
Genome-Wide Identification and Analysis of the Heat-Shock Protein Gene Superfamily in Bemisia tabaci and Expression Pattern Analysis under Heat Shock



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing Conditions
2.2. Identification of HSP Genes from Genome Database
2.3. Classification and Nomenclature of HSP Genes
2.4. Structural Information Prediction and Analysis
2.5. Heat-Stress Treatments
2.6. Quantitative Reverse Transcription PCR (RT-qPCR)
2.7. RNA Interference and Survival Rate Analysis
2.8. Data Statistical Analysis
3. Results
3.1. Identification of HSP Genes in the Genome of B. tabaci MED
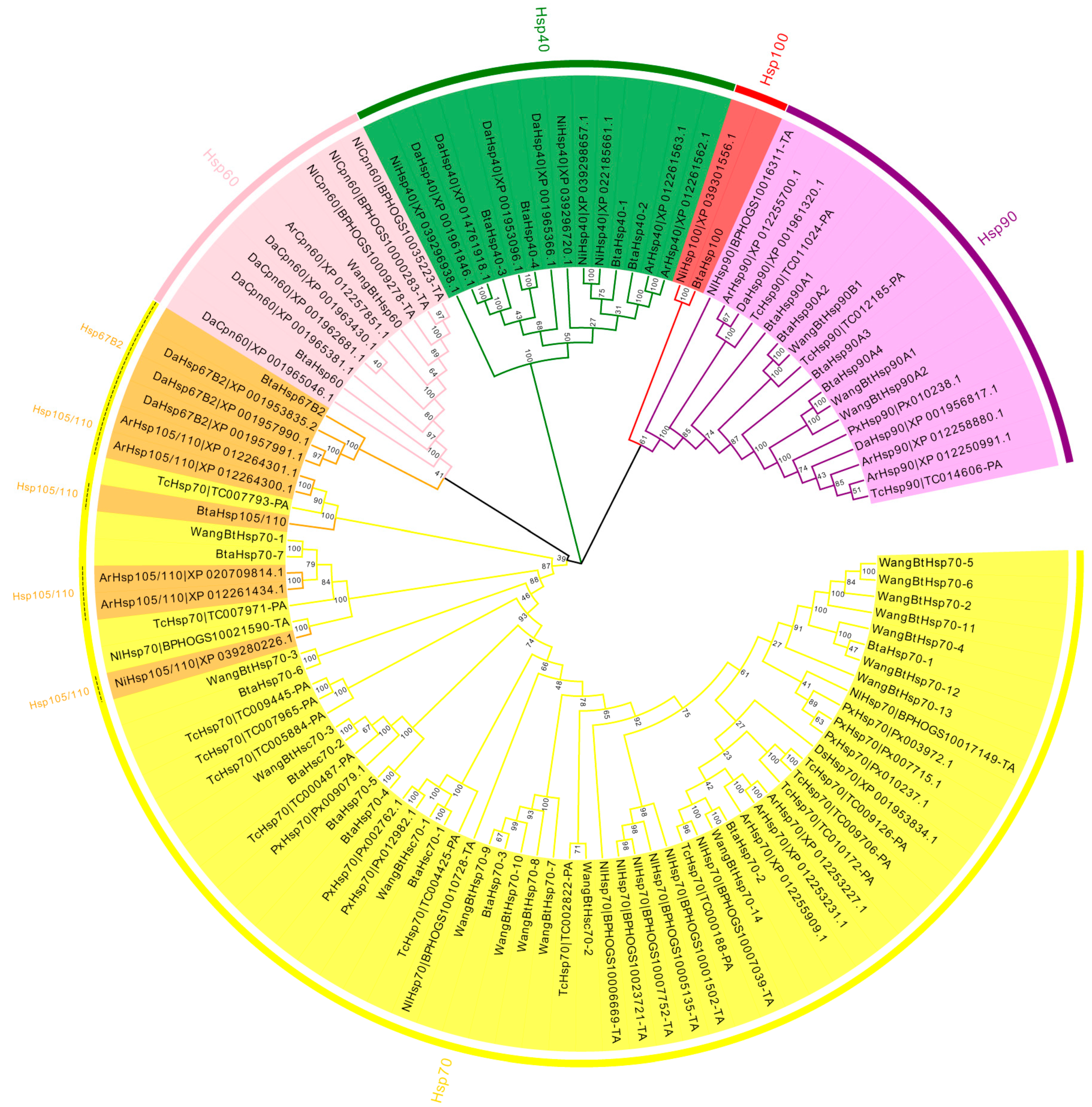
3.2. Phylogenetic Analysis for Classification Validation
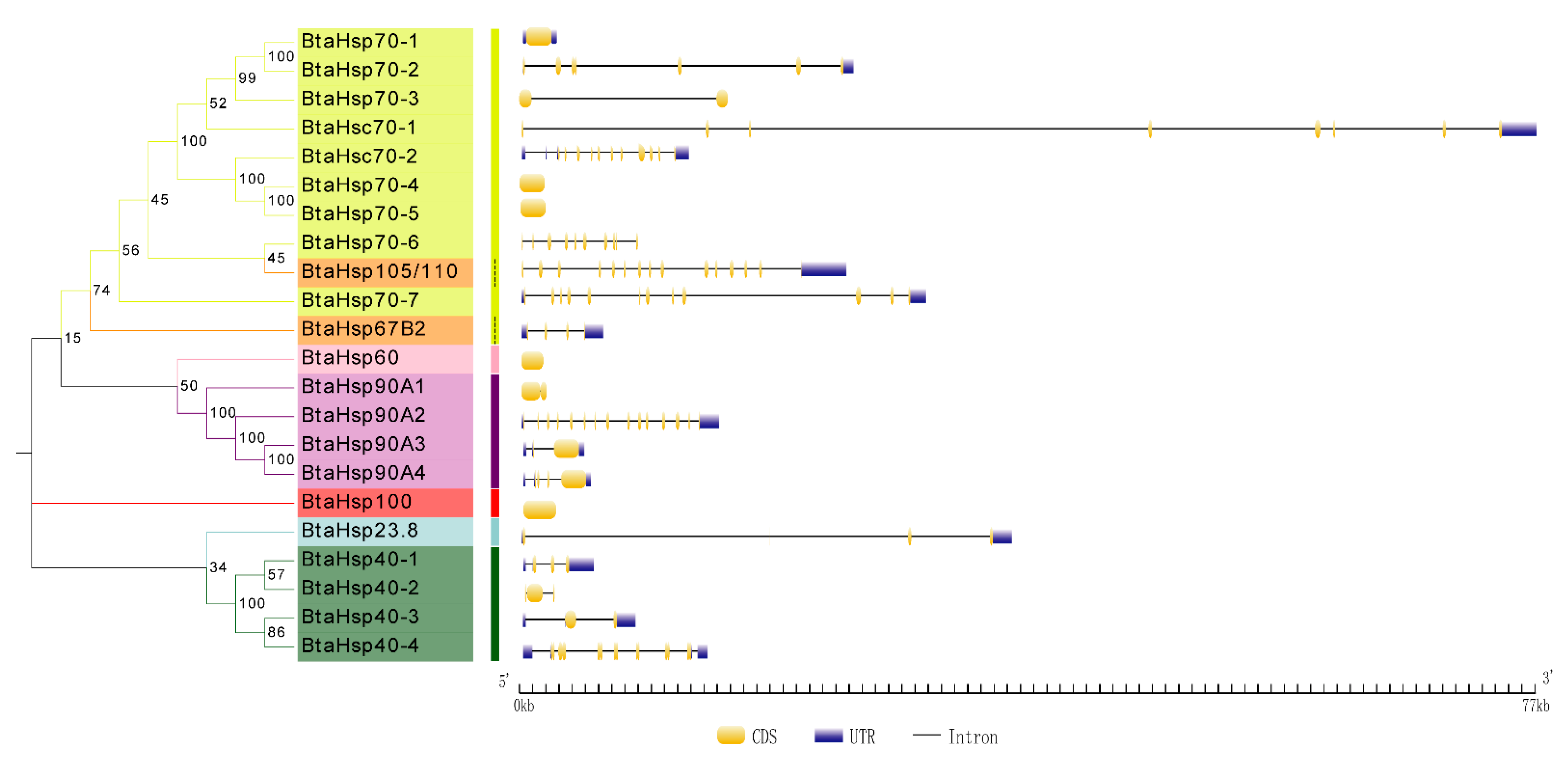
3.3. Phylogenetic and Gene Structure Analysis of BtaHSP Genes
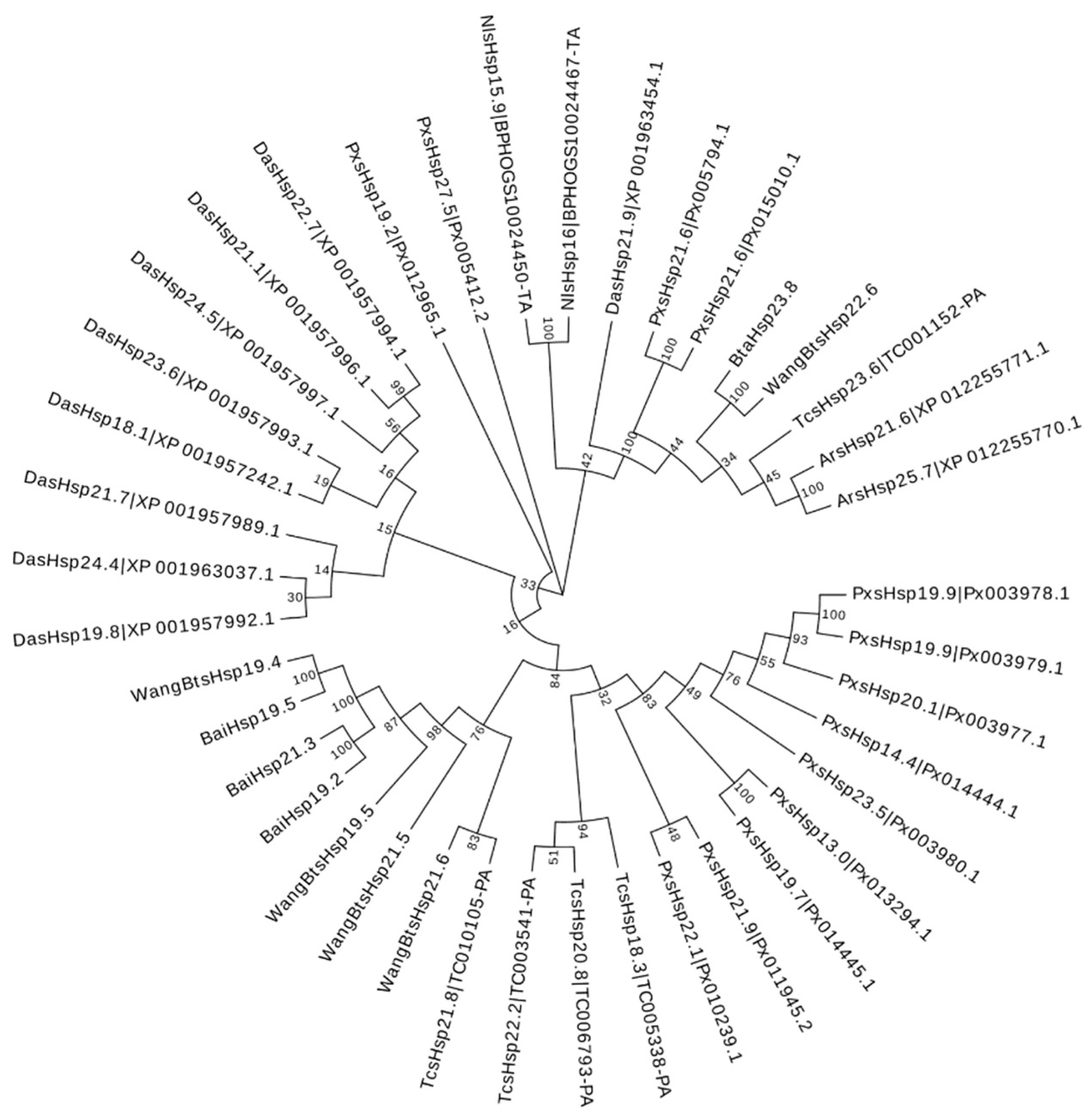
3.4. Phylogenetic and Gene Secondary Structure Analysis of BtaHSP Genes
3.5. Elevated Differential Expression of BtaHSP Genes at High Temperature
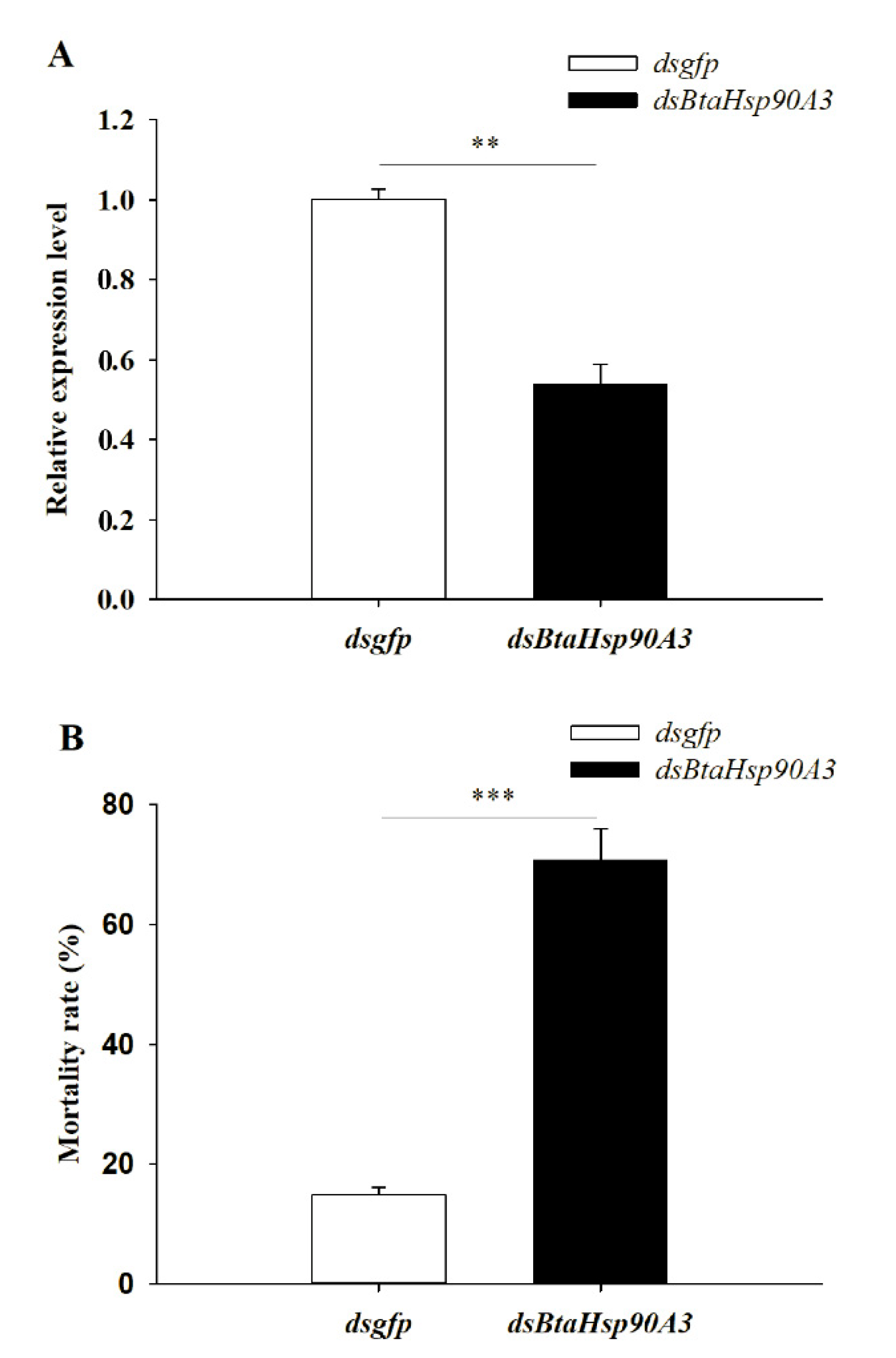
3.6. Effect of HSP Gene on High-Temperature Stress
4. Discussion
4.1. Expansion of HSP Gene Superfamily in B. tabaci
4.2. Conserved-Sequence Characteristics of Members of the B. tabaci HSP Superfamily
4.3. Specific BtaHSP Genes Are Important in Coping with Temperature Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boher, F.; Trefault, N.; Piulachs, M.D.; Belles, X.; Godoy, H.R.; Bozinovic, F. Biogeographic origin and thermal acclimation interact to determine survival and hsp90 expression in Drosophila species submitted to thermal stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 162, 391–396. [Google Scholar] [CrossRef]
- Stetina, T.; Kostal, V.; Korbelova, J. The role of inducible Hsp70, and other heat shock proteins, in adaptive complex of cold tolerance of the fruit fly (Drosophila melanogaster). PLoS ONE 2015, 10, e0128976. [Google Scholar] [CrossRef] [Green Version]
