
Transcriptome and Metabolome Analysis Reveals the Importance of Amino-Acid Metabolism in Spodoptera Frugiperda Exposed to Spinetoram



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Test Reagent
2.3. Acute Toxicity Assay
2.4. Sample Preparation
2.5. Transcriptome Analysis
2.6. Metabolite Extraction
2.7. LC-MS/MS Analysis
2.8. Processing and Statistical Analysis of Metabolomics Data
3. Results
3.1. Toxicity of Spinetoram to S. Frugiperda 3rd Instar Larvae
3.2. Transcriptome Analysis of S. Frugiperda in Responses to Spinetoram Application
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Todd, E.L.; Poole, R.W. Keys and illustrations for the armyworm moths of the Noctuid Genus Spodoptera Guenée from the Western hemisphere. Ann. Entomol. Soc. Am. 1980, 6, 722–738. [Google Scholar] [CrossRef]
- Jing, D.P.; Guo, F.F.; Jiang, Y.Y.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2020, 27, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Fleischer, S.; Meagher, R.L.; Hay-Roe, M.; Khan, A.; Gabriela, M.M.; Silvie, P.; Vergara, C.; Westbrook, J. Fall armyworm migration across the Lesser Antilles and the potential for genetic exchanges between North and South American populations. PLoS ONE 2017, 12, e0175076. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Liu, J.; Li, T.J.; Jiang, Y.Y.; Zeng, J.; Wang, Y.P. Occurrence characteristics of main maize diseases and insect pests in China in 2020. China Plant Prot. 2021, 41, 30–35. [Google Scholar]
- Gui, F.R.; Lan, T.M.; Zhao, Y.; Guo, W.; Dong, Y.; Fang, D.; Haimeng, H.L.; Wang, H.L.; Hao, R.S.; Cheng, X. Genomic and transcriptomic analysis unveils population evolution and development of pesticide resistance in fall armyworm Spodoptera Frugiperda. Protein Cell 2020, 13, 513–531. [Google Scholar] [CrossRef]
- Li, Y.; Gong, L.F.; Wang, H.H.; Li, X.; Sun, G.; Gu, S.H.; Liang, P.; Gao, X.W. Genotype and mutation frequency of ace-1, the target gene of organophosphorus and carbamate insecticides, in field populations of Spodoptera frugiperda (Lepidoptera: Noctuidae) in China. Acta Entomol. Sin. 2020, 63, 574–581. [Google Scholar]
- Rumbos, C.I.; Dutton, A.C.; Athanassiou, C.G. Insecticidal effect of spinetoram and thiamethoxam applied alone or in combination for the control of major stored-product beetle species. J. Stored Prod. Res. 2017, 75, 56–63. [Google Scholar] [CrossRef]
- Gao, Z.P.; Chen, Y.; He, K.L.; Guo, J.F.; Wang, Z.Y. Sublethal effects of the microbial-derived insecticide spinetoram on growth and fecundity of the fall armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 2020, 114, 1582–1587. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Ann. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Wang, D.; Wang, M.Y.; Liu, H.Y.; Xin ZXing, M. Lethal and sublethal effects of spinosad on Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol. 2012, 106, 1825–1831. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, M.; Habib, A.; Aziz, S.G.; Ali, R.B. Sublethal effects of indoxacarb on the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Appl.Entomol. Zool. 2019, 46, 75–80. [Google Scholar]
