
Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection and Processing
2.3. DNA Extraction, PCR Amplification, and Purification
2.4. Sequence Analysis
2.5. Measurements and Statistical Analysis
3. Results
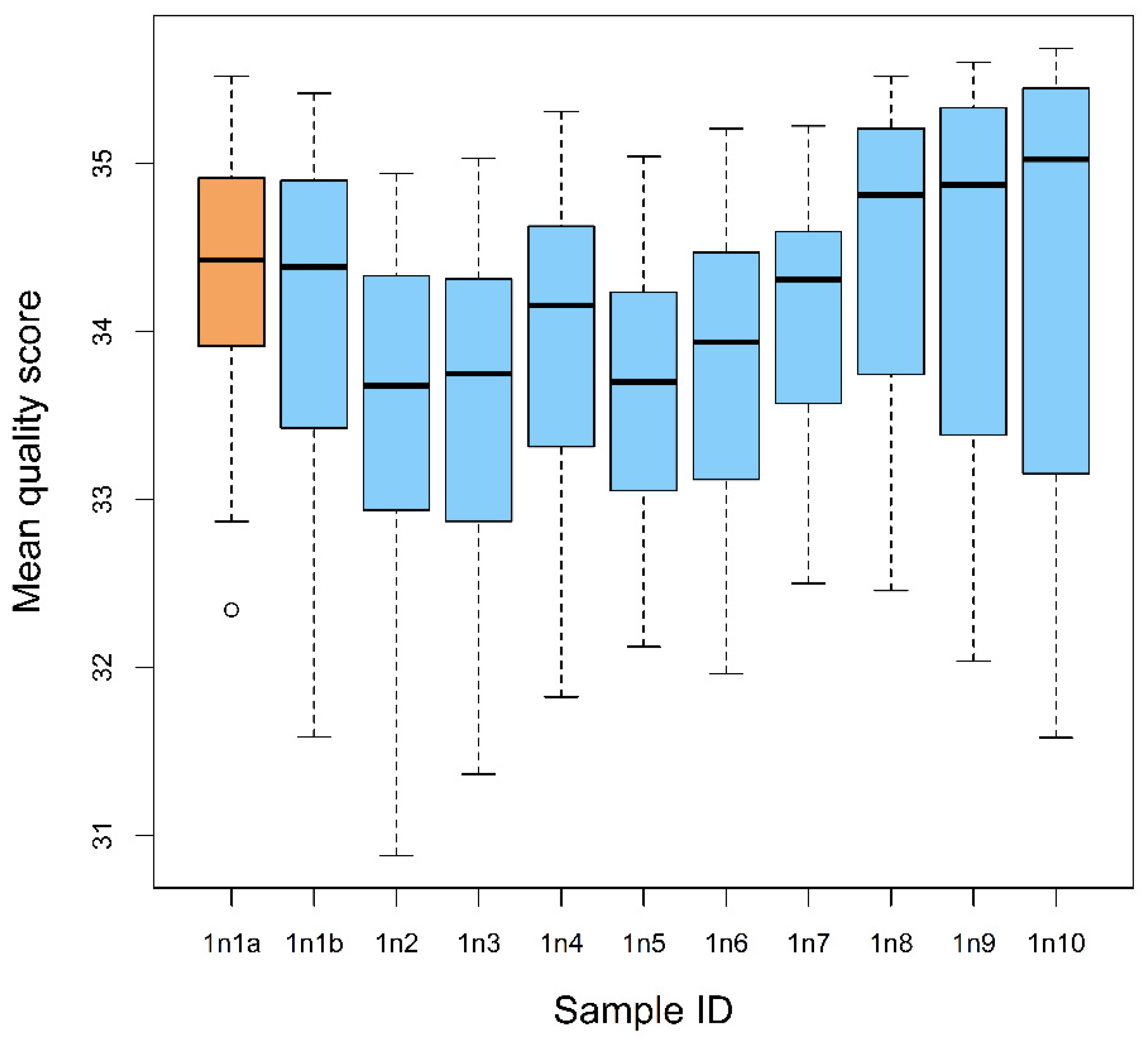
3.1. Basic Sequence Statistics
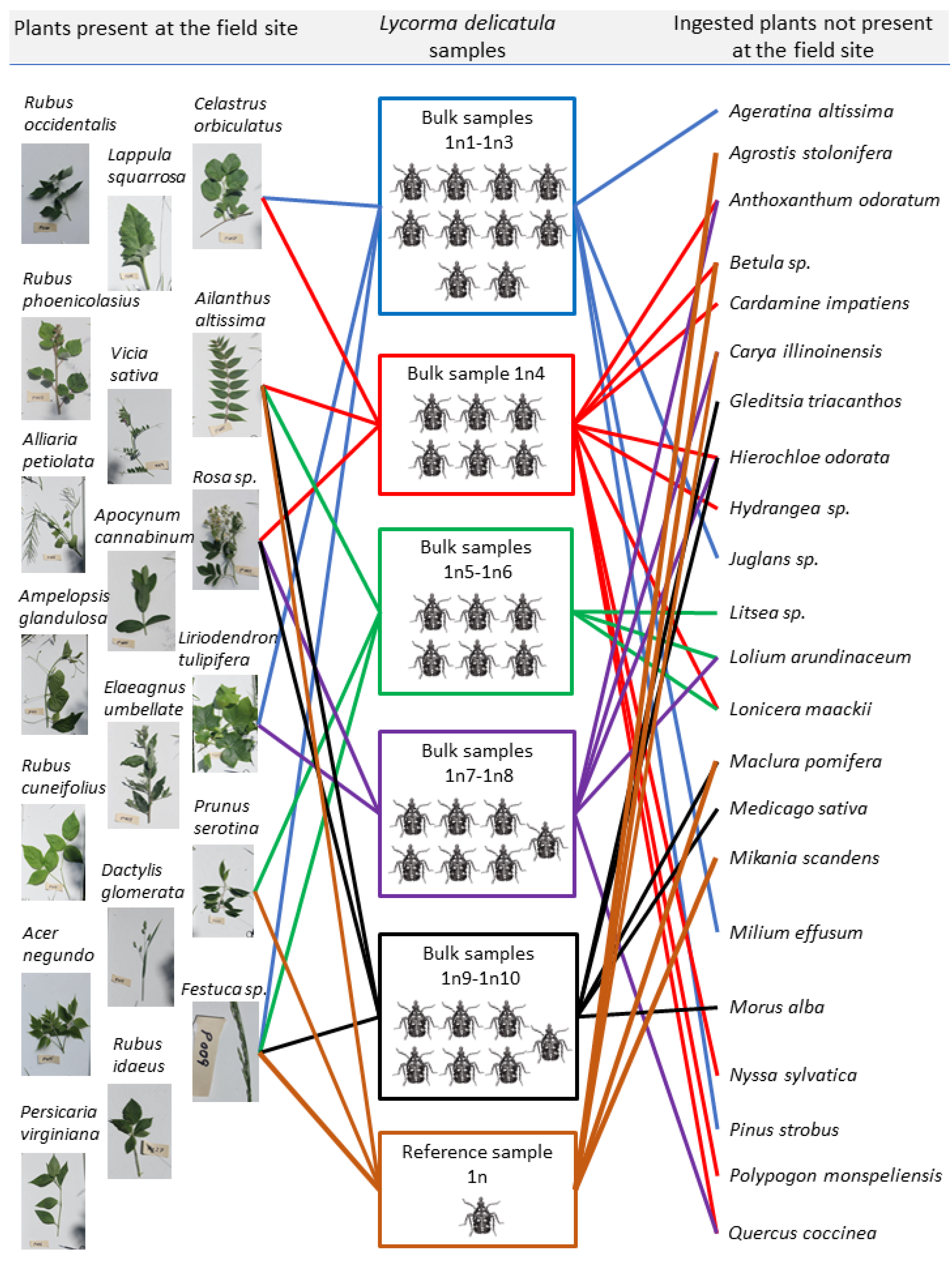
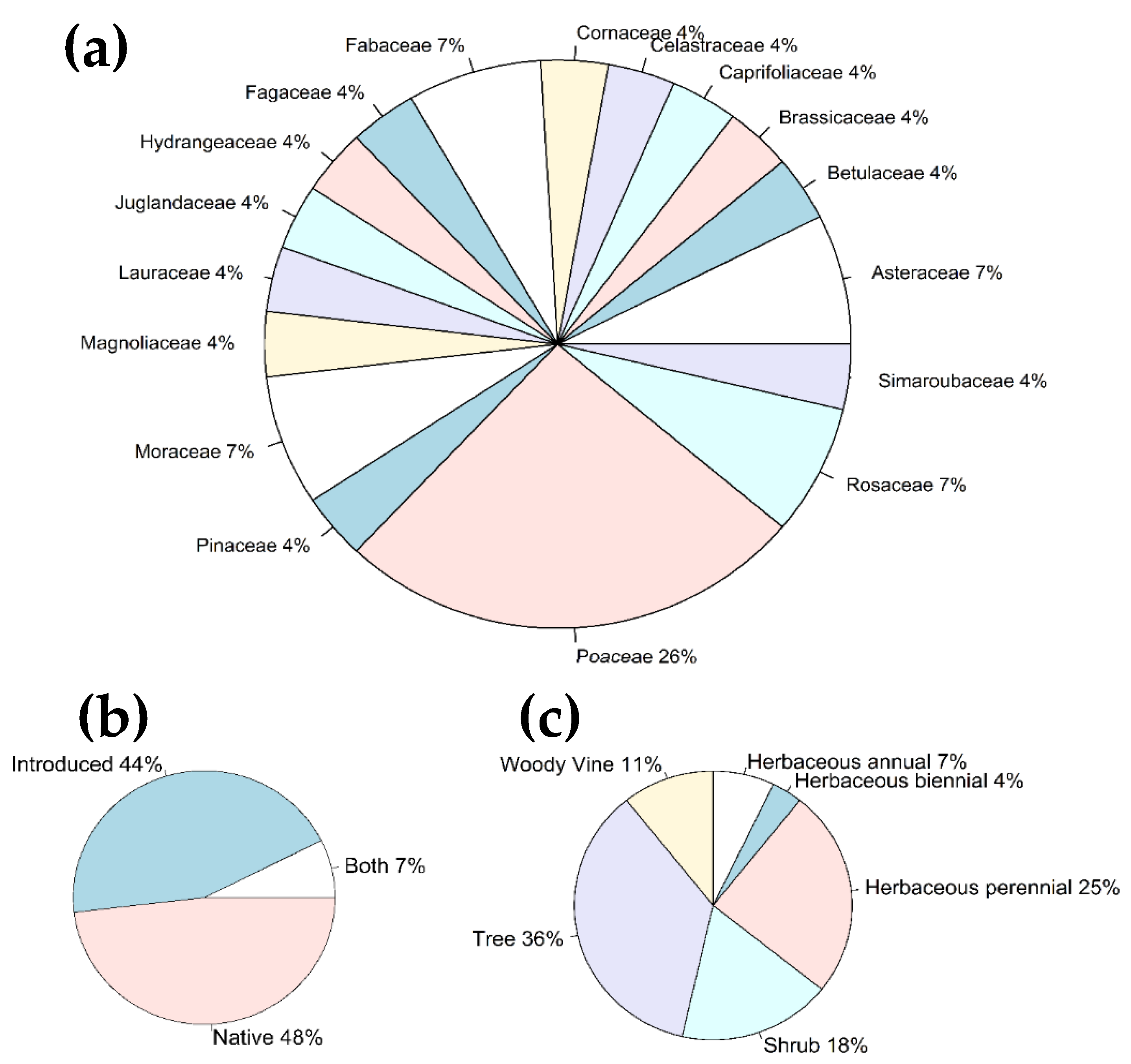
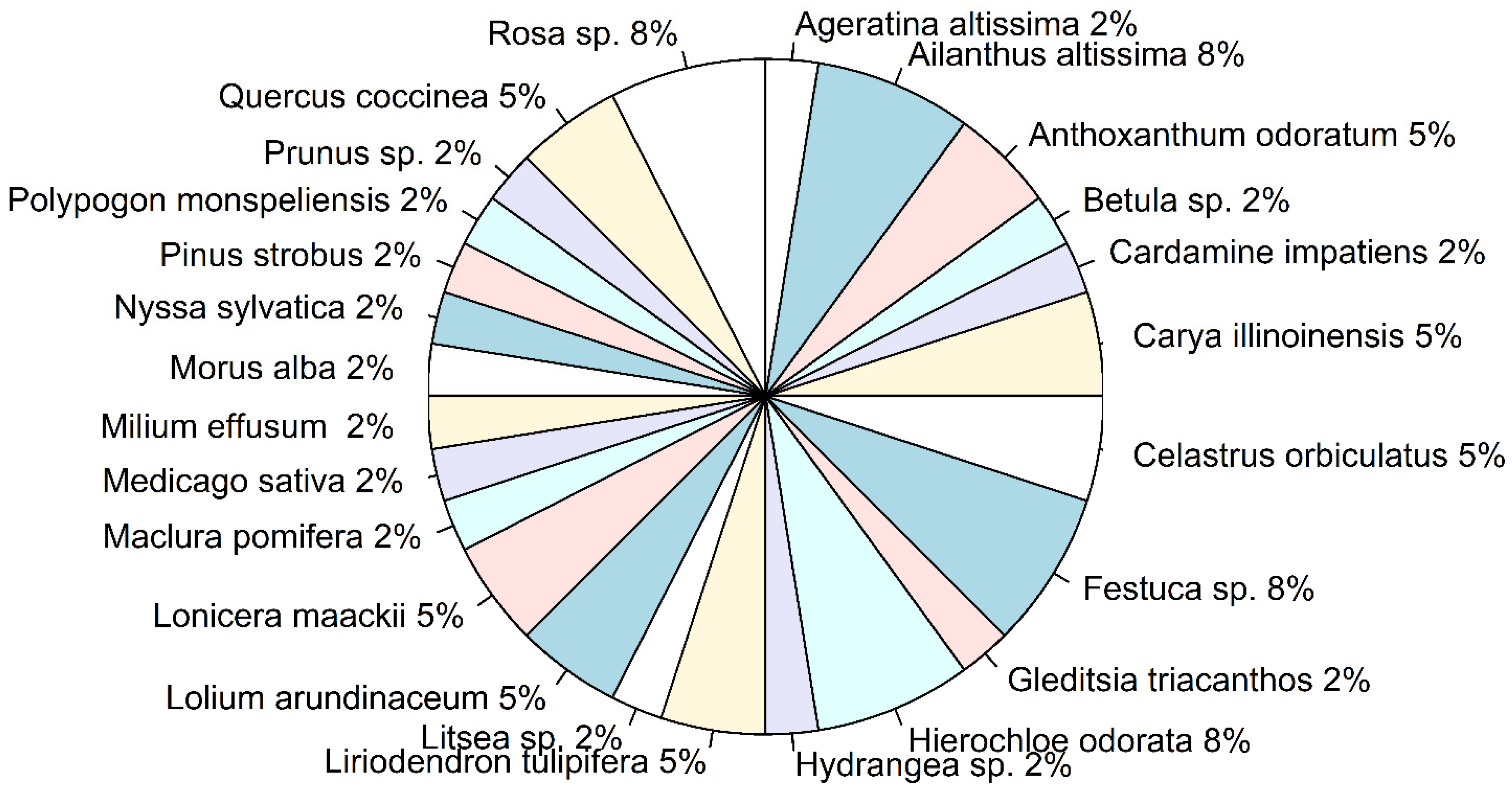
3.2. Diversity of Ingested Plants
4. Discussion
4.1. Diversity of Ingested Plants
4.2. Dispersal and Feeding of L. delicatula Nymphs