- Teets, N.M.; Peyton, J.T.; Colinet, H.; Renault, D.; Denlinger, D.L. Gene expression changes governing extreme dehydration tolerance in an antarctic insect. Proc. Natl. Acad. Sci. USA 2013, 109, 20744–20749. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.L.; Cheng, H.; Cheng, S.Y.; Li, L.L.; Xu, F.; Yu, W.W.; Yuan, H.H. Expression of selected Ginkgo biloba heat shock protein genes after cold treatment could be induced by other abiotic stress. Int. J. Mol. Sci. 2012, 13, 5768–5788. [Google Scholar] [CrossRef]
- Brigotti, M.; Petronini, P.G.; Carnicelli, D.; Alfieri, R.R.; Bonelli, M.A.; Borghetti, A.F.; Wheeler, K.P. Effects of osmolarity, ions and compatible osmolytes on cell−free protein synthesis. Biochem. J. 2003, 369, 369–374. [Google Scholar] [CrossRef]
- Xin, L.L.; Li, X.H.; Deng, H.X.; Kuang, D.; Dai, X.Y.; Huang, S.L.; Wang, F.; He, M.A.; Currie, R.W.; Wu, T.C. Development of stable HSPA1A promoter−driven luciferase reporter HepG2 cells for assessing the toxicity of organic pollutants present in air. Cell Stress Chaperones 2012, 17, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.D.; Du, Y.Z.; Lu, M.X.; Qiang, C.K. Cloning of the heat shock protein 60 gene from the stem borer, Chilo suppressalis, and analysis of expression characteristics under heat stress. J. Insect Sci. 2010, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Advani, N.K.; Kenkel, C.D.; Davies, S.W.; Parmesan, C.; Singer, M.C.; Matz, M.V. Variation in heat shock protein expression at the latitudinal range limits of a widely−distributed species, the Glanville fritillary butterfly (Melitaea cinxia). Physiol. Entomol. 2016, 41, 241–248. [Google Scholar] [CrossRef]
- Cahan, S.H.; Nguyen, A.D.; Stanton, G.J.; Penick, C.A.; Hernaiz, H.Y.; de Marco, B.B.; Gotelli, N.J. Modulation of the heat shock response is associated with acclimation to novel temperatures but not adaptation to climatic variation in the ants Aphaenogaster picea and A. rudis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 204, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.H.; Du, Y.; Wang, J.W. Molecular characterization and expression patterns of Spodoptera litura heat shock protein 70/90, and their response to zinc stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 158, 102–110. [Google Scholar] [CrossRef]