- Shahout, H.A.; Xu, J.X.; Qiao, J.; Jia, Q.D. Sublethal effects of methoxyfenozide on growth and development of common cutworm Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Res. J. Biol. Sci. 2012, 6, 58–64. [Google Scholar] [CrossRef][Green Version]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoidsthiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honeybees (Apis mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar]
- Fogel, M.N.; Schneider, M.I.; Desneux, N.; Gonz’alez, B.; Ronco, A.E. Impact of the neonicotinoid acetamiprid on immature stages of the predator Eriopis connexa (Coleoptera: Coccinellidae). Ecotoxicology 2013, 22, 1063–1071. [Google Scholar] [CrossRef]
- Delkash-Roudsari, S.; Chicas-Mosierc, A.M.; Goldansaz, S.H.; Talebi-Jahromi, K.; Ashouri, A.; Abramson, C.I. Assessment of lethal and sublethal effects of imidacloprid, ethion, and glyphosate on aversive conditioning, motility, and lifespan in honey bees (Apis mellifera L.). Ecotoxicol. Environ. Saf. 2022, 204, 111108. [Google Scholar] [CrossRef]
- Biondi, A.; Zappala, L.; Stark, J.D.; Desneux, N. Do biopesticides affect the demographic traits of a parasitoid wasp andits biocontrol services through sublethal effects? PLoS ONE 2013, 8, e76548. [Google Scholar] [CrossRef]
- Schmehl, D.R.; Teal, P.E.A.; Frazier, J.L.; Grozinger, C.M. Genomic analysis of the interaction between pesticide exposure and nutrition in honeybees (Apis melli fera). J. Insect Physiol. 2014, 71, 177–190. [Google Scholar] [CrossRef]
- Marco, L.; De, S.D.; Epis, S.; Mastrantonio, V.; Ferrari, M.; Ricci, I.; Comandatore, F.; Bandi CPorretta, D.; Urbanelli, S. The choreography of the chemical defensome response to insecticide stress: Insights into the Anopheles stephensi transcriptome using RNA-Seq. Sci. Rep. 2017, 7, 41312. [Google Scholar] [CrossRef]
- Wang, R.L.; Zhu-Salzman, K.; Baersonm, S.R.; Xin, X.W.; Li, J.; Su, Y.J.; Zeng, R.S. Identification of a novel cytochrome P450 CYP321B1 gene from tobacco cutworm (Spodoptera litura) and RNA interference to evaluate its role in commonly used insecticides. Insect Sci. 2017, 24, 235–247. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mole. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Pavlidi, N.; Vontas, J.; Leeuwen, T.V. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Fang, S.M.; Huang, K.; Yu, Q.Y.; Zhang, Z. Characterization of an epsilon-class glutathione S-transferase involved in tolerance in the silkworm larvae after long term exposure to insecticides. Ecotoxicol. Environ. Saf. 2015, 120, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Qi, X.; Zhou, Y. Metabolomics and the Research of Traditional Chinese Medicine. Adv. Mater. Res. 2011, 396–398, 1676–1679. [Google Scholar] [CrossRef]
- Bordag, N.; Janakiraman, V.; Nachtigall, J.; Maldonado, S.G.; Bethan, B.; Laine, J.; Fux, E. Fast filtration of bacterial or mammalian suspension cell cultures for optimal metabolomics results. PLoS ONE 2016, 11, e0159389. [Google Scholar] [CrossRef]
