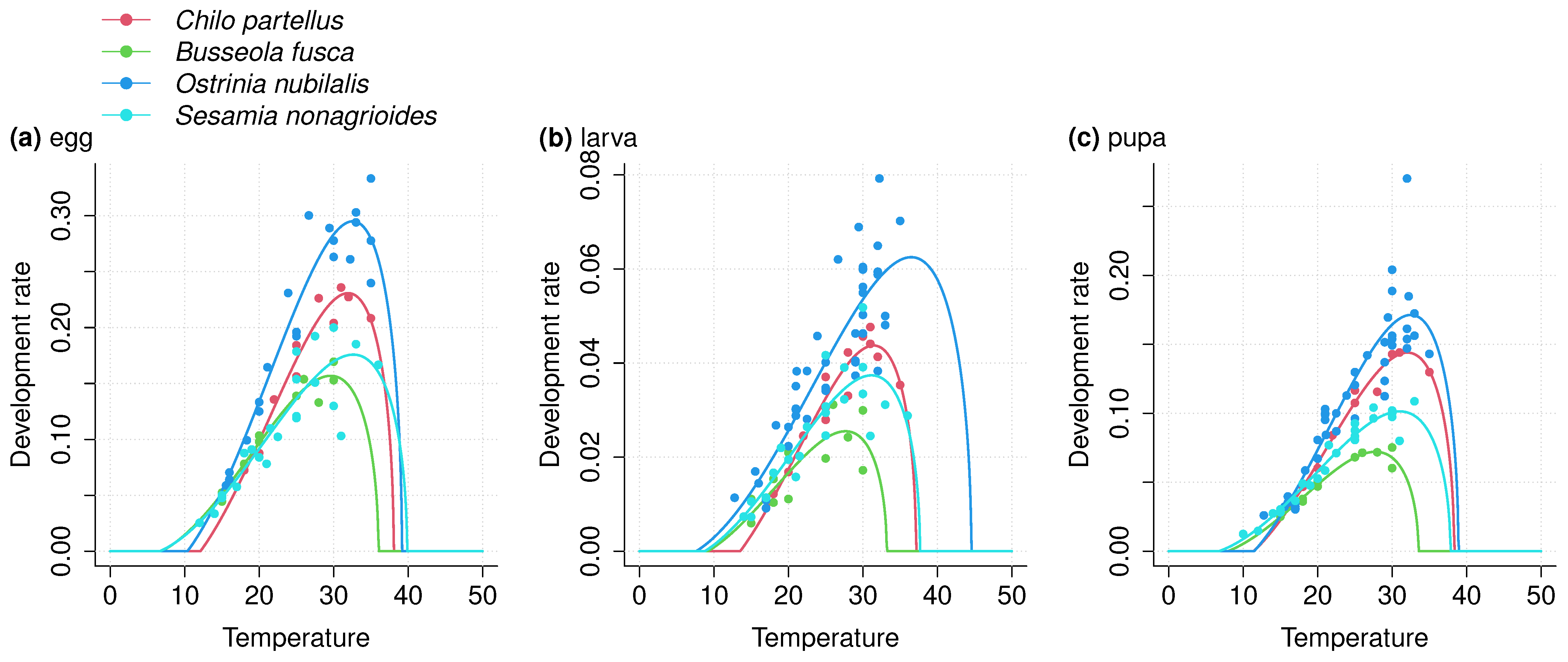
3.1. Thermal Performance Curves
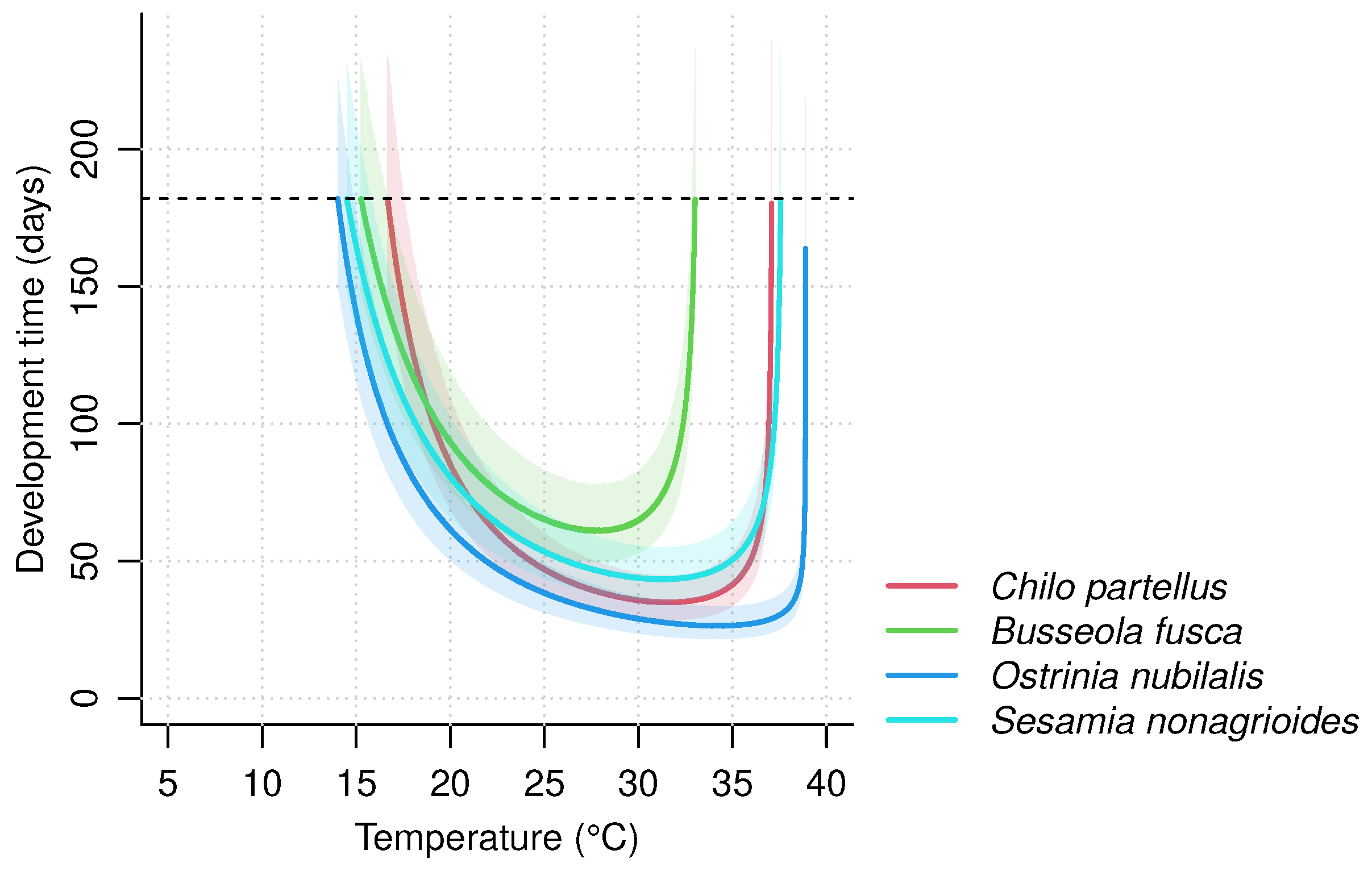
3.2. Impact of Temperature on Development Time
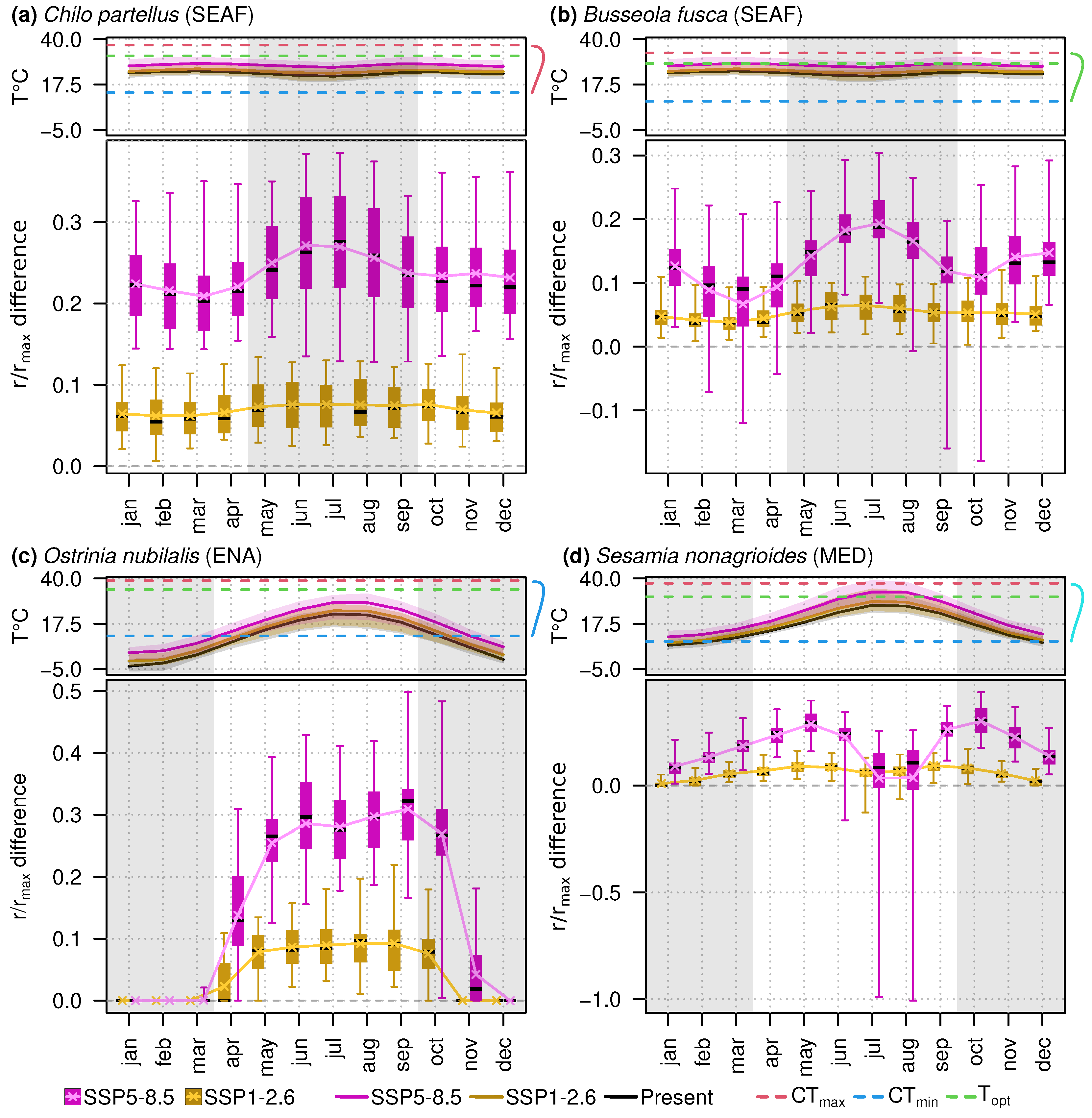
3.3. Impacts of Warming Temperatures
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lister, B.C.; Garcia, A. Climate-driven declines in arthropod abundance restructure a rainforest food web. Proc. Natl. Acad. Sci. USA 2018, 115, E10397–E10406. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Tilman, D. Plant diversity and the stability of foodwebs. Ecol. Lett. 2011, 14, 42–46. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef]
- Kingsolver, J.G. Weather and the population dynamics of insects: Integrating physiological and population ecology. Physiol. Zool. 1989, 62, 314–334. [Google Scholar] [CrossRef]
- Chown, S.; Nicolson, S.W. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, NY, USA, 2004. [Google Scholar]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef]
- Pollard, C.P.; Griffin, C.T.; Andrade Moral, R.d.; Duffy, C.; Chuche, J.; Gaffney, M.T.; Fealy, R.M.; Fealy, R. phenModel: A temperature-dependent phenology/voltinism model for a herbivorous insect incorporating facultative diapause and budburst. Ecol. Model. 2020, 416, 108910. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Chang. Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Eizaguirre, M.; López, C.; Asín, L.; Albajes, R. Thermoperiodism, photoperiodism and sensitive stage in the diapause induction of Sesamia nonagrioides (Lepidoptera: Noctuidae). J. Insect Physiol. 1994, 40, 113–119. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr. Thermal Adaptation; Oxford University Press: Oxford, NY, USA, 2009. [Google Scholar] [CrossRef]
- Huey, R.B.; Stevenson, R. Integrating thermal physiology and ecology of ectotherms: A discussion of approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef]
- Shi, P.J.; Fan, M.L.; Reddy, G.V.P. Comparison of thermal performance equations in describing temperature-dependent developmental rates of insects: (III) Phenological applications. Ann. Entomol. Soc. Am. 2017, 110, 558–564. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431. [Google Scholar] [CrossRef]
- Harcourt, D.G.; Yee, J.M. Polynomial algorithm for predicting the duration of insect life stages. Environ. Entomol. 1982, 11, 581–584. [Google Scholar] [CrossRef]