- Kim, B.M.; Rhee, J.S.; Jeong, C.B.; Seo, J.S.; Park, G.S.; Lee, Y.M.; Lee, J.S. Heavy metals induce oxidative stress and trigger oxidative stress−mediated heat shock protein (hsp) modulation in the intertidal copepod Tigriopus japonicus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 166, 65–74. [Google Scholar] [CrossRef]
- Ashbrook, A.R.; Feder, J.L.; Scharf, M.E.; Bennett, G.W.; Gondhalekar, A.D. Characterization of heat exposure−associated escape behaviors and HSP gene expression in bed bugs (Cimex lectularius L.). Pest Manag. Sci. 2021, 78, 205–216. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat−shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Ann. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, C.A.; Kabaalu, R.; Visendi, P.; Sseruwagi, P.; Seal, S.E. Genetic diversity of whitefly (Bemisia spp.) on crop and uncultivated plants in Uganda: Implications for the control of this devastating pest species complex in Africa. J. Pest Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Guo, C.L.; Zhu, Y.Z.; Zhang, Y.J.; Keller, M.A.; Liu, T.X.; Chu, D. Invasion biology and management of sweetpotato whitefly (Hemiptera: Aleyrodidae) in China. J. Integr. Pest Manag. 2021, 12, pmaa024. [Google Scholar] [CrossRef]
- Díaz, F.; Orobio, R.F.; Chavarriaga, P.; Toro−Perea, N. Differential expression patterns among heat−shock protein genes and thermal responses in the whitefly Bemisia tabaci (MEAM1). J. Therm. Biol. 2015, 52, 199–207. [Google Scholar] [CrossRef]
- Su, Q.; Li, S.; Shi, C.; Zhang, J.; Zhang, G.; Jin, Z.; Zhang, Y. Implication of heat−shock protein 70 and UDP−glucuronosyltransferase in thiamethoxam−induced whitefly Bemisia tabaci thermotolerance. J. Pest Sci. 2018, 91, 469–478. [Google Scholar] [CrossRef]