- Lv, N.N.; Ma, K.S.; Li, R.; Liang, P.Z.; Liang, P.; Gao, X.W. Sublethal and lethal effects of the imidacloprid on the metabolic characteristics based on high-throughput non-targeted metabolomics in Aphis gossypii Glover. Ecotoxicol. Environ. Saf. 2021, 212, 111969. [Google Scholar] [CrossRef]
- Shi, T.; Burton, S.; Wang, Y.; Xu, S.; Zhang, W.; Yu, L. Metabolomic analysis of honeybee, Apis mellifera L. response to thiacloprid. Pestic. Biochem. Phys. 2018, 152, 17–23. [Google Scholar] [CrossRef]
- Feng, M.; Fei, S.G.; Xia, J.M.; Zhang, M.M.; Wu, H.Y.; Swevers, L.; Sun, J.C. Global metabolic profiling of baculovirus infection in silkworm hemolymph shows the importance of amino-acid metabolism. Viruses 2021, 13, 841. [Google Scholar] [CrossRef]
- Gao, Z.P.; Guo, J.F.; He, K.L.; Wang, Z.Y. Toxicity of spinetoram and its effects on the detoxifying enzyme and acetyl cholinesterase activities in Spodoptera frugiperda (Lepidoptera:Noctuidae) larvae. Acta Entomol. Sin. 2020, 63, 558–564. [Google Scholar]
- Nicholson, J.K. Global systems biology, personalized medicine and molecular epidemiology. Mol. Syst. Biol. 2006, 2, 52. [Google Scholar] [CrossRef]
- Clemente Plaza, N.; Reig García-Galbis, M.; Martínez-Espinosa, R.M. Effects of the usage of L-cysteine (L-cys) on human health. Molecules 2018, 23, 575. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.R.; Tompkins, S.C.; Taylor, E.B. Regulation of pyruvate metabolism and human disease. Cell. Mol. Life Sci. 2014, 71, 2577–2604. [Google Scholar] [CrossRef] [PubMed]
- Fotiadis, D.; Kanai, Y.; Palacín, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Aspects Med. 2013, 34, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Neckameyer, W.; Coleman, C.; Eadie, S.; Goodwin, S. Compartmentalization of neuronal and peripheral serotonin synthesis in Drosophila melanogaster. Genes Brain Behav. 2007, 6, 756–769. [Google Scholar] [CrossRef]
- Kalb, D.; Heinekamp, T.; Schieferdecker, S.; Nett, M.; Brakhage, A.A.; Hoffmeister, D. An iterative O-methyltransferase catalyzes 1, 11-dimethylation of Aspergillus fumigatus fumaric acid amides. ChemBioChem 2016, 17, 1813–1817. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Shu, Q.; Chen, J.; Fan, K.; Xu, P.; Zhou, Q.; Li, C.; Zheng, H. Antibiotic exposure has sex-dependent effects on the gut microbiota and metabolism of short-chain fatty acids and amino acids in mice. mSystems 2019, 4, e00048-19. [Google Scholar] [CrossRef]
- White, P.J.; Newgard, C.B. Branched-chain amino acids in disease. Science 2019, 363, 582.e583. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968. [Google Scholar] [CrossRef]
- Matsumoto, S.; Yamamoto, S.; Sai, K.; Maruo, K.; Adachi MSaitoh, M.; Nishizaki, T. Pipecolic acid induces apoptosis in neuronal cells. Brain Res. 2003, 980, 179.e184. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, G.; Wang-Pruski, G.; You, M. Phyllotreta striolata (Coleoptera: Chrysomelidae): Arginine kinase cloning and RNAi-based pest control. Eur. J. Entomo. 2008, 105, 815–822. [Google Scholar] [CrossRef]