- Hilbert, D.W.; Logan, J.A. Empirical model of nymphal development for the migratory grasshopper, Melanoplus sanguinipes (Orthoptera: Acrididae). Environ. Entomol. 1983, 12, 1–5. [Google Scholar] [CrossRef]
- Damos, P.; Savopoulou-Soultani, M. Temperature-driven models for insect development and vital thermal requirements. Psyche 2012, 2012, 123405. [Google Scholar] [CrossRef]
- Shi, P.J.; Reddy, G.V.P.; Chen, L.; Ge, F. Comparison of thermal performance equations in describing temperature-dependent developmental rates of insects: (I) Empirical models. Ann. Entomol. Soc. Am. 2015, 109, 211–215. [Google Scholar] [CrossRef]
- Shi, P.J.; Reddy, G.V.P.; Chen, L.; Ge, F. Comparison of thermal performance equations in describing temperature-dependent developmental rates of insects: (II) Two Thermodynamic models. Ann. Entomol. Soc. Am. 2016, 110, 113–120. [Google Scholar] [CrossRef]
- Rebaudo, F.; Rabhi, V.B. Modeling temperature-dependent development rate and phenology in insects: Review of major developments, challenges, and future directions. Entomol. Exp. Appl. 2018, 166, 607–617. [Google Scholar] [CrossRef]
- Glatz, J.; du Plessis, H.; Van den Berg, J. The effect of temperature on the development and reproduction of Busseola fusca (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2017, 107, 39–48. [Google Scholar] [CrossRef]
- Gagnon, A.E.; Bourgeois, G.; Bourdages, L.; Grenier, P.; Blondlot, A. Impact of climate change on Ostrinia nubilalis (Lepidoptera: Crambidae) phenology and its implications on pest management: Impact of climate change on O. nubilalis phenology. Agric. For. Entomol. 2019, 21, 253–264. [Google Scholar] [CrossRef]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of chikungunya and dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kreppel, K.S.; Telfer, S.; Rajerison, M.; Morse, A.; Baylis, M. Effect of temperature and relative humidity on the development times and survival of Synopsyllus fonquerniei and Xenopsylla cheopis, the flea vectors of plague in Madagascar. Parasites Vectors 2016, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vega, D.; Díaz-Aranda, L.M.; Baz, A.; Cifrián, B. Effect of temperature on the survival and development of three forensically relevant Dermestes species (Coleoptera: Dermestidae). J. Med. Entomol. 2017, 54, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef]