- Bai, J.; Wang, Y.C.; Liu, Y.C.; Chang, Y.W.; Liu, X.N.; Gong, W.R.; Du, Y.-Z. Isolation of two new genes encoding heat shock protein 70 in Bemisia tabaci and analysis during thermal stress. Int. J. Biol. Macromol. 2021, 193, 933–940. [Google Scholar] [CrossRef]
- Xiao, N.; Pan, L.L.; Zhang, C.R.; Shan, H.W.; Liu, S.S. Differential tolerance capacity to unfavourable low and high temperatures between two invasive whiteflies. Sci. Rep. 2016, 6, 24306. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Wan, F.H.; Guo, J.Y. Different thermal tolerance and hsp gene expression in invasive and indigenous sibling species of Bemisia tabaci. Biol. Invasions 2012, 14, 1587–1595. [Google Scholar] [CrossRef]
- Pusag, J.C.A.; Jahan, S.M.H.; Lee, K.S.; Lee, S.; Lee, K.Y. Upregulation of temperature susceptibility in Bemisia tabaci upon acquisition of tomato yellow leaf curl virus (TYLCV). J. Insect Physiol. 2012, 58, 1343–1348. [Google Scholar] [CrossRef]
- Wang, X.R.; Wang, C.; Ban, F.X.; Zhu, D.T.; Liu, S.S.; Wang, X.W. Genome−wide identification and characterization of HSP gene superfamily in whitefly (Bemisia tabaci) and expression profiling analysis under temperature stress. Insect Sci. 2019, 26, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Liu, X.N.; Lu, M.X.; Du, Y.Z. Characterization of genes encoding small heat shock proteins from Bemisia tabaci and expression under thermal stress. PeerJ 2019, 7, e6992. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.; Hu, X.; Gao, C.; Zhao, H.; Nichols, R.L.; Li, X. Use of mitochondrial cytochrome oxidase I polymerase chain reaction−restriction fragment length polymorphism for identifying subclades of Bemisia tabaci Mediterranean group. J. Econ. Entomol. 2012, 105, 242–251. [Google Scholar] [CrossRef]