- Gutierrez, M.C.; Delgado-Coello, B.A. Influence of pipecolic acid on thus, may cause hepatic encephalopathy by inducing 3 the release and uptake of [3H] GABA from brain slices of mouse cerebral cortex. Neurochem. Res. 1989, 14, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Hori, T.; Nakajima, M.; Takeshita, K. Plasma levels of pipecolic acid in patients with chronic liver disease. Hepatology 1988, 8, 286–289. [Google Scholar] [CrossRef]
- Takahama, K.; Hashimoto, T.; Wang, M.W.; Akaike, N.; Hitoshi, T.; Okano, Y.; Kasé, Y.; Miyata, T. Pipecolic acid enhancement of GABA route in the rat brain. Neuropharmacology 1986, 25, 339–342. [Google Scholar] [CrossRef]
- Takahama, K.; Miyata, T.; Hashimoto, T.; Okano, Y.; Hitoshi, T.; Kase, Y. Pipecolic acid: A new type of α-amino acid possessing bicuculline-sensitive action in the mammalian brain. Brain Res. 1982, 239, 294–298. [Google Scholar] [CrossRef]
- Yang, L.F.; Venneti, S.; Nagrath, D. Glutaminolysis: A Hallmark of Cancer Metabolism. Ann Rev. Biomed. Eng. 2017, 19, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Poupin, N.; Tremblay-Franco, M.; Amiel, A.; Canlet, C.; Remond, D.; Debrauwer, L.; Dardevet, D.; Thiele, I.; Aurich, M.K.; Jourdan, F.; et al. Arterio-venous metabolomics exploration reveals major changes across liver and intestine in the obese Yucatan minipig. Sci. Rep. 2019, 9, 12527. [Google Scholar] [CrossRef]
- Clelend, T.A. Inhibitory Glutamate Receptor Channels. Mol. Neurobiol. 1996, 13, 97–136. [Google Scholar] [CrossRef]
- Sceniak, M.P.; Spitsbergen, J.B.; Sabo, S.L.; Yuan, Y.; Atchison, W.D. Acute neurotoxicant exposure induces hyperexcitability in mouse lumbar spinal motor neurons. J. Neurophysiol. 2020, 123, 1448–1459. [Google Scholar] [CrossRef]
- Lambe, E.K.; Picciotto, M.R.; Aghajanian, G.K. Nicotine induces glutamate release from thalamocortical terminals in prefrontal cortex. Neuropsychopharmacology 2003, 28, 216–225. [Google Scholar] [CrossRef]
- Humphries, P.; Pretorius, E.; Naude, H. Direct and indirect cellular effects of aspartame on the brain. Eur. J. Clin. Nutr. 2008, 62, 451–462. [Google Scholar] [CrossRef]
- Patetsini, E.; Dimitriadis, V.; Kaloyianni, M. Biomarkers in marine mussels, Mytilus galloprovincialis, exposed to environmentally relevant levels of the pesticides, chlorpyrifos and penoxsulam. Aquat. Toxicol. 2013, 126, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Anadón, A.; Wu, Q.; Qiao, F.; Ares, I.; Martínez-Larrañaga, R.; Yuan, Z.H.; Martínez, M. Mechanism of neonicotinoid toxicity: Impact on oxidative stress and metabolism. Annu. Rev. Pharmacol. 2018, 58, 471–507. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Meng, Z.; Tian, S.; Teng, M.; Yan, J.; Jia, M.; Li, R.; Zhou, Z.; Zhu, W. Neonicotinoid insecticides exposure cause amino acid metabolism disorders, lipid accumulation and oxidative stress in ICR mice. Chemosphere 2020, 246, 125661. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G.; Johnsoncadwell, L.; Vesce, S.; Jekabsons, M.B.; Yadava, N. Bioenergetics of mitochondria in cultured neurons and their role in glutamate excitotoxicity. J. Neurosci. Res. 2007, 85, 3206–3212. [Google Scholar] [CrossRef]