- Gilioli, G.; Sperandio, G.; Colturato, M.; Pasquali, S.; Gervasio, P.; Wilstermann, A.; Dominic, A.R.; Schrader, G. Non-linear physiological responses to climate change: The case of Ceratitis capitata distribution and abundance in Europe. Biol. Invasions 2022, 24, 261–279. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Marshall, K.E.; Sewell, M.A.; Levesque, D.L.; Willett, C.S.; Slotsbo, S.; Dong, Y.; Harley, C.D.G.; Marshall, D.J.; Helmuth, B.S.; et al. Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures? Ecol. Lett. 2016, 19, 1372–1385. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef] [PubMed]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Ghalambor, C.K. Are mountain passes higher in the tropics? janzen’s hypothesis revisited. Integr. Comp. Biol. 2006, 46, 5–17. [Google Scholar] [CrossRef]
- Johansson, F.; Orizaola, G.; Nilsson-Örtman, V. Temperate insects with narrow seasonal activity periods can be as vulnerable to climate change as tropical insect species. Sci. Rep. 2020, 10, 8822. [Google Scholar] [CrossRef]
- Martin, T.L.; Huey, R.B. Why “suboptimal” is optimal: Jensen’s inequality and ectotherm thermal preferences. Am. Nat. 2008, 171, E102–E118. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Mailafiya, D.M.; Le Ru, B.P.; Kairu, E.W.; Calatayud, P.A.; Dupas, S. Species diversity of lepidopteran stem borer parasitoids in cultivated and natural habitats in Kenya. J. Appl. Entomol. 2009, 133, 416–429. [Google Scholar] [CrossRef]
- De Groote, H. Maize Yield Losses from stemborers in Kenya. Int. J. Trop. Insect Sci. 2002, 22, 89–96. [Google Scholar] [CrossRef]
- Meissle, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.P.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z.; et al. Pests, pesticide use and alternative options in European maize production: Current status and future prospects: Pest management in European maize production. J. Appl. Entomol. 2009, 134, 357–375. [Google Scholar] [CrossRef]
- Calvin, D.D.; Higgins, R.A.; Knapp, M.C.; Poston, F.L.; Welch, S.M.; Showers, W.B.; Witkowski, J.F.; Mason, C.E.; Chiang, H.C.; Keaster, A.J. Similarities in developmental rates of geographically separate European corn borer (Lepidoptera: Pyralidae) populations. Environ. Entomol. 1991, 20, 441–449. [Google Scholar] [CrossRef]
- Moyal, P.; Tokro, P.; Bayram, A.; Savopoulou-Soultani, M.; Conti, E.; Eizaguirre, M.; Le Rü, B.; Avand-Faghih, A.; Frérot, B.; Andreadis, S. Origin and taxonomic status of the Palearctic population of the stem borer Sesamia nonagrioides (Lefèbvre) (Lepidoptera: Noctuidae). Biol. J. Linn. Soc. 2011, 103, 904–922. [Google Scholar] [CrossRef]
