




The Effect of Feeding with Central European Local Mulberry Genotypes on the Development and Health Status of Silkworms and Quality Parameters of Raw Silk


 , ,
, ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mulberry Material for Silkworm Feeding Experiment
- (A)
- Slovenian mulberry genotypes: selected, locally adapted genotypes, obtained by cuttings from old (trunk diameter > 180 cm) Slovenian M. alba trees (n = 10);
- (B)
- Hungarian mulberry genotypes: selected, locally adapted genotypes, obtained by cuttings from old (trunk diameter > 180 cm) Hungarian M. alba trees (n = 16);
- (C)
- reference sericultural M. alba varieties, obtained from the mulberry gene bank of the CREA-AA (‘Kokusou-20’, ‘Morettiana’, ‘Florio’, ‘Giazzola’);
- (D)
- varieties of M. alba, M. alba × rubra, M. nigra and M. australis grown for fruit production.
2.2. Determination of the Chemical Composition of Mulberry Leaves
2.3. Silkworm Rearing
2.3.1. Silkworm Genetic Material
2.3.2. Silkworm Rearing Technology
2.3.3. Monitoring the Health and Microbiological Status of Silkworm Larvae
Molecular Detection of Bombyx Mori Nucleopolyhedrovirus
Identification of Gut Microbiome and Potential Bacterial Pathogens by Metagenomic Studies
Comparison of Selected Qualitative and Quantitative Hemogram Parameters in the Main Feeding Groups of Larvae
2.4. Methods for Evaluation of Quality of Silk Cocoons