- Dikalov, S.; Nazarewicz, R.R.; Panov, A.; Harrison, D.G.; Dikalova, A. Crosstalk between mitochondrial ROS and NADPH oxidases in cardiovascular and degenerative diseases: Application of mitochondria-targeted antioxidants. Free Radical Bio. Med. 2011, 51, S85–S86. [Google Scholar] [CrossRef]
- El-Gendy, K.S.; Aly, N.M.; Mahmoud, F.H.; Kenawy, A.; El-Sebae, A.K.H. The role of vitamin C as antioxidant in protection of oxidative stress induced by imidacloprid. Food Chem. Toxicol. 2010, 48, 215–221. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532.e553. [Google Scholar] [CrossRef]
- John, X. The physiological role of dehydroascorbic acid. FEBS Lett. 2002, 527, 1–3. [Google Scholar]
- Hideg, E.; Jansen, M.A.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar]
- Stevens, R.; Page, D.; Gouble, B.; Garchery, C.; Zamir, D.; Causse, M. Tomato fruit ascorbic acid content is linked with monodehydroascorbate reductase activity and tolerance to chilling stress. Plant Cell Environ. 2008, 31, 1086–1096. [Google Scholar] [CrossRef]
- Kadoma, Y.; Ishihara, M.; Okada, N.; Fujisawa, S. Free radical interaction between vitamin E (alpha-, beta-, gamma- and delta-tocopherol): Ascorbate and flavonoids. In Vivo 2006, 20, 823–827. [Google Scholar]
- Wang, X.; Tan, Z.; Chen, S.; Lin, G.; Li, X.C.; Ke, D.; Hou, L.P.; Leung, J.Y.S. Norethindrone causes cellular and hepatic injury in zebrafish by compromising the metabolic processes associated with antioxidant defence: Insights from metabolomics. Chemosphere 2010, 275, 130049. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Bio. 2012, 13, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Gupte, R.; Liu, Z.; Kraus, W.L. PARPs and ADP-ribosylation: Recent advances linking molecular functions to biological outcomes. Gen. Dev. 2017, 31, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Chang, P. Insights into the biogenesis, function, and regulation of ADP-ribosylation. Nat. Chem. Biol. 2018, 14, 236–243. [Google Scholar] [CrossRef]
- Prutz, W.A.; Kissner, R.; Koppenol, W.H.; Rüegger, H. On the irreversible destruction of reduced nicotinamide nucleotides by hypohalous acids. Arch. Biochem. Biophys. 2000, 380, 181–191. [Google Scholar] [CrossRef]
- Zhu, B.T. Development of selective immune tolerance towards the allogeneic fetus during pregnancy: Role of tryptophan catabolites. J. Mole. Med. 2010, 25, 831–835. [Google Scholar] [CrossRef]
- Gareth, J. Sanger. 5-Hydroxytryptamine and the gastrointestinal tract: Where next? Trends Pharmacol. Sci. 2008, 29, 465–471. [Google Scholar]