- Kfir, R.; Overholt, W.A.; Khan, Z.R.; Polaszek, A. Biology and management of economically important lepidopteran cereal stem borers in Africa. Annu. Rev. Entomol. 2002, 47, 701–731. [Google Scholar] [CrossRef] [PubMed]
- CABI. Invasive Species Compendium; CAB International: Wallingford, UK, 2022. [Google Scholar]
- Udayagiri, S.; Mason, C.E. Host plant constituents as oviposition stimulants for a generalist herbivore: European corn borer. Entomol. Exp. Appl. 1995, 76, 59–65. [Google Scholar] [CrossRef]
- Willson, H.R. European corn borer, Ostrinia nubilalis (Lepidoptera: Pyralidae), on wheat. Can. Entomol. 1980, 112, 861–863. [Google Scholar] [CrossRef]
- Savinelli, C.E.; Bacheler, J.S.; Bradley, J.R. Ovipositional preferences of the European Corn Borer (Lepidoptera: Pyralidae) for field corn and cotton under field cage conditions in North Carolina. Environ. Entomol. 1988, 17, 688–690. [Google Scholar] [CrossRef]
- Le Rü, B.P.; Ong’amo, G.O.; Moyal, P.; Muchugu, E.; Ngala, L.; Musyoka, B.; Abdullah, Z.; Matama-Kauma, T.; Lada, V.Y.; Pallangyo, B.; et al. Geographic distribution and host plant ranges of East African noctuid stem borers. Ann. Soc. Entomol. Fr. 2006, 42, 353–361. [Google Scholar] [CrossRef]
- Showers, W.B. Geographic variation of the diapause response in the European Corn Borer. In Insect Life History Patterns: Habitat and Geographic Variation; Proceedings in Life Sciences; Springer: New York, NY, USA, 1981. [Google Scholar]
- Moyal, P. Why Sesamia nonagrioides (Lefèbvre) (Lepidoptera: Noctuidae) Is a Pest Only in Portions of Its Geographic Range: The Influence of Climate; Technical Report; IRD: Bondy, France, 2014. [Google Scholar]
- Tams, W. New species of African Heterocera. Entomologist 1932, 65, 1241–1249. [Google Scholar]
- Kfir, R. Competitive displacement of Busseola fusca (Lepidoptera: Noctuidae) by Chilo partellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1997, 90, 619–624. [Google Scholar] [CrossRef]
- Ong’amo, G.O.; Le Rü, B.P.; Dupas, S.; Moyal, P.; Calatayud, P.A.; Silvain, J.F. Distribution, pest status and agro-climatic preferences of lepidopteran stem borers of maize in Kenya. Ann. Soc. Entomol. Fr. 2006, 42, 171–177. [Google Scholar] [CrossRef]
- Ofomata, V.C.; Overholt, W.A.; Egwuatu, R.I. Diapause termination of Chilo partellus (Swinhoe) and Chilo orichalcociliellu Strand (Lepidoptera: Pyralidae). Int. J. Trop. Insect Sci. 1999, 19, 187–191. [Google Scholar] [CrossRef]