4.3. Potential Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.G.; Lee, E.H.; Seo, Y.M.; Kim, N.Y. Cyclic behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on host plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The first New World record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Dara, S.K.; Barringer, L.; Arthurs, S.P. Lycorma delicatula (Hemiptera: Fulgoridae): A new invasive pest in the United States. J. Integr. Pest Manag. 2015, 6, 20. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, Y.L.; Leskey, T.C. A review of biology and management of Lycorma delicatula (Hemiptera: Fulgoridae), an emerging global invasive species. J. Asia-Pac. Entomol. 2019, 22, 589–596. [Google Scholar] [CrossRef]
- Liu, H.; Hartlieb, R.J. Spatial Distribution of Lycorma delicatula (Hemiptera: Fulgoridae) Egg Masses on Tree-of-Heaven, Black Walnut, and Siberian Elm in North America. J. Econ. Entomol. 2019, 113, 1028–1032. [Google Scholar] [CrossRef]
- Avanesyan, A.; Lamp, W.O. Use of molecular gut content analysis to decipher the range of food plants of the invasive spotted lanternfly, Lycorma delicatula. Insects 2020, 11, 215. [Google Scholar] [CrossRef]
- Wakie, T.T.; Neven, L.G.; Yee, W.L.; Lu, Z. The establishment risk of Lycorma delicatula (Hemiptera: Fulgoridae) in the United States and globally. J. Econ. Entomol. 2020, 113, 306–314. [Google Scholar] [CrossRef] [PubMed]
- New York State Integrated Pest Management: Spotted Lanternfly. Available online: https://nysipm.cornell.edu/environment/invasive-species-exotic-pests/spotted-lanternfly/ (accessed on 7 April 2022).