2.5. Statistical Data Analysis
3. Results
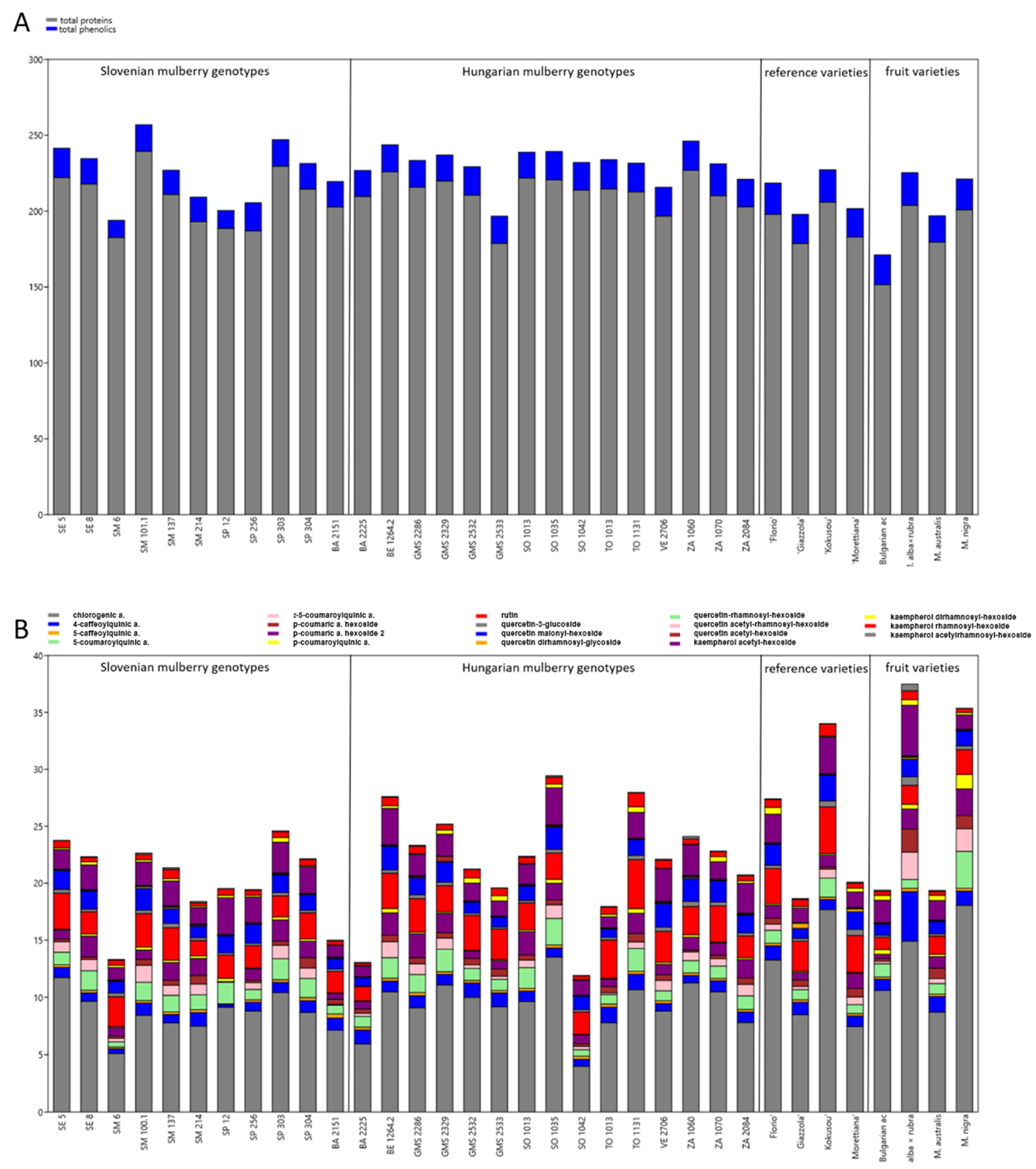
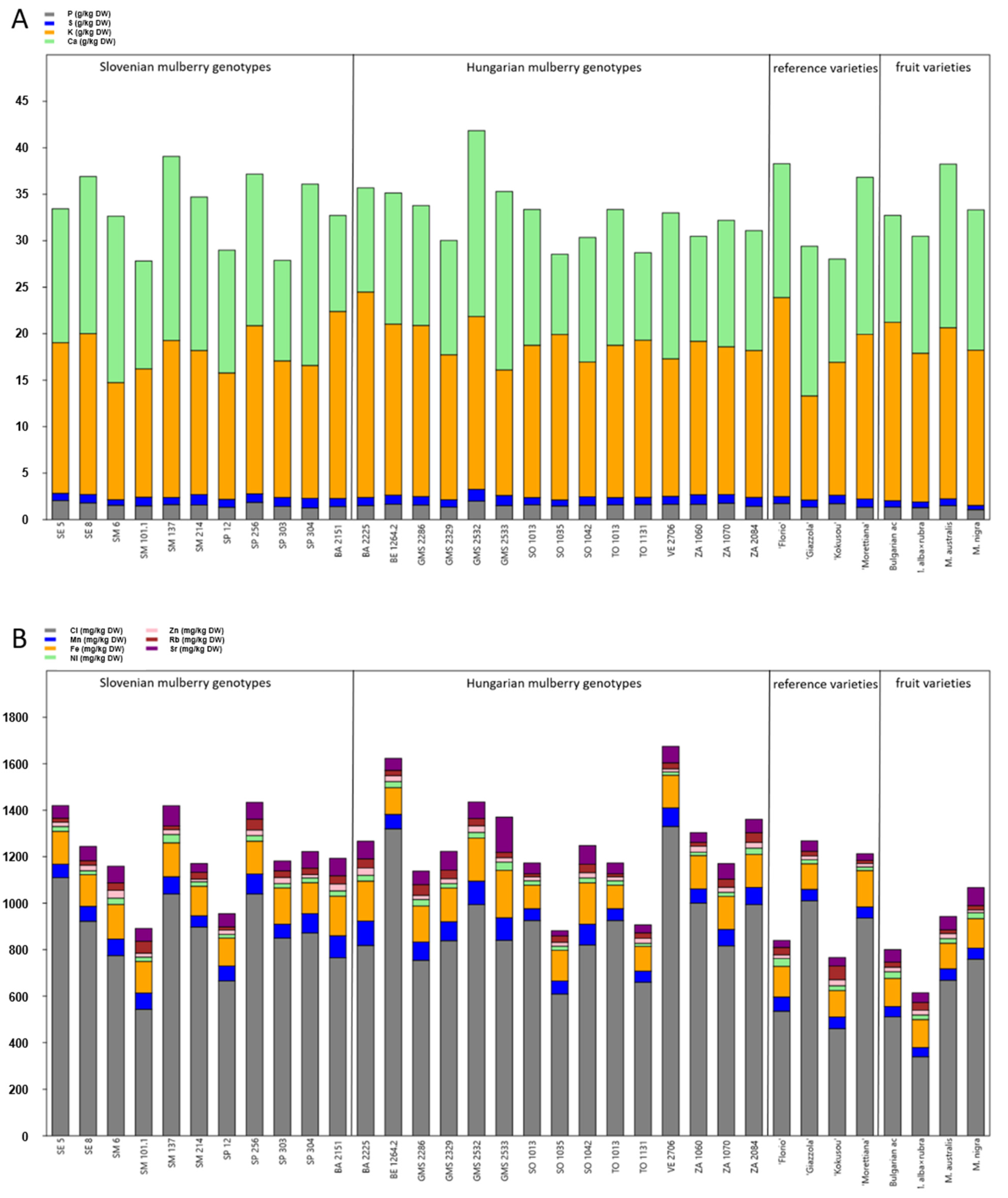
3.1. Chemical Composition of the Leaves of Selected Slovenian and Hungarian Mulberry Genotypes, Reference Sericultural and Fruit Varieties
3.2. Origin Dependent Differences in Chemical Composition of Mulberry Leaves
3.3. Description of the Influence of Feeding Silkworms with Leaves of Various Genotypes of Mulberry Trees on Larval Development
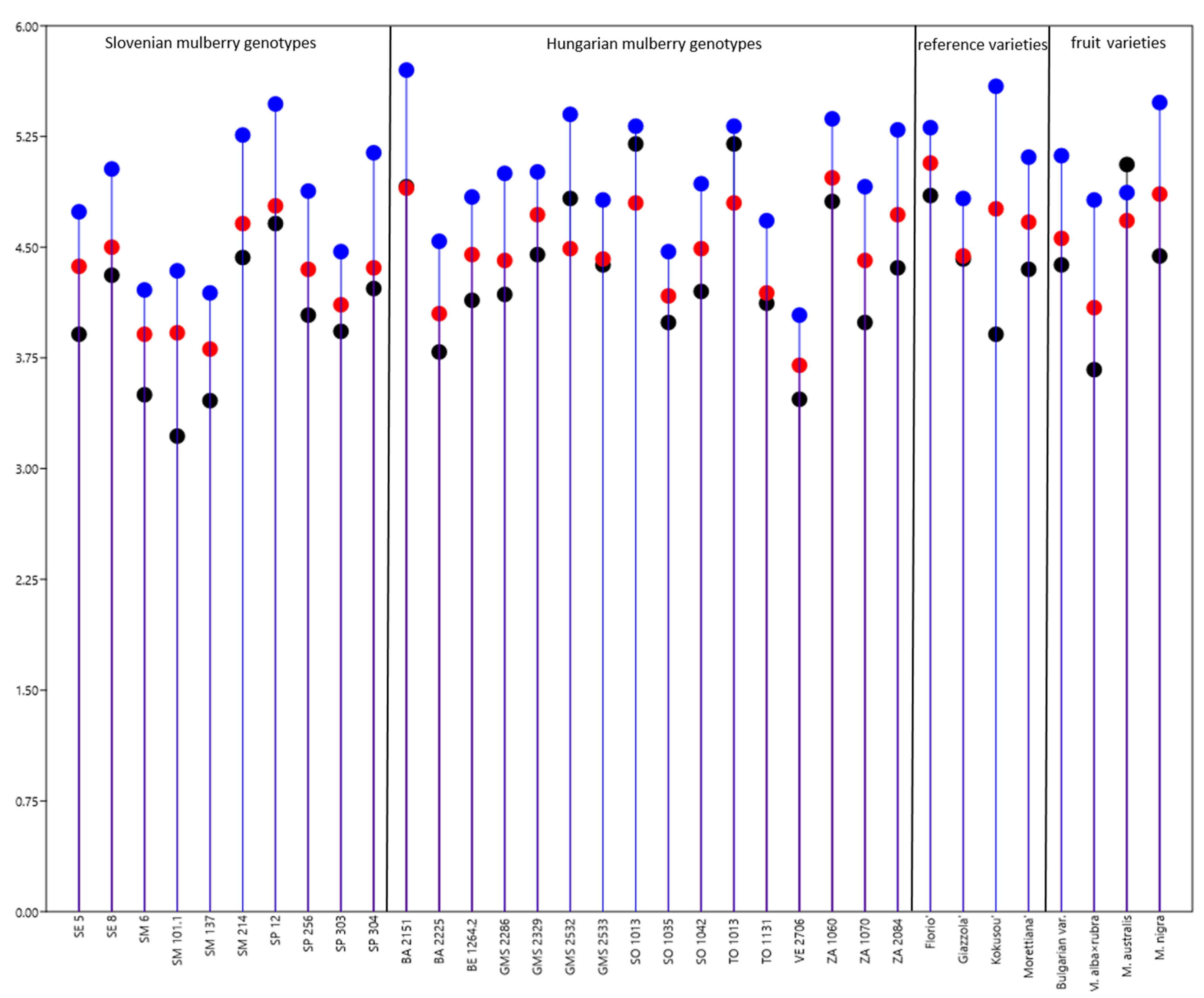
3.4. Origin Dependent Differences in the Growth of the Silkworm Larvae and Cocoon Weight
3.5. Description of the Influence of Feeding Silkworms with Leaves of Various Mulberry Genotypes on the Quality of Raw Silk
3.6. Origin Dependent Differences in the Quality of Raw Silk
3.7. Correlation between Mulberry Metabolites, Silkworm, and Raw Silk Parameters
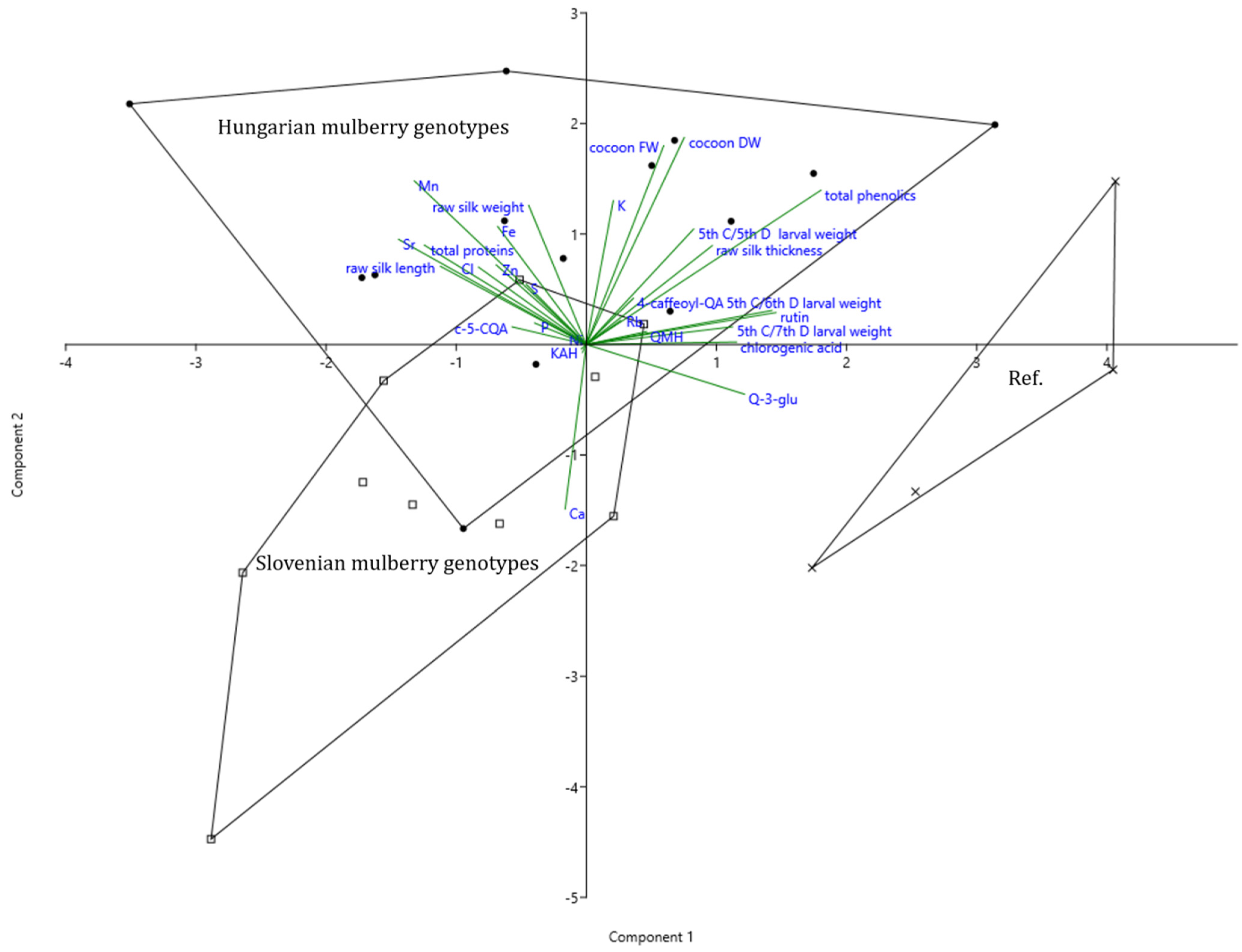
3.8. Principal Component Analysis of Mulberry Metabolites, Silkworm, and Raw Silk Parameters
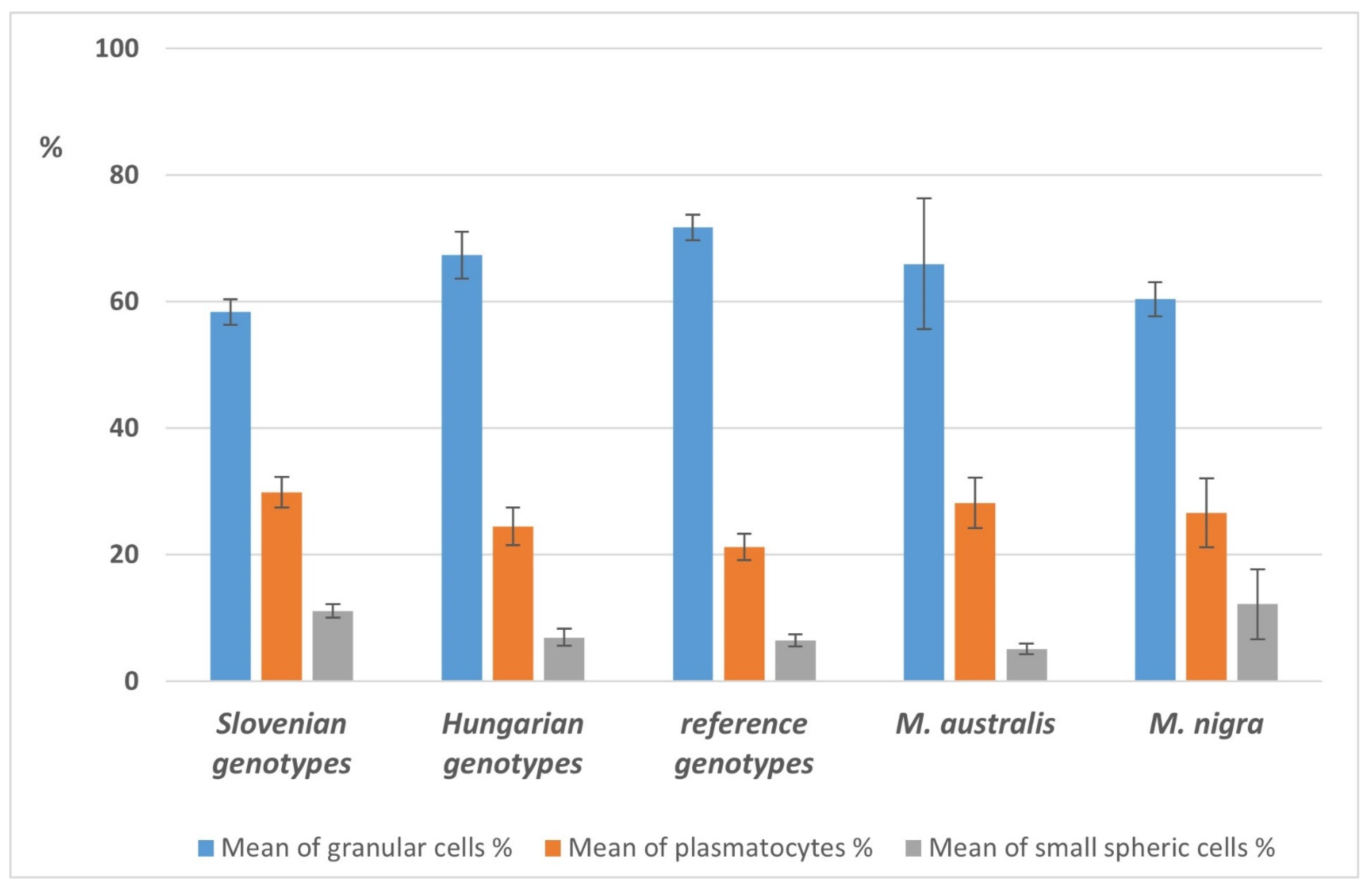
3.9. Health and Microbiological Status of Silkworm Larvae
3.9.1. Molecular Detection of BmNPV by Real-Time PCR
- 1st day, third instar: 1 dead (liquid rotting) larva; cT mean 40.63, standard deviation (SD) 0.84;
- 2nd day, third instar: 1 cachectic larva; cT mean 40.70, SD 1.49;
- 1st day, fourth instar: 1 larva died during moulting, showed signs of Grasserie; cT mean 18.32, SD 0.17.