- Okuda, T. Factors inducing and terminating larval diapause in a stem borer, Busseola fusca in western Kenya. Jpn. Agric. Res. Q. 1991, 25, 47–54. [Google Scholar]
- Andreadis, S.S.; Kagkelaris, N.K.; Eliopoulos, P.A.; Savopoulou-Soultani, M. Temperature-dependent development of Sesamia nonagrioides. J. Pest Sci. 2013, 86, 409–417. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.; Chatzoglou, C.S. Development of immature stages of Sesamia nonagrioides (Lepidoptera: Noctuidae) under alternating and constant temperatures. Environ. Entomol. 2003, 32, 1337–1342. [Google Scholar] [CrossRef]
- López, C.; Sans, A.; Asin, L.; EizaGuirre, M. Phenological model for Sesamia nonagrioides (Lepidoptera: Noctuidae). Environ. Entomol. 2001, 30, 23–30. [Google Scholar] [CrossRef]
- Hilal, A. Etude du développement de Sesamia nonagrioides et établissement de modèles pour la prévision de ses populations dans la nature. EPPO Bull. 1981, 11, 107–112. [Google Scholar] [CrossRef]
- Matteson, J.W.; Decker, G.C. Development of the European Corn Borer at controlled constant and variable temperatures. J. Econ. Entomol. 1965, 58, 344–349. [Google Scholar] [CrossRef]
- Khadioli, N.; Tonnang, Z.; Muchugu, E.; Ong’amo, G.; Achia, T.; Kipchirchir, I.; Kroschel, J.; Le Ru, B. Effect of temperature on the phenology of Chilo partellus (Swinhoe) (Lepidoptera, Crambidae); simulation and visualization of the potential future distribution of C. partellus in Africa under warmer temperatures through the development of life-table parameters. Bull. Entomol. Res. 2014, 104, 809–822. [Google Scholar] [CrossRef]
- Mbapila, J.C.; Overholt, W.A.; Kayumbo, H.Y. Comparative development and population growth of an exotic stemborer, Chilo partellus (Swinhoe), and an ecologically similar congener, C. orichalcociliellus (Strand) (Lepidoptera: Crambidae). Int. J. Trop. Insect Sci. 2002, 22, 21–27. [Google Scholar] [CrossRef]
- Khadioli, N.; Tonnang, Z.; Ong’amo, G.; Achia, T.; Kipchirchir, I.; Kroschel, J.; Le Ru, B. Effect of temperature on the life history parameters of noctuid lepidopteran stem borers, Busseola fusca and Sesamia calamistis. Ann. Appl. Biol. 2014, 165, 373–386. [Google Scholar] [CrossRef]