- Marchler−Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Tian, L.; Xie, J.T.; Huang, Q.Y.; Feng, M.G.; Keyhani, N.O. A fungal sirtuin modulates development and virulence in the insect pathogen, Beauveria bassiana. Environ. Microbiol. 2021, 23, 5164–5183. [Google Scholar] [CrossRef]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for Gram−negative bacteria by support vector machines based on n−peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, P.; Ingham, V.A.; Balabanidou, V.; Vontas, J.; Lycett, G.; Ranson, H. The Anopheles gambiae ATP−binding cassette transporter family: Phylogenetic analysis and tissue localization provide clues on function and role in insecticide resistance. Insect Mol. Biol. 2018, 27, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, S.; Craig, E.A. The heat−shock proteins. Ann. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Li, Z.W.; Li, X.; Yu, Q.Y.; Xiang, Z.H.; Kishino, H.; Zhang, Z. The small heat shock protein (sHSP) genes in the silkworm, Bombyx mori, and comparative analysis with other insect sHSP genes. BMC Evol. Biol. 2009, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kiang, J.G.; Tsokos, G.C. Heat shock protein 70 kDa: Molecular biology, biochemistry, and physiology. Pharmacol. Ther. 1998, 80, 183–201. [Google Scholar] [CrossRef]
- Chen, B.; Zhong, D.; Monteiro, A. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genom. 2006, 7, 596–609. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.Y.H.; Yue, H.; Huang, L.P.; Zhang, D.Y.; Zhang, Z.H.; Zhang, Z.; Zhang, Y.J.; Li, F.; Yan, F.; Zhou, X.G.; et al. Suppression of Bta11975, an α-glucosidase, by RNA interference reduces transmission of tomato chlorosis virus by Bemisia tabaci. Pest Manag. Sci. 2021, 77, 5294–5303. [Google Scholar] [CrossRef]
- Gharbi, S.; Shamsara, M.; Khateri, S.; Soroush, M.R.; Mowla, S.J. Identification of reliable reference genes for quantification of microRNAs in serum samples of sulfur mustard−exposed veterans. Cell J. 2015, 17, 494–501. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real−time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- Nobuyuki, Y.; Hajime, F.; Youhei, S.; Takumi, H. Hsp105β upregulates hsp70 gene expression through signal transducer and activator of transcription−3. FEBS J. 2009, 276, 5870–5880. [Google Scholar] [CrossRef]