- Wang, L.S.; Zhang, M.F.; Tao, X.; Zhou, Y.F.; Liu, X.M.; Pan, R.L.; Liao, Y.H.; Chang, Q. Lc-ms/ms-based quantification of tryptophan metabolites and neurotransmitters in the serum and brain of mice. J. Chromatogr. B 2019, 1112, 24–32. [Google Scholar] [CrossRef]
- Russo, S.; Kema, I.P.; Bosker, F.; Haavik, J.; Korf, J. Tryptophan as an evolutionarily conserved signal to brain serotonin: Molecular evidence and psychiatric implications. World J. Biol. Psychia. 2009, 10, 258–268. [Google Scholar] [CrossRef]
- Liu, S.Y.; Aliyari, R.; Chikere, K.; Li, G.; Marsden, M.D.; Smith, J.K.; Pernet, O.; Guo, H.T.; Nusbaum, R.; Zack, J.A.; et al. Interferoninducible cholesterol-25-hydroxylase broadly inhibits viral entry by production of 25-hydroxycholesterol. Immunity 2013, 38, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Lee, W.J. mmune-metabolic interactions during systemic and enteric infection in Drosophila. Curr. Opin. Insect Sci. 2018, 29, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.A.; Narasimhamurthy, R.K.; Joshi, M.B.; Dsouza, H.S.; Mumbrekar, K.D. Pesticides exposure-induced changes in brain metabolome: Implications in the pathogenesis of neurodegenerative disorders. Neurotoxi. Res. 2022. [CrossRef] [PubMed]
- Besson, M.J.; Cheramy, A.; Feltz, P.; Glowinski, J. Release of newly synthesized dopamine from dopamine-containing terminals in the striatum of the rat. Pro. Nat. Acad. Sci. USA. 1969, 62, 741–748. [Google Scholar] [CrossRef]
- Gotti, C.; Zoli, M.; Clementi, F. Brain nicotinic acetylcholine receptors: Native subtypes and their relevance. Trends Pharmacol. Sci. 2006, 27, 482–491. [Google Scholar] [CrossRef]
- Fhan, S. Description of Parkinson’s disease as a clinical syndrome. Ann. N. Y. Acad. Sci. 2003, 991, 1–14. [Google Scholar] [CrossRef]
- Miller, D.B.; O’Callaghan, J.P. Biomarkers of Parkinson’s disease: Present and future. Metabolism 2015, 64, S40–S46. [Google Scholar] [CrossRef]
- Chen, X.; Zheng, J.; Teng, M.; Zhang, J.; Qian, L.; Duan, M.; Zhao, F.; Zhao, W.; Wang, Z.; Wang, C. Bioaccumulation, metabolism and the toxic effects of chlorfenapyr in zebrafish (Danio rerio). J. Agric. Food Chem. 2021, 69, 8110–8119. [Google Scholar] [CrossRef]
- McCormack, A.L.; Atienza, J.G.; Langston, J.W.; Di Monte, D.A. Decreased susceptibility to oxidative stress underlies the resistance of specific dopaminergic cell populations to paraquat-induced degeneration. Neuroscience 2006, 141, 929–937. [Google Scholar] [CrossRef]
- Arruda, P.C.; de Magalhães, L.; Camilloa, M.A.; Rogero, J.R.; Troncone, L.R. Ionotropic glutamate receptors regulating labeled acetylcholine release from rat striatal tissue in vitro: Possible involvement of receptor modulation in magnesium sensitivity. Neurosc. Res. 2004, 49, 289–295. [Google Scholar]





{kind=link}
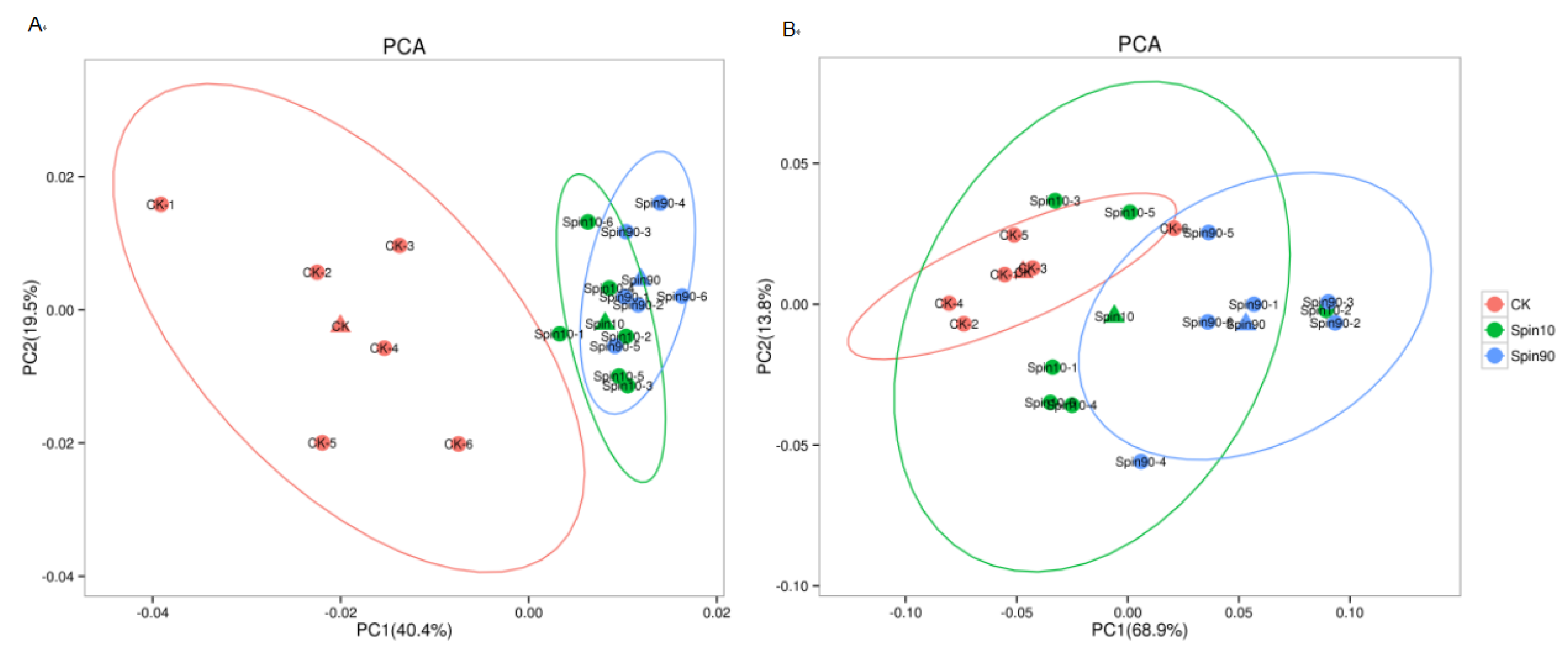{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads | Clean Bases | Mapped Read | Guanine-Cytosine Content (%) | Q20(%) | Q30(%) | |
|---|---|---|---|---|---|---|---|
| CK | Control-1 | 24,804,561 | 7,399,530,118 | 36,072,981 (72.71%) | 48.04 | 98.29 | 94.93 |