- In the fifth instar, BmNPV DNA was detected in three out of the 1340 larvae (0.22%). Positive larvae were ill or were found dead:
- 1st day: Hu-BE 1264/2 (signs of Grasserie); cT mean 37.38, cT SD 0.69;
- end of 5th instar: Hu-ZA 2084.2 (moribund larva with black stripes), cT mean 20-09, SD 0.14;
- end of 5th instar: Hu-mix (dead larva, signs of Grasserie), CT mean 37.43, SD 0.92.
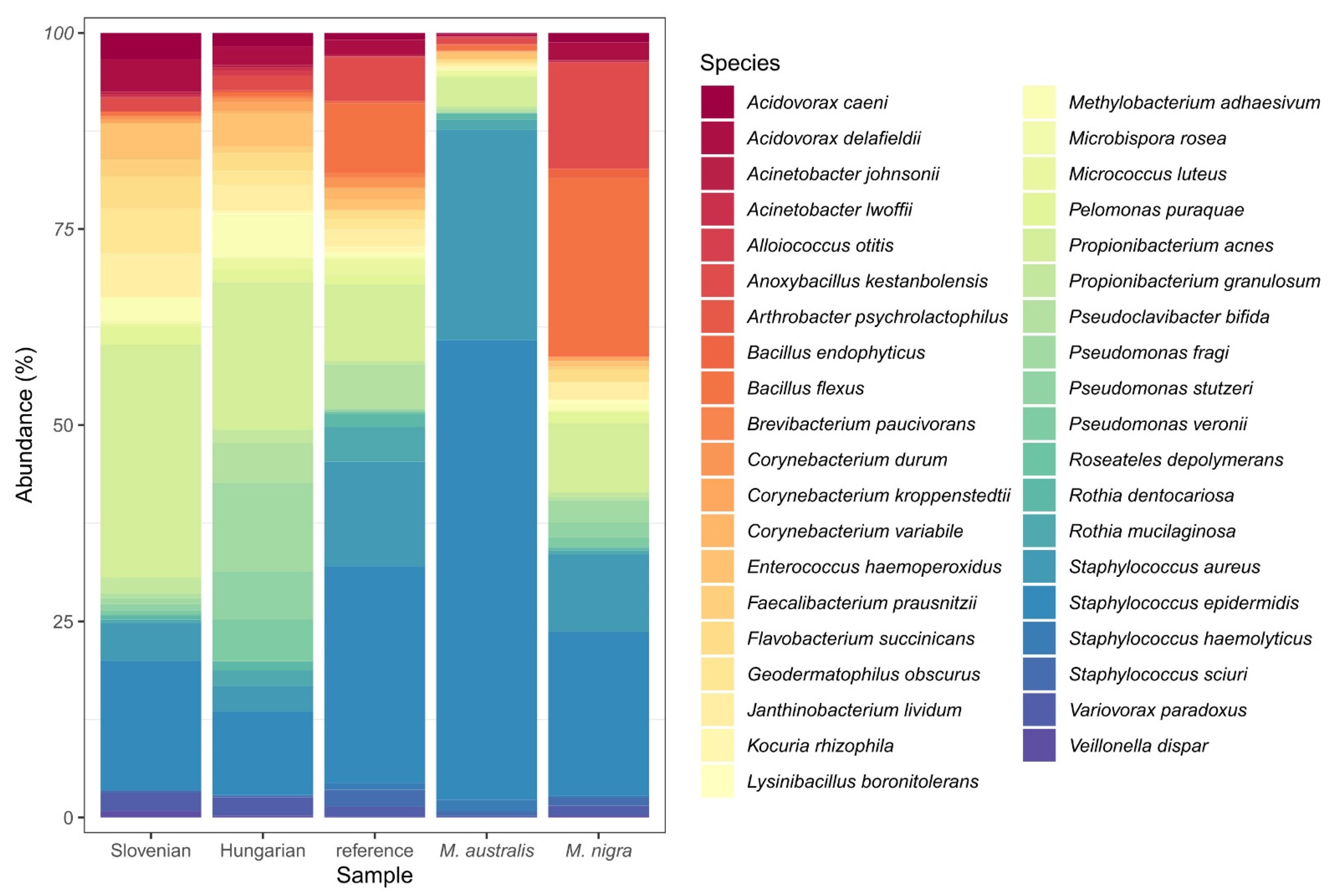
3.9.2. Identification of Gut Microbiome and Potential Bacterial Pathogens by Metagenomic Studies
- (1)
- 57% of representatives of Chloroplast (99% Streptophyta) were found by Slovenian genotypes and 47% by Hungarian genotypes, almost half less (22%) by reference genotypes, while M. australis and M. nigra had only 16% and 12%.
- (2)
- The range was quite opposite by class Betaproteobacteria (mainly representatives from order Burkholderiales): 44% by M. nigra, 39% by reference genotypes, 24% by Slovenian genotypes, 22% by Hungarian genotypes, and 18% by M. australis.
- (3)
- The representatives from Alphaproteobacteria (99% of representatives coming from order Rickettsiales) were in the following ranges: M. nigra and Hungarian genotypes 8%, Slovenian genotypes 6%, reference genotypes 3%, and M. australis 2% (representatives mainly from orders: Rhodobacterales 40%, Rhizobiales 24%, Sphingomonadales 12%, Rhodospirillales 12%, 9% Rickettsiales).