- Analytis, S. Über die relation zwischen biologischer entwicklung und temperatur bei phytopathogenen pilzen. J. Phytopathol. 1977, 90, 64–76. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Lowry, R.K.; McMeekin, T.A.; Stokes, A.N.; Chandler, R.E. Model for bacterial culture growth rate throughout the entire biokinetic temperature range. J. Bacteriol. 1983, 154, 1222. [Google Scholar] [CrossRef] [PubMed]
- Yin, X. A nonlinear model for crop development as a function of temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef]
- Shi, P.J.; Chen, L.; Hui, C.; Grissino-Mayer, H.D. Capture the time when plants reach their maximum body size by using the beta sigmoid growth equation. Ecol. Model. 2016, 320, 177–181. [Google Scholar] [CrossRef]
- Briere, J.F.; Pracros, P.; Le Roux, A.Y.; Pierre, J.S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Kontodimas, D.C.; Eliopoulos, P.A.; Stathas, G.J.; Economou, L.P. Comparative temperature-dependent development of Nephus includens (Kirsch) and Nephus bisignatus (Boheman) (Coleoptera: Coccinellidae)preying on Planococcus citri (Risso) (Homoptera: Pseudococcidae): Evaluation of a linear and various nonlinear models using specific criteria. Environ. Entomol. 2004, 33, 1–11. [Google Scholar] [CrossRef]
- Shi, P.; Ge, F.; Sun, Y.; Chen, C. A simple model for describing the effect of temperature on insect developmental rate. J. Asia-Pac. Entomol. 2011, 14, 15–20. [Google Scholar] [CrossRef]
- Régnière, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef]
- Levenberg, K. A method for the solution of certain non-linear problems in least squares. Q. Appl. Math. 1944, 2, 164–168. [Google Scholar] [CrossRef]
- Marquardt, D.W. An algorithm for least-squares estimation of nonlinear parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Régnier, B.; Legrand, J.; Rebaudo, F. Modeling temperature-dependent development rate in insects and implications of experimental design. Environ. Entomol. 2022, 51, 132–144. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R.; Burnham, K.P. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Sinclair, B.J.; Williams, C.M.; Terblanche, J.S. Variation in thermal performance among insect populations. Physiol. Biochem. Zool. 2012, 85, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Jones, R.; Narisma, G.; Alves, L.; Amjad, M.; Gorodetskaya, I.; Grose, M.; Klutse, N.; Krakovska, S.; Li, J.; et al. 2021: Atlas. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; Available online: http://interactive-atlas.ipcc.ch/ (accessed on 11 October 2022).