| Control-2 | 25,406,000 | 7,581,035,800 | 367,63,311 (72.35%) | 48.00 | 97.86 | 94.93 | |
| Control-3 | 20,529,036 | 6,132,802,190 | 29,967,481 (72.99%) | 47.78 | 97.94 | 93.77 | |
| Control-4 | 21,455,769 | 6,405,367,636 | 31,161,221 (72.62%) | 49.17 | 97.71 | 93.29 | |
| LC10 | Spin10-1 | 24,388,290 | 7,285,860,770 | 34,931,682 (71.62%) | 48.22 | 97.95 | 93.82 |
| Spin10-2 | 23,612,040 | 7,059,990,216 | 33,693,849 (71.35%) | 47.86 | 97.71 | 93.32 | |
| Spin10-3 | 23,612,040 | 6,567,603,744 | 31,630,174 (71.99%) | 47.92 | 97.59 | 93.03 | |
| Spin10-4 | 21,780,281 | 6,504,361,012 | 31,270,395 (71.79%) | 48.20 | 97.78 | 93.44 | |
| LC90 | Spin90-1 | 20,923,896 | 6,261,061,732 | 29,946,614 (71.56%) | 47.73 | 97.63 | 93.03 |
| Spin90-2 | 20,796,109 | 6,220,125,194 | 29,921,237 (71.94%) | 47.40 | 97.64 | 93.10 | |
| Spin90-3 | 25,473,623 | 7,599,203,900 | 36,194,890 (71.04%) | 48.25 | 97.83 | 93.53 | |
| Spin90-4 | 27,731,793 | 8,274,619,114 | 39,174,512 (70.63%) | 48.49 | 97.86 | 93.65 |
| Unigene ID | Gene Annotation | Type of Regulation | |
|---|---|---|---|
| LC10 Group | LC90 Group | ||
| LOC118266960 | 5-hydroxytryptamine receptor | Up | Up |
| LOC118275809 | 5-hydroxytryptamine receptor 1A | - | Up |
| LOC118262451 | acetylcholinesterase | Up | Up |
| LOC118268389 | acetylcholine receptor subunit alpha | Up | Up |
| LOC118275478 | cholinesterase 2 | - | Up |
| LOC118276148 | esterase B1 | - | Up |
| LOC118276231 | juvenile hormone esterase-like | Down | Down |
| LOC118274652 | glucose dehydrogenase | Up | Up |
| LOC118277268 | glucose-1-phosphatase-like | Up | - |
| LOC118277428 | facilitated trehalose transporter Tret | Up | - |
| LOC118277941 | UTP--glucose-1-phosphate uridylyl transferase | Up | - |
| LOC118269585 | glucose transporter type | Up | Up |
| LOC118280239 | dehydrogenase | Up | Up |
| LOC118280972 | UDP-N-acetyl hexosamine pyrophosphorylase-1 | Up | Up |
| LOC118262085 | acetyl-coenzyme A synthetase | - | Down |
| LOC118266444 | 3-ketoacyl-CoA thiolase | Up | - |
| LOC118280410 | acetyl-CoA carboxylase | Up | Up |
| LOC118267788 | connectin | - | Up |
| LOC118266022 | hydroxymethylglutaryl-CoA synthase | - | Down |
| LOC118281649 | 2-oxoglutarate dehydrogenase | - | Up |
| LOC118278502 | ATP-citrate synthase | Up | - |
| LOC118262464 | isocitrate dehydrogenase [NADP] cytoplasmic | Down | Down |
| LOC118266957 | 4-aminobutyrate aminotransferase | Up | Up |
| LOC118272711 | gamma-aminobutyric acid receptor | Up | Up |
| LOC118262156 | glutamate synthase [NADH] | Up | Up |
| LOC118265687 | glutamyl-tRNA (Gln) amidotransferase subunit B | Up | Up |
| LOC118282360 | asparagine synthetase | Down | Down |
| LOC118268389 | acetylcholine receptor subunit alpha | Up | Up |
| LOC118276145 | muscarinic acetylcholine receptor DM1 | Up | Up |
| LOC118282418 | catalase | Down | Down |
| LOC118266989 | superoxide dismutase | Up | Up |
| LOC118269785 | glutathione S-transferase 1 | Down | Down |
| LOC118271638 | glutathione S-transferase 2 | Down | Down |