- (4)
- The representatives from class Gammaproteobacteria were in following ranges: Hungarian mulberry genotypes 4% (Pseudomonadales 3%, Enterobacteriaceae 1%, Xanthomonadales 0.1%), M. nigra 3% (Pseudomonadales 2%, Enterobacteriaceae 0.07%, Xanthomonadales 0.1%, Oceanospirillales 0.1%, Alteromonadales 0.1%), Slovenian genotypes 1% (Pseudomonadales 1%, Enterobacteriaceae 0.06%), reference genotypes 1% (Pseudomonadales 0.7%, Enterobacteriaceae 0.4%), M. australis 1% (Pseudomonadales 0.8%, Enterobacteriaceae 0.3%).
- (5)
- The representatives from class Actinobacteria were in the following ranges: reference genotypes 14%, Hungarian genotypes 9%, M. australis 9%, M. nigra 5%, and Slovenian genotypes 4%.
- (6)
- The representatives from class Bacilli were in the following ranges: Slovenian genotypes 4%, Hungarian genotypes 9%, reference genotypes 14%, M. australis 9%, and M. nigra 5%.
- (7)
- The representatives from class Clostridia were in the following ranges: M. nigra 3%, M. australis 2%, Slovenian genotypes 1%, Hungarian genotypes 1%, reference genotypes 0.8%.
- (8)
- The representatives from class Flavobacteriales were in the following ranges: M. nigra 3%, M. australis 2%, Slovenian genotypes 1%, Hungarian genotypes 1%, reference genotypes 0.8%.
- (9)
- The representatives from other classes were in the following ranges: Slovenian genotypes 4%, Hungarian genotypes 3%, reference genotypes 2.2%, M. nigra 2%, M. australis 1.2 %.
3.9.3. Comparison of Selected Qualitative and Quantitative Hemogram Parameters in the Main Feeding Groups of Larvae
4. Discussion
4.1. The Influence of Proteins and Phenolics on Larval, Cocoon and Raw Silk Parameters
4.2. The Influence of Macro- and Microelements on Larval, Cocoon and Raw Silk Parameters
4.3. Correlations between Test Parameters in Multivariate Analysis
4.4. Development and Health Status of Larvae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inserco 2022, International Sericultural Commission Web Page. Available online: https://www.inserco.org/en/ (accessed on 13 September 2022).
- Žontar, J. Svilogojstvo in Svilarstvo na Slovenskem od 16. do 20. Stoletja; SAZU, Razred za Zgodovinske in Družbene Vede: Ljubljana, Slovenia, 1957; pp. 1–160. [Google Scholar]
- Žontar, J. Gojitev sviloprejk. In Gospodarska in Družbena Zgodovina Slovencev: Zgodovina Agrarnih Panog; I. Zvezek Agrarno Gospodarstvo; Blaznik, P., Grafenauer, B., Vilfan, S., Eds.; DZS: Ljubljana, Slovenia, 1970; pp. 409–415. [Google Scholar]
- Papp, Z. A selyemhernyó-tenyésztés történeti áttekintése és újrahonosításának időszerűsége. Erdészeti Lapok 2000, 85, 1–12. [Google Scholar]
- Tzenov, P.; Cappellozza, S.; Saviane, A. Black, Caspian Seas and Central Asia Silk Association (BACSA) for the Future of Sericulture in Europe and Central Asia. Insects 2022, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Urbanek Krajnc, A.; Ugulin, T.; Paušič, A.; Rabensteiner, J.; Bukovac, V.; Mikulič Petkovšek, M.; Janžekovič, F.; Bakonyi, T.; Berčič, R.L.; Felicijan, M. Morphometric and biochemical screening of old mulberry trees (Morus alba L.) in the former sericulture region of Slovenia. Acta Soc. Bot. Pol. 2019, 88, 3614. [Google Scholar] [CrossRef]
- Šelih, M.; Mikulič Petkovšek, M.; Krajnc, D.; Berčič, R.L.; Urbanek Krajnc, A. Screening of leaf metabolites in historical mulberry trees (Morus alba L.) from different eco-geographical regions of Slovenia 2020. Trees 2020, 34, 971–986. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitation of icrogram quantities of protein utilizing the principle of proteindye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Nečemer, M.; Kump, P.; Ščančar, J.; Jaćimović, R.; Simčič, J.; Pelicon, P.; Budnar, M.; Jeran, Z.; Pongrac, P.; Regvar, M.; et al. Application of X-ray fluorescence analytical techniques in phytoremediation and plant biology studies. Spectrochim. Acta Part B 2008, 63, 1240–1247. [Google Scholar] [CrossRef]
- Nečemer, M.; Kump, P.; Vogel-Mikuš, K. Energy Dispersive X-ray Fluorescence Analysis of Biological Materials. In X-ray Fluorescence in Biological Sciences: Principles, Instrumentation, and Applications; Singh, V.K., Kawai, J., Tripathi, D.K., Eds.; John Wiley & Sons: Newark, NJ, USA, 2022; pp. 311–325. [Google Scholar] [CrossRef]
- Kump, P.; Nečemer, M.; Rupnik, Z.; Pelicon, P.; Ponikvar, D.; Vogel-Mikuš, K. Improvement of the XRF quantification and enhancement of the combined applications by EDXRF and micro-PIXE. In Integration of Nuclear Spectrometry Methods as a New Approach to Material Research; International Atomic Energy Agency: Vienna, Austria, 2012; pp. 101–109. Available online: https://www.iaea.org/publications/8423/integration-of-nuclear-spectrometry-methods-as-a-new-approach-to-material-research (accessed on 13 September 2022).
- Brion, H. Praxishandbuch Seidenraupen-Aufzucht, 2nd ed.; Vereinigung Schweizer Seidenproduzenten: Wohlen bei Bern, Switzerland, 2016; p. 19. [Google Scholar]
- Cappellozza, S. (Council for Agricultural Research and Economics, Research Center for Agriculture and Environment, Padua, Italy). Personal communication, 2015. [Google Scholar]
- Gani, M.; Chouhan, S.; Lal, B.; Gupta, R.K.; Khan, G.; Kumar, N.B.; Saini, P.; Ghosh, M.K. Bombyx mori nucleopolyhedrovirus (BmBPV): Its impact on silkworm rearing and management strategies. J. Biol. Control 2017, 31, 189–193. [Google Scholar] [CrossRef]
- Khurad, A.M.; Mahulikara, A.; Rathod, M.K.; Rai, M.M.; Kanginakudru, S.; Nagaraju, J. Vertical transmission of nucleopolyhedrovirus in the silkworm, Bombyx mori L. J. Invertebr. Pathol. 2004, 87, 8–15. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequence (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2005, 33, D501–D504. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Australia, 2021; Available online: http://www.R-project.org/ (accessed on 13 September 2022).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Nittono, Y.; Tomabechi, S.; Onodera, N. Formation of haemocytes near the imaginal wingdisc in the silkworm, Bombyx mori L. J. Sericult. Sci. Jpn. 1964, 33, 43–45. [Google Scholar] [CrossRef]
- Yamashita, M.; Iwabuchi, K. Bombyx mori prohemocyte division and differentiation in individual microcultures. J. Insect Physiol. 2001, 47, 325–331. [Google Scholar] [CrossRef]
- Lee, Y.W. Silk Reeling and Testing Manual; FAO Agricultural Services Bulletin, No. 136; Food and Agriculture Organization of the United Nations: Rome, Italy, 1999; p. 139. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 13 September 2022).