- Iturbide, M.; Fernández, J.; Gutiérrez, J.M.; Bedia, J.; Cimadevilla, E.; Díez-Sierra, J.; Manzanas, R.; Casanueva, A.; Baño-Medina, J.; Milovac, J.; et al. Repository supporting the implementation of FAIR principles in the IPCC-WGI Atlas. Zenodo 2021. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Vol. In Press; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar] [CrossRef]
- Iturbide, M.; Gutiérrez, J.M.; Alves, L.M.; Bedia, J.; Cerezo-Mota, R.; Cimadevilla, E.; Cofiño, A.S.; Di Luca, A.; Faria, S.H.; Gorodetskaya, I.V.; et al. An update of IPCC climate reference regions for subcontinental analysis of climate model data: Definition and aggregated datasets. Earth Syst. Sci. Data 2020, 12, 2959–2970. [Google Scholar] [CrossRef]
- Vasseur, D.A.; DeLong, J.P.; Gilbert, B.; Greig, H.S.; Harley, C.D.G.; McCann, K.S.; Savage, V.; Tunney, T.D.; O’Connor, M.I. Increased temperature variation poses a greater risk to species than climate warming. Proc. R. Soc. Biol. Sci. 2014, 281, 20132612. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blanford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Rebaudo, F.; Struelens, Q.; Dangles, O. Modelling temperature-dependent development rate and phenology in arthropods: The devRate package for R. Methods Ecol. Evol. 2018, 9, 1144–1150. [Google Scholar] [CrossRef]
- Landau, W. The targets R package: A dynamic Make-like function-oriented pipeline toolkit for reproducibility and high-performance computing. J. Open Source Softw. 2021, 6, 2959. [Google Scholar] [CrossRef]
- Sharpe, P.J.; Curry, G.L.; DeMichele, D.W.; Cole, C.L. Distribution model of organism development times. J. Theor. Biol. 1977, 66, 21–38. [Google Scholar] [CrossRef]
- Gilbert, E.; Powell, J.; Logan, J.; Bentz, B. Comparison of three models predicting developmental milestones given environmental and individual variation. Bull. Math. Biol. 2004, 66, 1821–1850. [Google Scholar] [CrossRef]
- Yurk, B.P.; Powell, J.A. Modeling the effects of developmental variation on insect phenology. Bull. Math. Biol. 2010, 72, 1334–1360. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Ratkowsky, D.A.; Wang, N.; Li, Y.; Zhao, L.; Reddy, G.V.; Li, B.L. Comparison of five methods for parameter estimation under Taylor’s power law. Ecol. Complex. 2017, 32, 121–130. [Google Scholar] [CrossRef]
- Schmalensee, L.; Hulda Gunnarsdóttir, K.; Näslund, J.; Gotthard, K.; Lehmann, P. Thermal performance under constant temperatures can accurately predict insect development times across naturally variable microclimates. Ecol. Lett. 2021, 24, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Tatebe, H.; Ogura, T.; Nitta, T.; Komuro, Y.; Ogochi, K.; Takemura, T.; Sudo, K.; Sekiguchi, M.; Abe, M.; Saito, F.; et al. Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model Dev. 2019, 12, 2727–2765. [Google Scholar] [CrossRef]
- Got, B.; Piry, S.; Migeon, A.; Labatte, J.M. Comparison of different models for predicting development time of the European Corn Borer (Lepidoptera: Pyralidae). Environ. Entomol. 1997, 26, 46–60. [Google Scholar] [CrossRef]
- Logan, J.A.; Wollkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An analytic model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Mwalusepo, S.; Tonnang, H.E.Z.; Massawe, E.S.; Okuku, G.O.; Khadioli, N.; Johansson, T.; Calatayud, P.A.; Le Ru, B.P. Predicting the impact of temperature change on the future distribution of maize stem borers and their natural enemies along East African mountain gradients using phenology models. PLoS ONE 2015, 10, e0130427. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation of climate extremes with global warming in the Mediterranean region and its north versus south contrast. Reg. Environ. Chang. 2020, 20, 31. [Google Scholar] [CrossRef]
- Maiorano, A.; Donatelli, M.; Fumagalli, D. Potential distribution and phenological development of the Mediterranean Corn Borer (Sesamia nonagrioides) under warming climate in Europe. Int. Congr. Environ. Model. Softw. 2012, 324. [Google Scholar]
- Boé, J.; Somot, S.; Corre, L.; Nabat, P. Large discrepancies in summer climate change over Europe as projected by global and regional climate models: Causes and consequences. Clim. Dyn. 2020, 54, 2981–3002. [Google Scholar] [CrossRef]
- Pincebourde, S.; Woods, H.A. Climate uncertainty on leaf surfaces: The biophysics of leaf microclimates and their consequences for leaf-dwelling organisms. Funct. Ecol. 2012, 26, 844–853. [Google Scholar] [CrossRef]