- Barringer, L.; Ciafré, C.M. Worldwide feeding host plants of spotted lanternfly, with significant additions from North America. Environ. Entomol. 2020, 49, 999–1011. [Google Scholar] [CrossRef]
- Uyi, O.; Keller, J.A.; Swackhamer, E.; Hoover, K. Performance and Host Association of Spotted Lanternfly (Lycorma Delicatula) among Common Woody Ornamentals. Sci. Rep. 2021, 11, 15774. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.M.; Calvin, D.; Hills-Stevenson, J. Early Response (2018–2020) to the Threat of Spotted Lanternfly, Lycorma Delicatula (Hemiptera: Fulgoridae) in Pennsylvania. Ann. Entomol. Soc. Am. 2021, 114, 709–718. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Lee, Y.; Lee, H.S.; Lee, S.J.; Lee, J.H. Tracing the Origin of Korean Invasive Populations of the Spotted Lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae). Insects 2021, 12, 539. [Google Scholar] [CrossRef]
- Liu, H. Oviposition substrate selection, egg mass characteristics, host preference, and life history of the spotted lanternfly (Hemiptera: Fulgoridae) in North America. Environ. Entomol. 2019, 48, 1452–1468. [Google Scholar] [CrossRef]
- Cooper, W.R.; Marshall, A.T.; Foutz, J.; Wildung, M.R.; Northfield, T.D.; Crowder, D.W.; Leach, H.; Leskey, T.C.; Halbert, S.E.; Snyder, J.B. Directed Sequencing of Plant Specific DNA Identifies the Dietary History of Four Species of Auchenorrhyncha (Hemiptera). Ann. Entomol. Soc. Am. 2021, 115, 275–284. [Google Scholar] [CrossRef]
- Avanesyan, A.; Sutton, H.; Lamp, W.O. Choosing an effective PCR-based approach for diet analysis of insect herbivores: A systematic review. J. Econ. Entomol. 2021, 114, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Mazzi, D.; Dorn, S. Movement of insect pests in agricultural landscapes. Ann. Appl. Biol. 2012, 160, 97–113. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Brown, R.L.; Sperling, F.A. Tracing an invasion: Phylogeography of Cactoblastis cactorum (Lepidoptera: Pyralidae) in the United States based on mitochondrial DNA. Ann. Entomol. Soc. Am. 2008, 101, 899–905. [Google Scholar] [CrossRef]
- Wang, Q.; Bao, W.F.; Yang, F.; Xu, B.; Yang, Y.Z. The specific host plant DNA detection suggests a potential migration of Apolygus lucorum from cotton to mungbean fields. PLoS ONE 2017, 12, e0177789. [Google Scholar] [CrossRef]
- Wang, Q.; Bao, W.; Zhang, Q.; Fu, X.; Yang, Y.; Lu, Y. Host plant use of a polyphagous mirid, Apolygus lucorum: Molecular evidence from migratory individuals. Ecol. Evol. 2019, 9, 11518–11528. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Kandemir, I.; Walsh, D.B.; Zalom, F.G.; Lavine, L.C. Identification of Lygus hesperus by DNA barcoding reveals insignificant levels of genetic structure among distant and habitat diverse populations. PLoS ONE 2012, 7, e34528. [Google Scholar] [CrossRef][Green Version]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Al Homaidan, A.A.; Al Farhan, A.H.; Khan, H.A. DNA barcoding of arid wild plants using rbcL gene sequences. Genet. Mol. Res. 2012, 11, 1934–1941. [Google Scholar] [CrossRef]
- De la Cadena, G.; Papadopoulou, A.; Maes, J.M.; Gómez-Zurita, J. Evaluation of bias on the assessment of diet breadth of herbivorous insects using molecular methods. Insect Sci. 2017, 24, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gravel, D.; He, F. Seeing is believing? Comparing plant–herbivore networks constructed by field co-occurrence and DNA barcoding methods for gaining insights into network structures. Ecol. Evol. 2019, 9, 1764–1776. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bunce, M.; Dixon, K.W.; Fernandes, K.; Majer, J.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Evaluating Restoration Trajectories Using DNA Metabarcoding of Ground-dwelling and Airborne Invertebrates and Associated Plant Communities. Mol. Ecol. 2022, 31, 2172–2188. [Google Scholar] [CrossRef] [PubMed]