- Andadari, L.; Minarningsih, M.; Suwandi. The effect of feeding various species of mulberry (Morus spp.) on the growth of silkworm and quality of cocoon hybrid BS 09. IOP Conf. Ser. Earth Environ. Sci. 2021, 914, 012017. [Google Scholar] [CrossRef]
- Kumar, R.V.; Chauhan, S.; Kumar, D.; More, N. Nutritional composition in leaves of some mulberry varieties: A comparative study. In Proceedings of the International Conference of Bioinformatics and Biomedical Technology, Chengdu, China, 16–18 April 2010; pp. 438–442. [Google Scholar] [CrossRef]
- Radjabi, R. Effect of mulberry leaves enrichment with amino acid supplementary nutrients on silkworm, Bombyx mori L. at north of Iran. Acad. J. Entomol. 2010, 3, 45–51. [Google Scholar]
- Wang, C.; Yang, F.; Wang, Q.; Zhou, X.; Xie, M.; Kang, P.; Wang, Y.; Peng, X. Nutritive Value of Mulberry Leaf Meal and its Effect on the Performance of 35-70-Day-Old Geese. Int. J. Poult. Sci. 2016, 54, 41–46. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rajabi, K.; Ebadi, R.; Mirhosseini, R.; Seidavi, S.Z.; Zolfaghari, M.; Etebari, K. A review on nutritive effect of mulberry leaves enrichment with vitamins on economic traits and biological parameters of silkworm Bombyx mori L. Invert. Surviv. J. 2007, 4, 86–91. [Google Scholar]
- Mendonça, G.A.; de Marchini, L.C.; Macedo, L.P.M. Cocoon production of the silkworm, Bombyx mori L. (Lepidoptera: Bombycidae), fed on leaves of mulberry hybrids. Caatinga 2010, 23, 118–122. [Google Scholar]
- El-Banna, A.A.; Moustafa, M.N.; Mahmoud, S.M.; El-Shafei, A.M.; Moustafa, A.A. Effect of feeding different mulberry varieties on some biological characteristics of the silkworm, Bombyx mori L. Egypt J. Pure Appl. Sci. 2013, 51, 55–60. [Google Scholar] [CrossRef]
- Adeduntan, S. Influence of different varieties of mulberry leaves (Morus alba) on growth and cocoon performance of biovoltine strain of silkworm (Bombyx mori). Int. J. Biol. Chem. Sci. 2015, 9, 751–757. [Google Scholar] [CrossRef]
- Radojković, M.M.; Zeković, Z.P.; Vidović, S.S.; Kočar, D.D.; Mašković, P.Z. Free radical scavenging activity and total phenolic and flavonoid contents of mulberry (Morus spp. L., Moraceae) extracts. Hem. Ind. 2012, 66, 552. [Google Scholar] [CrossRef]
- Chung, H.I.; Kim, J.; Kim, J.Y.; Kwon, O. Acute intake of mulberry leaf aqueous extract affects postprandial glucose response after maltose loading: Randomized double-blind placebo-controlled pilot study. J. Funct. Foods 2013, 5, 1502–1506. [Google Scholar] [CrossRef]
- Hunyadi, A.; Liktor-Busa, E.; Márki, A.; Martins, A.; Jedlinszki, N.; Hsieh, T.J.; Báthori, M.; Hohmann, J.; Zupkó, I. Metabolic Effects of Mulberry Leaves: Exploring Potential Benefits in Type 2 Diabetes and Hyperuricemia. Evid.-Based Complement. Altern. Med. 2013, 2013, 948627. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Flaczyk, E.; Jeszka, J.; Krejpcio, Z.; Król, E.; Buchowski, M.S. Mulberry leaf extract intake reduces hyperglycaemia in streptozotocin (STZ)-induced diabetic rats fed high-fat diet. J. Funct. Foods 2014, 8, 9–17. [Google Scholar] [CrossRef]
- Chen, H.; He, X.; Liu, Y.; Li, J.; He, Q.; Zhang, C.; Wei, B.; Zhang, Y.; Wang, J. Extraction, purification and anti-fatigue activity of γ-aminobutyric acid from mulberry (Morus alba L.) leaves. Sci. Rep. 2016, 6, 18933. [Google Scholar] [CrossRef] [PubMed]
- Gryn-Rynko, A.; Bazylaka, G.; Olszewska-Slonina, D. New potential phytotherapeutics obtained from white mulberry (Morus alba L.) leaves. Biomed. Pharmacother. 2016, 84, 628–636. [Google Scholar] [CrossRef]
- Sugiyama, M.; Katsube, T.; Koyama, A.; Itamura, H. Seasonal Changes in Functional Component Contents in Mulberry (Morus alba L.) Leaves. Hortic. J. 2017, 86, 534–542. [Google Scholar] [CrossRef]
- Ju, W.-T.; Kwon, O.-C.; Kim, H.-B.; Sung, G.-B.; Kim, H.-W.; Kim, Y.-S. Qualitative and quantitative analysis of flavonoids from 12 species of Korean mulberry leaves. J. Food Sci. Technol. 2018, 55, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.-T.; Kwon, O.-C.; Lee, M.-K.; Kim, H.-B.; Sung, G.-B.; Kim, Y.-S. Quali-Quantitative Analysis of Flavonoids for Mulberry Leaf and Fruit of ‘Suhyang’. Korean J. Environ. Agric. 2017, 36, 249–255. [Google Scholar] [CrossRef]
- Memon, A.A.; Memon, N.; Luthria, D.L.; Bhanger, M.I.; Pitafi, A.A. Phenolic acids profiling and antioxidant potential of mulberry (Morus laevigata W., Morus nigra L., Morus alba L.) leaves and fruits grown in Pakistan. Pol. J. Food Nutr. Sci. 2010, 60, 25–32. [Google Scholar]
- Zou, Y.; Liao, S.; Shen, W.; Liu, F.; Tang, C.; Chen, C.-Y.O.; Sun, Y. Phenolics and Antioxidant Activity of Mulberry Leaves Depend on Cultivar and Harvest Month in Southern China. Int. J. Mol. Sci. 2012, 13, 16544–16553. [Google Scholar] [CrossRef]
- Iqbal, S.; Younas, U.; Sirajuddin; Chan, K.W.; Sarfraz, R.A.; Uddin, K. Proximate Composition and Antioxidant Potential of Leaves from Three Varieties of Mulberry (Morus sp.): A Comparative Study. Int. J. Mol. Sci. 2012, 13, 6651–6664. [Google Scholar] [CrossRef]
- Lee, W.J.; Choi, S.W. Quantitative Changes of Polyphenolic Compounds in Mulberry (Morus alba L.) Leaves in Relation to Varieties, Harvest Period and Heat Processing. Prev. Nutr. Food Sci. 2012, 17, 280–285. [Google Scholar] [CrossRef]
- Thabti, I.; Elfalleh, W.; Hannachi, H.; Ferchichi, A.; Campos, M.D.G. Identification and quantification of phenolic acids and flavonol glycosides in Tunisian Morus species by HPLC-DAD and HPLC–MS. J. Funct. Foods 2012, 4, 367–374. [Google Scholar] [CrossRef]
- Flaczyk, E.; Kobus-Cisowska, J.; Przeor, M.; Korczak, J.; Remiszewski, M.; Korbas, E.; Buchowski, M. Chemical characterization and antioxidative properties of Polish variety of Morus alba L. leaf aqueous extracts from the laboratory and pilot-scale processes. J. Agric. Sci. 2013, 4, 141–147. [Google Scholar] [CrossRef]
- Choi, S.W.; Lee, Y.J.; Ha, S.B.; Jeon, Y.H.; Lee, D.H. Evaluation of Biological Activity and Analysis of Functional Constituents from Different Parts of Mulberry (Morus alba L.) Tree. J. Korean Soc. Food Sci. Nutr. 2015, 6, 823–831. [Google Scholar] [CrossRef]
- Germanò, M.P.; D’Angelo, V.; Catania, S.; Miano, T.C.; Perna, V.; Farago, S.; Cappellozza, L.; Cappellozza, S. Phenolic content of leaf of different mulberry cultivars affects growth in the silkworm. In Proceedings of the 21st International Sericultural Congress of ISC, Athens, Greece, 3–6 November 2008; pp. 25–29. [Google Scholar]
- Hirayama, C.; Ono, H.; Tamura, Y.; Konno, K.; Nakamura, M. Regioselective formation of quercetin 5-O-glucoside from orally administered quercetin in the silkworm, Bombyx mori. Phytochemistry 2008, 69, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Daimon, T.; Hirayama, C.; Kanai, M.; Ruike, Y.; Meng, Y.; Kosegawa, E.; Nakamura, M.; Tsujimoto, G.; Katsuma, S.; Shimada, T. The silkworm Green b locus encodes a quercetin 5-O-glucosyltransferase that produces green cocoons with UV-shielding properties. Proc. Natl. Acad. Sci. USA 2010, 107, 11471–11476. [Google Scholar] [CrossRef]