- Pincebourde, S.; Woods, H.A. There is plenty of room at the bottom: Microclimates drive insect vulnerability to climate change. Curr. Opin. Insect Sci. 2020, 41, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Southworth, J.; Randolph, J.; Habeck, M.; Doering, O.; Pfeifer, R.; Rao, D.; Johnston, J. Consequences of future climate change and changing climate variability on maize yields in the midwestern United States. Agric. Ecosyst. Environ. 2000, 82, 139–158. [Google Scholar] [CrossRef]
- Jones, P.G.; Thornton, P.K. The potential impacts of climate change on maize production in Africa and Latin America in 2055. Glob. Environ. Chang. 2003, 13, 51–59. [Google Scholar] [CrossRef]
- Li, X.; Takahashi, T.; Suzuki, N.; Kaiser, H.M. The impact of climate change on maize yields in the United States and China. Agric. Syst. 2011, 104, 348–353. [Google Scholar] [CrossRef]
- Marshall, K.E.; Gotthard, K.; Williams, C.M. Evolutionary impacts of winter climate change on insects. Curr. Opin. Insect Sci. 2020, 41, 54–62. [Google Scholar] [CrossRef]
- Wall, C. Effect of temperature on embryonic development and diapause in Chesias legatella (Lepidoptera: Geometridae). J. Zool. 1974, 172, 147–168. [Google Scholar] [CrossRef]
- Kimberling, D.N.; Miller, J.C. Effects of temperature on larval eclosion of the winter moth, Operophtera brumata. Entomol. Exp. Appl. 1988, 47, 249–254. [Google Scholar] [CrossRef]
- Buckley, L.B.; Huey, R.B. How Extreme Temperatures Impact Organisms and the Evolution of their Thermal Tolerance. Integr. Comp. Biol. 2016, 56, 98–109. [Google Scholar] [CrossRef]
- Urban, M.C.; Richardson, J.L.; Freidenfelds, N.A. Plasticity and genetic adaptation mediate amphibian and reptile responses to climate change. Evol. Appl. 2014, 7, 88–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Region | Temperatures (°C) | Life Stages 1 | Reference |
|---|---|---|---|---|
| S. nonagrioides | Greece | 14, 17, 21, 25, 31 | e, l, p | [57] |
| S. nonagrioides | Greece | 20, 22.5, 25, 27.5, 30 | e, l, p | [58] |
| S. nonagrioides | Spain | 12, 15, 18, 21.5, 25, 27.5, 30, 33, 36 | e, l, p | [59] |
| S. nonagrioides | Morocco | 15, 19, 25, 30 | e, l, p | [60] |
| O. nubilalis | Iowa | 17, 21, 25, 29, 30, 32 | l, p | [42] |
| O. nubilalis | North Dakota | 17, 21, 29, 30, 32 | l, p | [42] |
| O. nubilalis | Delaware | 17, 21, 29, 30, 32 | l, p | [42] |
| O. nubilalis | Missouri | 17, 21, 29, 30, 32 | l, p | [42] |
| O. nubilalis | Illinois | 15.6, 18.3, 21.1, 23.9, 26.7, 29.4, 32.2, 35 | e, l, p | [61] |
| O. nubilalis 2 | Quebec | 16, 20, 22.5, 25, 30, 33, 35 | e, l, p | [25] |
| C. partellus | Kenya | 18, 20, 25, 30, 32, 35 | e, l, p | [62] |
| C. partellus | Kenya | 22, 25, 28, 31 | e, l, p | [63] |
| B. fusca | Kenya | 15, 18, 20, 25, 28, 30 | e, l, p | [64] |
| B. fusca | South Africa | 15, 18, 20, 26, 30 | e, l, p | [24] |
| Name | Equation | Reference |
|---|---|---|
| analytis_77 | [65] | |
| ratkowsky_83 | [66] | |
| hilbertLogan_83 | [19] | |
| beta_95 | [67] | |
| beta_16 | [68] | |
| briere1_99 | [69] | |
| briere2_99 | [69] | |
| kontodimas_04 | [70] | |
| shi_11 | [71] | |
| perf2_11 | [71] | |
| regniere_12 | [72] |
| Species | Stage | Model | Parameter | Estimate | Standard Error |
|---|---|---|---|---|---|
| Chilo partellus | egg | kontodimas_04 | a | ||
| larva | briere1_99 | a | |||
| pupa | briere1_99 | a | |||
| Busseola fusca | egg | kontodimas_04 | a | ||
| larva | briere1_99 | a | |||
| pupa | briere1_99 | a | |||
| Ostrinia nubilalis | egg | kontodimas_04 | a | ||
| larva | briere1_99 | a | |||
| pupa | perf2_11 | c | |||
| k | |||||
| Sesamia nonagrioides | egg | kontodimas_04 | a | ||
| larva | kontodimas_04 | a | |||
| pupa | briere1_99 | a | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Régnier, B.; Legrand, J.; Calatayud, P.-A.; Rebaudo, F. Developmental Differentiations of Major Maize Stemborers Due to Global Warming in Temperate and Tropical Climates. Insects 2023, 14, 51. https://doi.org/10.3390/insects14010051
Régnier B, Legrand J, Calatayud P-A, Rebaudo F. Developmental Differentiations of Major Maize Stemborers Due to Global Warming in Temperate and Tropical Climates. Insects. 2023; 14(1):51. https://doi.org/10.3390/insects14010051
Chicago/Turabian StyleRégnier, Baptiste, Judith Legrand, Paul-André Calatayud, and François Rebaudo. 2023. "Developmental Differentiations of Major Maize Stemborers Due to Global Warming in Temperate and Tropical Climates" Insects 14, no. 1: 51. https://doi.org/10.3390/insects14010051
APA StyleRégnier, B., Legrand, J., Calatayud, P.-A., & Rebaudo, F. (2023). Developmental Differentiations of Major Maize Stemborers Due to Global Warming in Temperate and Tropical Climates. Insects, 14(1), 51. https://doi.org/10.3390/insects14010051