- Dugal, L.; Thomas, L.; Wilkinson, S.P.; Richards, Z.T.; Alexander, J.B.; Adam, A.A.S.; Kennington, W.J.; Jarman, S.; Ryan, N.M.; Bunce, M.; et al. Coral Monitoring in Northwest Australia with Environmental DNA Metabarcoding Using a Curated Reference Database for Optimized Detection. Environ. DNA 2022, 4, 63–76. [Google Scholar] [CrossRef]
- Stat, M.; Huggett, M.J.; Bernasconi, R.; DiBattista, J.D.; Berry, T.E.; Newman, S.J.; Harvey, E.S.; Bunce, M. Ecosystem Biomonitoring with EDNA: Metabarcoding across the Tree of Life in a Tropical Marine Environment. Sci. Rep. 2017, 7, 12240. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bunce, M.; Wardell-Johnson, G.; Fernandes, K.; White, N.E.; Nevill, P. Testing multiple substrates for terrestrial biodiversity monitoring using environmental DNA metabarcoding. Mol. Ecol. Res. 2020, 20, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bateman, P.W.; Bunce, M.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Scat DNA provides important data for effective monitoring of mammal and bird biodiversity. Biodivers. Conserv. 2021, 30, 3585–3602. [Google Scholar] [CrossRef]
- Avanesyan, A.; Illahi, N.; Lamp, W.O. Detecting ingested host plant DNA in potato leafhopper (Hemiptera: Cicadellidae): Potential use of molecular markers for gut content analysis. J. Econ. Entomol. 2021, 114, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Siegert, C.M.; Levia, D.F. Seasonal and meteorological effects on differential stemflow funneling ratios for two deciduous tree species. J. Hydrol. 2014, 519, 446–454. [Google Scholar] [CrossRef]
- Avanesyan, A. Plant DNA detection from grasshopper gut contents: A step-by-step protocol, from tissues preparation to obtaining plant DNA sequences. Appl. Plant Sci. 2014, 2, 1300082. [Google Scholar] [CrossRef]
- Avanesyan, A.; Culley, T.M. Feeding preferences of Melanoplus femurrubrum grasshoppers on native and exotic grasses: Behavioral and molecular approaches. Entomol. Exp. Appl. 2015, 157, 152–163. [Google Scholar] [CrossRef]
- Goecks, J.; Nekrutenko, A.; Taylor, J. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 1 April 2022).
- Dechaine, A.C.; Sutphin, M.; Leskey, T.C.; Salom, S.M.; Kuhar, T.P.; Pfeiffer, D.G. Phenology of Lycorma delicatula (Hemiptera: Fulgoridae) in Virginia, USA. Environ. Entomol. 2021, 50, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Kim, S.; Kwon, S.W.; Lee, S.I.; Jablonski, P.G. Defense sequestration associated with narrowing of diet and ontogenetic change to aposematic colours in the spotted lanternfly. Sci. Rep. 2018, 8, 16831. [Google Scholar] [CrossRef] [PubMed]
- Nixon, L.J.; Jones, S.K.; Tang, L.; Urban, J.; Felton, K.; Leskey, T.C. Survivorship and development of the invasive Lycorma delicatula (Hemiptera: Fulgoridae) on wild and cultivated temperate host plants. Environ. Entomol. 2022, 51, 222–228. [Google Scholar] [CrossRef]
- Pearson, C.C.; Backus, E.A.; Shugart, H.J.; Munyaneza, J.E. Characterization and correlation of EPG waveforms of Bactericera cockerelli (Hemiptera: Triozidae): Variability in waveform appearance in relation to applied signal. Ann. Entomol. Soc. Am. 2014, 107, 650–666. [Google Scholar] [CrossRef]
- Essler, J.L.; Kane, S.A.; Collins, A.; Ryder, K.; DeAngelo, A.; Kaynaroglu, P.; Otto, C.M. Egg Masses as Training Aids for Spotted Lanternfly Lycorma Delicatula Detection Dogs. PLoS ONE 2021, 16, e0250945. [Google Scholar] [CrossRef]