- Hartl, F.U. Molecular chaperones in cellular protein folding. Nature 1996, 381, 571–580. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome−wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Guo, B.; He, S. The roles and evolutionary patterns of intronless genes in deuterostomes. Compar. Funct. Genom. 2011, 252–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Boorstein, W.R.; Ziegelhoffer, T.; Craig, E.A. Molecular evolution of the Hsp70 multigene family. J. Mol. Biol. 1994, 38, 1–17. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Kluck, C.J.; Patzelt, H.; Genevaux, P.; Brehmer, D.; Rist, W.; Schneider−Mergener, J.; Bukau, B.; Mayer, M.P. Structure−function analysis of HscC, the Escherichia coli member of a novel subfamily of specialized Hsp70 chaperones. J. Biol. Chem. 2002, 277, 41060–41069. [Google Scholar] [CrossRef] [Green Version]
- Walsh, P.; Bursa, D.; Law, Y.C.; Cyr, D.; Lithgow, T. The J−protein family: Modulating protein assembly, disassembly and translocation. EMBO Rep. 2004, 5, 567–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritha, H.; Michael, E.C.; Heini, W.D.; Gregory, L.B. Analysis of the levels of conservation of the J domain among the various types of DnaJ−like proteins. Cell Stress Chaperones 2000, 5, 347. [Google Scholar] [CrossRef]
- Jiang, X.; Zhai, H.; Wang, L.; Luo, L.; Zhang, S.L. Cloning of the heat shock protein 90 and 70 genes from the beet armyworm, Spodoptera exigua, and expression characteristics in relation to thermal stress and development. Cell Stress Chaperones 2012, 17, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. Recent advances in understanding catalysis of protein folding by molecular chaperones. FEBS Lett. 2020, 594, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Cao, Y.; Xu, Y.T.; Huai, D.; Chen, P.; Guo, J.; Li, M.; Dai, Y.Y. Overexpression of Hsc70 promotes proliferation, migration, and invasion of human glioma cells. J. Cell. Biochem. 2019, 120, 10707–10714. [Google Scholar] [CrossRef]