- Vihakas, M. Flavonoid and Other Phenolic Compounds: Characterization and Interactions with Lepidopteran and Sawfly Larvae. Doctoral Thesis, University of Turku, Turku, Finland, 2014. [Google Scholar]
- Kusurkar, T.S.; Tandon, I.; Sethy, N.K.; Bhargava, K.; Sarkar, S.; Singh, S.K.; Das, M. Fluorescent silk cocoon creating fluorescent diatom using a “Water glass-fluorophore ferry”. Sci. Rep. 2013, 3, 3290. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T. Basic and Applied Genetics of Silkworms. In Silkworm Biofactory; Silk to, Biology; Maenaka, K., Park, E.Y., Eds.; CRC Press: Boca Raton, FL, USA, 2018; p. 15. [Google Scholar] [CrossRef]
- Srivastava, S.; Kapoor, R.; Thathola, A.; Srivastava, R.P. Nutritional quality of leaves of some genotypes of mulberry (Morus alba). Int. J. Food Sci. Nutr. 2006, 57, 305–313. [Google Scholar] [CrossRef]
- Al-kirshi, R.A.; Alimon, A.R.; Zulkifli, I.; Zahari, M.W.; Sazili, A.Q. The chemical composition and nutritive value of mulberry leaf as a protein source in poultry diets. In Feed and Nutrition: The 1st International Seminar on Animal Industry, Bogor, Indonesia; Bogor Agricultural University: Bogor, Indonesia, 2009; pp. 98–102. [Google Scholar]
- Kumar, K.; Mohan, M.; Tiwari, N.; Kumar, S. Production potential and leaf quality evaluation of selected mulberry (Morus alba) clones. J. Pharmacogn. Phytochem. 2018, 7, 482–486. [Google Scholar]
- Sánchez-Salcedo, E.M.; Mena, P.; García-Viguera, C.; Martínez, J.J.; Hernández, F. Phytochemical evaluation of white (Morus alba L.) and black (Morus nigra L.) mulberry fruits, a starting point for the assessment of their beneficial properties. J. Funct. Foods 2015, 12, 399–408. [Google Scholar] [CrossRef]
- Yamagishi, T.; Endo, H.; Fukumura, K.; Nagata, S.; Hayakawa, T.; Adegawa, S.; Kasubuchi, M.; Sato, R. Glucose, some amino acids and a plant secondary metabolite, chlorogenic acid induce the secretion of a regulatory hormone, tachykinin-related peptide, from the silkworm midgut. Peptides 2018, 106, 21–27. [Google Scholar] [CrossRef]
- Kato, M.; Yamada, H. Chlorogenic acid as a growth factor of silkworm. J. Silkworm 1964, 16, 85. [Google Scholar] [CrossRef]
- Naito, K.; Hayashiya. Studies on the micro constituents in mulberry leaves. IV. Isolation of chlorogenic acid. J. Agric. Chem. Soc. Jpn. 1965, 39, 237–238. [Google Scholar]
- Hamamura, Y.; Kuwata, K.; Masuda, H. Effect of Gallic Acid on the Growth of the Silkworm Larvae Bombyx mori L. Nature 1966, 212, 1386–1387. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Kato, M. Chlorogenic Acid as an Indispensable Component of the Synthetic Diet for the Silkworm. Proc. Jpn. Acad. 1966, 42, 399–403. [Google Scholar] [CrossRef]
- Koike, S.; Iizuka, T.; Mizutani, J. Determination of Caffeic Acid in the Digestive Juice of Silkworm Larvae and Its Antibacterial Activity against the Pathogenic Streptococcus faecalis AD–4. Agric. Biol. Chem. 1979, 43, 1727–1731. [Google Scholar] [CrossRef]
- Naik, D.; Patil, G.M.; Biradar, R. Impact of fortification of chlorogenic acid rich botanical on silkworm growth and cocoon parameters of silkworm Bombyx mori L. Plant Arch. 2017, 17, 661–665. [Google Scholar]
- Jarienė, E.; Levickienė, D.; Danilčenko, H.; Vaitkevičienė, N.; Kulaitienė, J.; Jakštas, V.; Ivanauskas, L.; Gajewski, M. Effects of biodynamic preparations on concentration of phenolic compounds in the leaves of two white mulberry cultivars. Biol. Agric. Hortic. 2018, 35, 132–142. [Google Scholar] [CrossRef]
- Pothinuch, P.; Tongchitpakdee, S. Phenolic Analysis for Classification of Mulberry (Morus spp.) Leaves according to Cultivar and Leaf Age. J. Food Qual. 2019, 2019, 2807690. [Google Scholar] [CrossRef]
- Kurioka, A.; Yamazaki, M. Purification and Identification of Flavonoids from the Yellow Green Cocoon Shell (Sasamayu) of the Silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2002, 66, 1396–1399. [Google Scholar] [CrossRef]
- Kurioka, A.; Yamazaki, M. Antioxidant in the cocoon of the silkworm, Bombyx mori. J. Insect Biotechnol. Sericol. 2002, 71, 177–180. [Google Scholar] [CrossRef]
- Preiß, S.; Degenhardt, J.; Gershenzon, J. Plant animal dialogues. In Ecological Biochemistry: Environmental and Interspecies Interactions; Krauss, G.J., Nies, D.H., Eds.; Wiley-Blackwell: Oxford, UK, 2015; pp. 313–330. ISBN 978-3-527-31650-2. [Google Scholar]
- Zhang, Y.-E.; Ma, H.-J.; Feng, D.-D.; Lai, X.-F.; Chen, Z.-M.; Xu, M.-Y.; Yu, Q.-Y.; Zhang, Z. Induction of Detoxification Enzymes by Quercetin in the Silkworm. J. Econ. Èntomol. 2012, 105, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Enkhmaa, B.; Shiwaku, K.; Katsube, T.; Kitajima, K.; Anuurad, E.; Yamasaki, M.; Yamane, Y. Mulberry (Morus alba L.) Leaves and Their Major Flavonol Quercetin- 3-(6-Malonylglucoside) Attenuate Atherosclerotic Lesion Development in LDL Receptor-Deficient Mice. J. Nutr. 2005, 135, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Katsube, T.; Imawaka, N.; Kawano, Y.; Yamazaki, Y.; Shiwaku, K.; Yamane, Y. Antioxidant flavonol glycosides in mulberry (Morus alba L.) leaves isolated based on LDL antioxidant activity. Food Chem. 2006, 97, 25–31. [Google Scholar] [CrossRef]
- Katsube, T.; Tsurunaga, Y.; Sugiyama, M.; Furuno, T.; Yamasaki, Y. Effect of air-drying temperature on antioxidant capacity and stability of polyphenolic compounds in mulberry (Morus alba L.) leaves. Food Chem. 2009, 113, 964–969. [Google Scholar] [CrossRef]
- Katsube, T.; Yamasaki, M.; Shiwaku, K.; Ishijima, T.; Matsumoto, I.; Abe, K.; Yamasaki, Y. Effect of flavonol glycoside in mulberry (Morus alba L.) leaf on glucose metabolism and oxidative stress in liver in diet-induced obese mice. J. Sci. Food Agric. 2010, 90, 2386–2392. [Google Scholar] [CrossRef] [PubMed]
- Mahadeva, A. Nutritive Elemental Status in Mulberry (Morus sp.) Foliage under Jassids (Empoasca flavescenes F.) Infestation. Indian J. Nat. Sci. 2016, 7, 11537. [Google Scholar]
- Shifa, K.; Terefe, M.; Tilahun, A.; Ibrahim, A.; Nigusu, Y. Leaf macronutrient composition of mulberry (Morus indica) varieties and its relationship with productivity of mulberry silkworm (Bombyx mori L.). J. Agric. Sci. Res. 2020, 8, 390–398. [Google Scholar]
- Biskin, D.P.; Bloom, A. Mineral Nutrition. In Plant Physiology, 5th ed.; Taiz, L., Zeiger, P.E.E., Eds.; Sinauer Associatiates Publishers: Munderland, MA, USA, 2010; pp. 107–125. [Google Scholar]