| LOC118268807 | glutathione hydrolase 1 proenzyme | Up | Up |
| LOC118280652 | carbonyl reductase [NADPH] 3 | Down | Down |
| LOC118266432 | leukotriene A-4 hydrolase | - | Down |
| LOC118269281 | glutaryl-CoA dehydrogenase | Up | Up |
| LOC118273421 | tyrosine aminotransferase | Up | Up |
| LOC118267609 | dopamine N-acetyltransferase | - | Up |
| LOC118277744 | venom carboxylesterase-6 | Up | Up |
| LOC118281189 | cytochrome P450 307a | Down | Down |
| LOC118274175 | cytochrome P450 6B6 | Down | Down |
| LOC118270797 | cytochrome P450 4C1 | Up | Up |
| LOC118270458 | cytochrome P450 4c21 | Up | Up |
| LOC118272240 | cytochrome P450 4c3 | Up | Up |
| LOC118282431 | cytochrome P450 4d2 | Up | Up |
| LOC118266763 | cytochrome P450 6a2 | Up | Up |
| LOC118273800 | cytochrome P450 6B2 | Up | Up |
| LOC118262785 | cytochrome P450 6B5 | Up | Up |
| LOC118264635 | cytochrome P450 9e2 | Up | Up |
| LOC118271250 | cytochrome P450 301a1 | Up | Up |
| LOC118279412 | UDP-glucuronosyltransferase 2B15 | Up | Up |
| LOC118269333 | UDP-glucuronosyltransferase 2B19 | Up | Up |
| LOC118279357 | UDP-glucuronosyltransferase 2B7 | Up | Up |
| Metablite ID | Metabolite Name | LC10 Group | LC90 Group |
|---|---|---|---|
| neg_127 | D-Glucose 6-phosphate | Up | Up |
| neg_149 | Dehydroascorbic acid | - | Up |
| neg_39 | Tyrosine | - | Down |
| neg_399 | Arginine | Up | Up |
| neg_412 | Leucine | Up | Up |
| neg_58 | NADP+ | - | Up |
| neg_76 | Glutamate | Up | Up |
| pos_136 | L-Valine | Down | Down |
| pos_194 | Aspartic acid | Up | Up |
| pos_198 | Tryptophan | - | Up |
| pos_379 | Arachidyl carnitine | - | Up |
| pos_517 | delta. -Tocopherol | - | Up |
| pos_613 | DL-Phenylalanine | - | Down |
| pos_732 | Glutethimide | - | Down |
| pos_752 | Acetylcholine | - | Down |
| pos_766 | Allysine | Down | Down |
| pos_79 | NAD | - | UP |
| pos_904 | Epigallocatechin gallate | - | Down |
| pos_99 | L-Isoleucine | - | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Z.; Batool, R.; Xie, W.; Huang, X.; Wang, Z. Transcriptome and Metabolome Analysis Reveals the Importance of Amino-Acid Metabolism in Spodoptera Frugiperda Exposed to Spinetoram. Insects 2022, 13, 852. https://doi.org/10.3390/insects13090852
Gao Z, Batool R, Xie W, Huang X, Wang Z. Transcriptome and Metabolome Analysis Reveals the Importance of Amino-Acid Metabolism in Spodoptera Frugiperda Exposed to Spinetoram. Insects. 2022; 13(9):852. https://doi.org/10.3390/insects13090852
Chicago/Turabian StyleGao, Zupeng, Raufa Batool, Weifeng Xie, Xiaodan Huang, and Zhenying Wang. 2022. "Transcriptome and Metabolome Analysis Reveals the Importance of Amino-Acid Metabolism in Spodoptera Frugiperda Exposed to Spinetoram" Insects 13, no. 9: 852. https://doi.org/10.3390/insects13090852
APA StyleGao, Z., Batool, R., Xie, W., Huang, X., & Wang, Z. (2022). Transcriptome and Metabolome Analysis Reveals the Importance of Amino-Acid Metabolism in Spodoptera Frugiperda Exposed to Spinetoram. Insects, 13(9), 852. https://doi.org/10.3390/insects13090852