- Shim, J.K.; Lee, G.S.; Lee, S.; Lee, K.Y. Oral ingestion of heat shock protein 70 dsRNA is lethal under normal and thermal stress conditions in the sweetpotato whitefly, Bemisia tabaci. J. Asia Pac. Entomol. 2015, 18, 797–800. [Google Scholar] [CrossRef]
- Jiang, F.; Chang, G.; Li, Z.; Abouzaid, M.; Du, X.; Lin, Y. The HSP/co−chaperone network in environmental cold adaptation of Chilo suppressalis. Int. J. Biol. Macromol. 2021, 187, 780–788. [Google Scholar] [CrossRef]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef]
- Tania, M.L.; Matthias, P.M.; Stefan, G.R. The Hsp70−Hsp90 chaperone cascade in protein folding. Trends Cell Biol. 2019, 29, 164–177. [Google Scholar] [CrossRef]
- Karagoez, G.E.; Rudiger, S.G.D. Hsp90 interaction with clients. Trends Biochem. Sci. 2015, 40, 117–125. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Gene Identifier | Gene Name | CDS | Mw (kDa) | pI | Subcelluar Location | Strand |
|---|---|---|---|---|---|---|---|
| sHSP | OM176688 | BtaHsp23.8 | 91–720 | 23.8 | 6.92 | Extracellular/nuclear | Plus |
| HSP40 | OM176682 | BtaHsp40-1 | 202–957 | 28.9 | 6.55 | Nuclear | Plus |
| OM176693 | BtaHsp40-2 | 1–1350 | 50.2 | 9.31 | Nuclear | Plus | |
| OM176684 | BtaHsp40-3 | 232–1284 | 38.8 | 9.11 | Nuclear | Plus | |
| OM176675 | BtaHsp40-4 | 380–1579 | 44.4 | 6.35 | Nuclear | Plus | |
| HSP60 | OM176680 | BtaHsp60 | 1–1668 | 58.9 | 5.38 | Cytoplasmic/nuclear | Plus |
| HSP70 | OM176673 | BtaHsp70-1 | 228–2180 | 72 | 8.22 | Mitochondrial/nuclear | Plus |
| OM176691 | BtaHsp70-2 | 7–1659 | 60.7 | 5.6 | Cytoplasmic/nuclear | Plus | |
| OM176686 | BtaHsp70-3 | 1–1776 | 65.7 | 5.92 | Cytoplasmic/nuclear | Plus | |
| OM176692 | BtaHsc70-1 | 1–1758 | 65.3 | 5.21 | ER/nuclear | Plus | |
| OM176674 | BtaHsc70-2 | 394–2253 | 67.5 | 5.3 | Nuclear | Plus | |
| OM176678 | BtaHsp70-4 | 1–1908 | 68.7 | 4.94 | Cytoplasmic/nuclear | Plus | |