- Ito, T.; Nimura, M. Nutrition of the silkworm Bombyx mori L., its specific nutrient requirements and its nutrition in relation to the mineral nutrition of its host plant mulberry, Morus indica L. Indian J. Exptl. Biol. 1966, 4, 31–36. [Google Scholar]
- Chakrabarti, S.; Subramanyam, M.R.; Singhal, B.K.; Datta, R.K. Nutrient deficiency management in mulberry. In Handbook of Mulberry Nutrition; Shankar, M.A., Shetty, G.P., Eds.; Multiplex, Karnataka Agro Chemicals: Bangalore, India, 1997; pp. 19–75. [Google Scholar]
- Miller, A.J. Plant Mineral Nutrition. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2014. [Google Scholar] [CrossRef]
- Munirathnam Reddy, M.; Subbaswamy, M.R.; Sinha, A.K. Sulphur nutrition in mulberry plants. Indian Silk 1990, 29, 45–46. [Google Scholar]
- Mahadevappa, L.; Magadi, S.P.; Prabhuraj, D.K.; Thimmareddy, H.; Bongale, U.D. Available sulphur in some mulberry growing soils of Karnataka. In Nutritional Management and Quality Improvement in Sericulture, Proceedings of the National Seminar on Mulberry Sericulture Research in India, Karnataka, India, 26–28 November 2001; State Sericulture Research and Development Institute: Bangalore, India, 2001; pp. 161–165. [Google Scholar]
- Shree, M.P.; Anuradha, R.; Nagaveni, V. Impact of rust disease on the mineral nutrition of mulberry plants. Sericologia 2005, 45, 115–121. [Google Scholar]
- Shankar, M.A. Handbook of Mulberry Nutrition; Shetty, G.P., Ed.; Multiplex, Karnataka Agro Chemicals: Bangalore, India, 1997. [Google Scholar]
- Lokanath, R.; Shivashankar, K.; Kasiviswanathan, K. Effect of foliar application of magnesium and micronutrients to mulberry on the quality and production of cocoons. Indian J. Seric 1986, 24, 40–45. [Google Scholar]
- Barretto, D.A.; Gadwala, M.; Vootla, S.K. Chapter 1—The silkworm gut microbiota: A potential source for biotechnological applications. Methods Microbiol. 2021, 49, 1–26. [Google Scholar] [CrossRef]
- Nematollahian, S.; Bagheri, M.; Yousefi, Z.; Khezrian, A.; Zahmatkesh, Z. Haematological Changes in Bombyx mori (Lepidoptera: Bombycidae) Infected by Nosema bombycis. J. Agric. Urban Entomol. 2021, 37, 38–47. [Google Scholar] [CrossRef]
- Saxena, S.; Chandra, R.T.; Kallare, P.S.; Arunkumar, K.P. MicroRNAs in the silkworm-pathogen interactions. Methods Microbiol. 2021, 49, 97–113. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Total Proteins | Total PH | Chlorogenic a. | 4-CQA | c-CQA | Rutin | Q-3-glu | QMH | KAH * | |
|---|---|---|---|---|---|---|---|---|---|---|
| Slovenian genotypes | 19 | 211.48 ± 18.51 a | 17.03 ± 2.07 b | 9.07 ± 1.66 c | 0.9 2± 0.30 b | 1.50 ± 0.32 a | 2.32 ± 0.63 b | 0.2 6 ± 0.06 b | 1.51 ± 0.30 ab | 2.15 ± 0.48 a |
| Hungarian genotypes | 25 | 211.89 ± 13.77 a | 18.19 ± 2.02 b | 9.97 ± 1.61 bc | 0.91 ± 0.25 b | 1.52 ± 0.48 a | 2.81 ± 0.64 a | 0.31 ± 0.08 ab | 1.63 ± 0.43 a | 2.29 ± 0.65 a |
| reference varieties | 4 | 191.38 ± 12.72 ab | 19.95 ± 1.21 a | 11.72 ± 4.36 ab | 1.03 ± 0.14 b | 1.12 ± 0.37 a | 3.27 ± 0.57 a | 0.40 ± 0.15 a | 1.63 ± 0.53 a | 2.09 ± 0.87 a |
| fruit varieties | 4 | 183.95 ± 24.20 b | 20.07 ± 2.03 a | 13.43 ± 3.61 a | 2.47 ± 1.64 a | 1.53 ± 1.07 a | 1.63 ± 0.40 c | 0.41 ± 0.26 a | 1.26 ± 0.24 b | 2.35 ± 1.31 a |
| n | P | S | K | Ca | Cl | ||
|---|---|---|---|---|---|---|---|
| Slovenian genotypes | 19 | 1.75 ± 0.75 | 0.88 ± 0.23 a | 15.92 ± 3.59 | 17.14 ± 4.90 | 788.16 ± 36.26 | |
| Hungarian genotypes | 25 | 1.42 ± 0.32 | 0.83 ± 0.19 ab | 16.47 ± 3.16 | 14.03 ± 3.47 | 953.20 ± 34.69 | |
| reference varieties | 4 | 1.52 ± 0.23 | 0.84 ± 0.80 ab | 16.15 ± 4.39 | 14.63 ± 2.57 | 735.25 ± 27.79 | |
| fruit varieties | 4 | 1.30 ± 0.18 | 0.62 ± 0.12 b | 17.58 ± 1.48 | 14.20 ± 2.72 | 569.25 ± 18.45 | |
| Mn | Fe | Ni | Zn | Rb | Sr | ||
| Slovenian genotypes | 19 | 66.04 ± 14.58 a | 147.42 ± 26.27 ab | 28.35 ± 11.88 | 23.91 ± 11.34 | 36.23 ± 28.72 | 54.02 ± 25.16 ab |
| Hungarian genotypes | 25 | 67.32 ± 21.29 a | 154.36 ± 29.89 a | 24.19 ± 9.40 | 19.98 ± 5.66 | 26.75 ± 9.73 | 72.01 ± 31.40 a |
| reference varieties | 4 | 52.03 ± 6.16 ab | 128.25 ± 21.42 ab | 21.68 ± 8.06 | 18.20 ± 5.32 | 31.20 ± 20.27 | 35.10 ± 7.16 b |
| fruit varieties | 4 | 45.38 ± 4.22 b | 119.50 ± 7.05 b | 23.65 ± 4.20 | 18.40 ± 4.59 | 22.40 ± 7.73 | 57.53 ± 15.07 ab |
| n | 5th Instar/5th D | 5th Instar/6th D | 5th Instar/7th D | Cocoon FW | |
|---|---|---|---|---|---|
| Slovenian mulberry genotypes | 19 | 3.97 ± 0.46 b | 4.28 ± 0.33 b | 4.77 ± 0.46 b | 2.14 ± 0.12 a |
| Hungarian mulberry genotypes | 25 | 4.32 ± 0.46 ab | 4.46 ± 0.34 b | 4.96 ± 0.43 b | 2.26 ± 0.09 a |
| reference sericultural varieties | 4 | 4.54 ± 0.38 a | 4.85 ± 0.23 a | 5.36 ± 0.36 a | 2.25 ± 0.14 a |
| fruit varieties | 4 | 4.39 ± 0.57 ab | 4.55 ± 0.33 ab | 5.07 ± 0.30 ab | 2.20 ± 0.14 a |
| n | Silk Thread Parameters | Reeling Wastes | ||||
|---|---|---|---|---|---|---|
| Length | Weight | Thickness | Struse | Telette | ||
| Slovenian genotypes | 95 | 1419.09 ± 64.14 a | 0.43 ± 0.03 a | 2.72 ± 0.13 a | 0.036 ± 0.013 b | 0.017 ± 0.009 a |
| Hungarian genotypes | 211 | 1429.18 ± 94.82 a | 0.44 ± 0.03 a | 2.82 ± 0.15 a | 0.047 ± 0.022 a | 0.021 ± 0.019 a |
| reference varieties | 70 | 1363.20 ± 135.94 a | 0.43 ± 0.02 a | 2.84 ± 0.19 a | 0.043 ± 0.016 ab | 0.017 ± 0.016 a |
| fruit varieties | 30 | 1244.80 ± 44.03 b | 0.37 ± 0.03 b | 2.73 ± 0.13 a | 0.040 ± 0.020 b | 0.022 ± 0.016 a |
| Silkworm Weight (Larvae/g) | Cocoon Weight | Silk Thread | Reeling Waste | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Correlations | 5th C/5th D | 6th D | 7th D | Mean 3d Weight | Fresh | Dry | Length | Weight | Thickness | Struse | Telette |
| 5th C/5th D | 1 | 00.856** | 0.764 ** | 0.934 ** | 0.777 ** | 0.766 ** | −0.11 | 0.191 | 0.438 * | 0.234 | 0.121 |
| 5th C/6th D | 0.856 ** | 1 | 0.907 ** | 0.967 ** | 0.713 ** | 0.715 ** | −0.088 | 0.21 | 0.452 ** | 0.18 | 0.066 |
| 5th C/7th D | 0.764 ** | 0.907 ** | 1 | 0.935 ** | 0.607 ** | 0.618 ** | −0.032 | 0.166 | 0.298 | 0.226 | 0.133 |
| mean 3d weight | 0.934 ** | 0.967 ** | 0.935 ** | 1 | 0.744 ** | 0.745 ** | −0.082 | 0.199 | 0.418 * | 0.229 | 0.116 |
| cocoon FW | 0.777 ** | 0.713 ** | 0.607 ** | 0.744 ** | 1 | 0.992 ** | 0.024 | 0.376 * | 0.562 ** | 0.193 | 0.081 |
| cocoon DW | 0.766 ** | 0.715 ** | 0.618 ** | 0.745 ** | 0.992 ** | 1 | 0.016 | 0.378 * | 0.583 ** | 0.189 | 0.076 |
| length | −0.11 | −0.088 | −0.032 | −0.082 | 0.024 | 0.016 | 1 | 0.785 ** | −0.075 | −00.368 * | −0.183 |
| weight | 0.191 | 0.21 | 0.166 | 0.199 | 0.376 * | 0.378 * | 0.785 ** | 1 | 0.555 ** | −00.455 ** | −0.07 |
| thickness | 0.438 * | 0.452 ** | 0.298 | 0.418 * | 0.562 ** | 0.583 ** | −0.075 | 0.555 ** | 1 | −0.269 | 0.099 |
| struse | 0.234 | 0.18 | 0.226 | 0.229 | 0.193 | 0.189 | −0.368 * | −0.455 ** | −0.269 | 1 | 0.176 |