| OM176681 | BtaHsp70-5 | 1–1926 | 70.7 | 5.5 | Cytoplasmic/nuclear | Plus | |
| OM176685 | BtaHsp70-6 | 1–1572 | 57.6 | 5.57 | Plasma membrane | Plus | |
| OM176676 | BtaHsp70-7 | 207–2618 | 89.9 | 5.43 | Cytoplasmic/nuclear | Plus | |
| OM176689 | BtaHsp105/110 | 1–2904 | 107.6 | 5.32 | Cytoplasmic/ER | Plus | |
| OM176683 | BtaHsp67B2 | 420–917 | 18.6 | 8.91 | Mitochondrial | Plus | |
| HSP90 | OM176677 | BtaHsp90A1 | 1–1902 | 72.4 | 5.11 | Cytoplasmic/nuclear | Plus |
| OM176694 | BtaHsp90A2 | 135–2462 | 88.7 | 5.09 | Cytoplasmic/cytoskeletal/nuclear | Plus | |
| OM176687 | BtaHsp90A3 | 308–2242 | 74.2 | 5.07 | Cytoplasmic/nuclear | Plus | |
| OM176690 | BtaHsp90A4 | 207–2402 | 84 | 5.01 | Cytoplasmic/nuclear | Plus | |
| HSP100 | OM176679 | BtaHsp100 | 1–2472 | 92.4 | 5.96 | Cytoplasmic | Plus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.-Y.; Qin, P.-H.; Yang, K.; Liu, T.-X.; Zhang, Y.-J.; Chu, D. Genome-Wide Identification and Analysis of the Heat-Shock Protein Gene Superfamily in Bemisia tabaci and Expression Pattern Analysis under Heat Shock. Insects 2022, 13, 570. https://doi.org/10.3390/insects13070570
Zheng H-Y, Qin P-H, Yang K, Liu T-X, Zhang Y-J, Chu D. Genome-Wide Identification and Analysis of the Heat-Shock Protein Gene Superfamily in Bemisia tabaci and Expression Pattern Analysis under Heat Shock. Insects. 2022; 13(7):570. https://doi.org/10.3390/insects13070570
Chicago/Turabian StyleZheng, Hao-Yuan, Peng-Hao Qin, Kun Yang, Tong-Xian Liu, You-Jun Zhang, and Dong Chu. 2022. "Genome-Wide Identification and Analysis of the Heat-Shock Protein Gene Superfamily in Bemisia tabaci and Expression Pattern Analysis under Heat Shock" Insects 13, no. 7: 570. https://doi.org/10.3390/insects13070570
APA StyleZheng, H.-Y., Qin, P.-H., Yang, K., Liu, T.-X., Zhang, Y.-J., & Chu, D. (2022). Genome-Wide Identification and Analysis of the Heat-Shock Protein Gene Superfamily in Bemisia tabaci and Expression Pattern Analysis under Heat Shock. Insects, 13(7), 570. https://doi.org/10.3390/insects13070570