| telette | 0.121 | 0.066 | 0.133 | 0.116 | 0.081 | 0.076 | −0.183 | −0.07 | 0.099 | 0.176 | 1 |
| total proteins | −0.198 | −0.139 | −0.116 | −0.164 | 0.083 | 0.073 | 0.448 ** | 0.495 ** | 0.175 | −0.024 | −0.262 |
| total phenolics | −0.001 | 0.131 | 0.137 | 0.087 | 0.096 | 0.108 | −0.197 | −0.12 | 0.069 | 0.191 | 0.13 |
| chlorogenic a. | 0.02 | 0.202 | 0.228 | 0.148 | 0.205 | 0.205 | −0.198 | −0.068 | 0.165 | 0.211 | 0.15 |
| 4-caffeoyl-QA | −0.092 | −0.12 | 0.006 | −0.071 | −0.125 | −0.136 | −0.329 | −0.312 | −0.098 | 0.211 | 0.246 |
| 5-caffeoyl-QA | −0.031 | −0.068 | −0.002 | −0.034 | 0.004 | 0.019 | 0.109 | 0.079 | −0.023 | 0.046 | −0.15 |
| total caffeoyl-QA | −0.003 | 0.154 | 0.206 | 0.117 | 0.157 | 0.155 | −0.248 | −0.127 | 0.126 | 0.236 | 0.186 |
| c-5-CQA | 0.023 | 0.034 | 0.037 | 0.033 | 0.231 | 0.225 | 0.21 | 0.272 | 0.176 | 0.034 | −0.167 |
| t-5-CQA | −0.451 ** | −0.328 | −0.281 | −0.382 | −0.265 | −0.28 | −0.01 | 0.002 | 0.003 | −0.1 | −0.092 |
| p-CAH | −0.048 | −0.15 | −0.059 | −0.084 | −0.123 | −0.13 | −0.378 * | −0.322 | −0.045 | 0.158 | 0.196 |
| p-CAH2 | −0.044 | −0.057 | −0.1 | −0.07 | 0.028 | 0.008 | 0.15 | 0.168 | 0.066 | 0.05 | −0.281 |
| p-CQA | −0.147 | −0.181 | −0.154 | −0.168 | −0.094 | −0.09 | −0.283 | −0.313 | −0.097 | 0.121 | −0.034 |
| total coumaroyl-QA | −0.183 | −0.171 | −0.146 | −0.177 | −0.04 | −0.055 | 0.007 | 0.05 | 0.065 | 0.044 | −0.145 |
| rutin | −0.179 | −0.054 | −0.05 | −0.108 | 0.081 | 0.101 | −0.02 | −0.041 | −0.033 | 0.364 * | −0.018 |
| Q-3-glu | −0.326 | −0.105 | −0.056 | −0.185 | −0.163 | −0.153 | −0.247 | −0.342 | −0.225 | 0.298 | 0.033 |
| QMH | −0.268 | 0.02 | −0.047 | −0.122 | 0.098 | 0.11 | 0.085 | 0.177 | 0.193 | −0.029 | −0.161 |
| Q-diR-gly | −0.082 | −0.098 | −0.132 | −0.109 | −0.155 | −0.128 | −0.209 | −0.228 | −0.088 | 0.086 | 0.011 |
| QRH | −0.299 | −0.301 | −0.235 | −0.295 | −0.25 | −0.27 | −0.16 | −0.304 | −0.303 | 0.127 | 0.155 |
| Q-acetyl-RH | −0.207 | −0.185 | −0.084 | −0.168 | −0.044 | −0.065 | −0.082 | −0.125 | −0.1 | 0.046 | −0.047 |
| QAH | 0.009 | 0.135 | 0.006 | 0.045 | 0.284 | 0.275 | −0.132 | 0.054 | 0.264 | 0.189 | −0.12 |
| total quercetin-gly | −0.27 | −0.044 | −0.071 | −0.15 | 0.077 | 0.097 | −0.037 | −0.018 | 0.033 | 0.298 | −0.077 |
| KAH | −0.197 | −0.071 | −0.007 | −0.105 | −0.043 | −0.044 | −0.077 | −0.028 | 0.066 | 0.086 | 0.054 |
| K-diRH | 0.126 | 0.054 | −0.044 | 0.052 | 0.05 | 0.01 | −0.525 ** | −0.407 * | 0.042 | 0.284 | 0.231 |
| KRH | −0.199 | −0.175 | −0.075 | −0.16 | 0.003 | 0.012 | −0.152 | −0.204 | −0.125 | 0.482 ** | 0.106 |
| K-acetyl-RH | −0.192 | −0.175 | −0.091 | −0.163 | −0.216 | −0.23 | −0.247 | −0.324 | −0.22 | 0.163 | 0.167 |
| total K-gly. deriv. * | −0.194 | −0.101 | −0.038 | −0.123 | −0.046 | −0.053 | −0.202 | −0.162 | 0.011 | 0.231 | 0.118 |
| Silkworm Weight (Larvae/g) | Cocoon Weight | Silk Thread Parameters | Reeling Waste | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Correlations | 5th C/5th D | 5th C/6th D | 5th C/7th D | 3d Weight | Fresh | Dry | Length | Weight | Thickness | Struse | Telette |
| 5th C/5th D | 1 | 0.856 ** | 0.764 ** | 0.934 ** | 0.777 ** | 0.766 ** | −0.11 | 0.191 | 0.438 * | 0.234 | 0.121 |
| 5th C/6th D | 0.856 ** | 1 | 0.907 ** | 0.967 ** | 0.713 ** | 0.715 ** | −0.088 | 0.21 | 0.452 ** | 0.18 | 0.066 |
| 5th C/7th D | 0.764 ** | 0.907 ** | 1 | 0.935 ** | 0.607 ** | 0.618 ** | −0.032 | 0.166 | 0.298 | 0.226 | 0.133 |
| mean 3d weight | 0.934 ** | 0.967 ** | 0.935 ** | 1 | 0.744 ** | 0.745 ** | −0.082 | 0.199 | 0.418 * | 0.229 | 0.116 |
| cocoon FW | 0.777 ** | 0.713 ** | 0.607 ** | 0.744 ** | 1 | 0.992 ** | 0.024 | 0.376 * | 0.562 ** | 0.193 | 0.081 |
| cocoon DW | 0.766 ** | 0.715 ** | 0.618 ** | 0.745 ** | 0.992 ** | 1 | 0.016 | 0.378 * | 0.583 ** | 0.189 | 0.076 |
| length | −0.11 | −0.088 | −0.032 | −0.082 | 0.024 | 0.016 | 1 | 0.785 ** | −0.075 | −0.368 * | −0.183 |
| weight | 0.191 | 0.21 | 0.166 | 0.199 | 0.376 * | 0.378 * | 0.785 ** | 1 | 0.555 ** | −0.455 ** | −0.07 |
| thickness | 0.438 * | 0.452 ** | 0.298 | 0.418 * | 0.562 ** | 0.583 ** | −0.075 | 0.555 ** | 1 | −0.269 | 0.099 |
| struse | 0.234 | 0.18 | 0.226 | 0.229 | 0.193 | 0.189 | −0.368 * | −0.455 ** | −0.269 | 1 | 0.176 |
| telette | 0.121 | 0.066 | 0.133 | 0.116 | 0.081 | 0.076 | −0.183 | −0.07 | 0.099 | 0.176 | 1 |
| P | −0.016 | 0.001 | −0.001 | −0.006 | 0.144 | 0.129 | 0.269 | 0.261 | 0.052 | −0.068 | −0.097 |
| S | 0.207 | 0.212 | 0.347 | 0.271 | 0.075 | 0.08 | 0.228 | 0.287 | 0.135 | −0.017 | 0.331 |
| K | 0.288 | 0.236 | 0.208 | 0.262 | 0.282 | 0.296 | 0.178 | 0.234 | 0.176 | −0.14 | −0.155 |
| Ca | 0.074 | −0.108 | −0.086 | −0.032 | −0.103 | −0.141 | −0.16 | −0.1 | 0.021 | −0.108 | 0.166 |
| Cl | −0.003 | −0.136 | −0.194 | −0.11 | 0.042 | 0.063 | 0.237 | 0.352 * | 0.249 | −0.211 | −0.022 |
| Mn | −0.058 | −0.152 | −0.05 | −0.086 | −0.035 | −0.041 | 0.334 | 0.324 | 0.058 | −0.106 | 0.226 |
| Fe | −0.074 | −0.115 | −0.062 | −0.086 | −0.184 | −0.202 | 0.167 | 0.113 | −0.062 | −0.019 | 0.434 * |
| Ni | −0.035 | −0.027 | −0.049 | −0.04 | −0.161 | −0.211 | 0.027 | 0.012 | −0.02 | −0.151 | 0.185 |
| Zn | −0.052 | −0.048 | 0.025 | −0.027 | −0.139 | −0.126 | 0.081 | 0.037 | −0.032 | −0.105 | −0.016 |
| Rb | −0.367 | −0.094 | 0.056 | −0.159 | −0.276 | −0.267 | 0.26 | 0.097 | −0.185 | −0.119 | −0.16 |
| Sr | −0.083 | −0.205 | −0.168 | −0.154 | −0.083 | −0.123 | −0.003 | 0.005 | −0.024 | −0.042 | 0.533 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanek Krajnc, A.; Bakonyi, T.; Ando, I.; Kurucz, E.; Solymosi, N.; Pongrac, P.; Berčič, R.L. The Effect of Feeding with Central European Local Mulberry Genotypes on the Development and Health Status of Silkworms and Quality Parameters of Raw Silk. Insects 2022, 13, 836. https://doi.org/10.3390/insects13090836
Urbanek Krajnc A, Bakonyi T, Ando I, Kurucz E, Solymosi N, Pongrac P, Berčič RL. The Effect of Feeding with Central European Local Mulberry Genotypes on the Development and Health Status of Silkworms and Quality Parameters of Raw Silk. Insects. 2022; 13(9):836. https://doi.org/10.3390/insects13090836
Chicago/Turabian StyleUrbanek Krajnc, Andreja, Tamas Bakonyi, Istvan Ando, Eva Kurucz, Norbert Solymosi, Paula Pongrac, and Rebeka Lucijana Berčič. 2022. "The Effect of Feeding with Central European Local Mulberry Genotypes on the Development and Health Status of Silkworms and Quality Parameters of Raw Silk" Insects 13, no. 9: 836. https://doi.org/10.3390/insects13090836
APA StyleUrbanek Krajnc, A., Bakonyi, T., Ando, I., Kurucz, E., Solymosi, N., Pongrac, P., & Berčič, R. L. (2022). The Effect of Feeding with Central European Local Mulberry Genotypes on the Development and Health Status of Silkworms and Quality Parameters of Raw Silk. Insects, 13(9), 836. https://doi.org/10.3390/insects13090836