- Keller, J.A.; Johnson, A.E.; Uyi, O.; Wurzbacher, S.; Long, D.; Hoover, K. Dispersal of Lycorma delicatula (Hemiptera: Fulgoridae) nymphs through contiguous, deciduous forest. Environ. Entomol. 2020, 49, 1012–1018. [Google Scholar] [CrossRef]
- Wolfin, M.S.; Binyameen, M.; Wang, Y.; Urban, J.M.; Roberts, D.C.; Baker, T.C. Flight dispersal capabilities of female spotted lanternflies (Lycorma delicatula) related to size and mating status. J. Insect Behav. 2019, 32, 188–200. [Google Scholar] [CrossRef]
- Baker, T.C.; Smyers, E.C.; Urban, J.M.; Meng, Z.; Damadaram, K.P.; Myrick, A.J.; Cooperband, M.F.; Domingue, M.J. Progression of seasonal activities of adults of the spotted lanternfly, Lycorma delicatula, during the 2017 season of mass flight dispersal behavior in eastern Pennsylvania. J. Asia-Pac. Entomol. 2019, 22, 705–713. [Google Scholar] [CrossRef]
- Baker, T.C.; Myrick, A.J.; Wolfin, M.S.; Wang, Y. Visual Responses of Flight-Dispersing Spotted Lanternflies, Lycorma delicatula toward a Tall Vertical Silhouette in a Vineyard. J. Insect Behav. 2021, 34, 49–60. [Google Scholar] [CrossRef]
- Domingue, M.J.; Baker, T.C. Orientation of flight for physically disturbed spotted lanternflies, Lycorma delicatula,(Hemiptera, Fulgoridae). J. Asia-Pac. Entomol. 2019, 22, 117–120. [Google Scholar] [CrossRef]
- Cooperband, M.; Murman, K.; Cannon, S.; Abreu, L.; Wallace, M. Dispersal and host preference of marked and released spotted lanternfly. In Otis Laboratory 2018 Annual Report; Trepanowski, N., Vieira, K., Heller, S., Booth, E., Eds.; United States Department of Agriculture: Buzzards Bay, MA, USA, 2019; p. 97. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Plant Family | Plant Species ID | Common Name | Nymphs Presence | Date Submitted to GenBank | GenBank Accession Number |
|---|---|---|---|---|---|---|
| p001 | Rosaceae | Rosa sp. | Rose | + | 12/27/2021 | OM038103 |
| p002 | Rosaceae | Rubus phoenicolasius | Wine raspberry | + | 12/27/2021 | OM038104 |
| p003 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 12/27/2021 | OM038105 |
| p004 | Apocynaceae | Apocynum cannabinum | Indian hemp | − | 12/27/2021 | OM038106 |
| p005 | Simaroubaceae | Ailanthus altissima | Tree of heaven | + | 1/31/2022 | OM470964 |
| p006 | Polygonaceae | Persicaria virginiana | Jumpseed | − | 1/31/2022 | OM470965 |
| p007 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | + | 11/7/2021 | OL539536 |
| p008 | Brassicaceae | Alliaria petiolata | Garlic mustard | − | 11/21/2021 | OL603937 |
| p009 | Poaceae | Festuca sp. | Fescue | − | 11/24/2021 | OL657222 |
| p010 | Fabaceae | Vicia sativa | Common vetch | − | 11/24/2021 | OL657223 |
| p011 | Vitaceae | Ampelopsis glandulosa | Amur peppervine | − | 1/31/2022 | OM470966 |
| p012 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 1/31/2022 | OM470967 |
| p013 | Poaceae | Dactylis glomerata | Cat grass | − | 2/4/2022 | OM523099 |
| p014 | Rosaceae | Rubus cuneifolius | Sand blackberry | − | 2/4/2022 | OM523098 |
| p015 | Sapindaceae | Acer negundo | Boxelder maple | − | 2/11/2022 | OM672995 |
| p016 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 2/11/2022 | OM672996 |
| p017 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 2/4/2022 | OM523100 |
| p018 | Boraginaceae | Lappula squarrosa | Bristly sheepburr | − | 2/4/2022 | OM523101 |
| p019 | Poaceae | Dactylis glomerata | Cat grass | − | 2/11/2022 | OM672997 |
| p020 | Rosaceae | Rubus occidentalis | Black Raspberry | − | 2/11/2022 | OM672998 |
| p021 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 2/11/2022 | OM672999 |
| p022 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 3/8/2022 | OM964639 |
| p023 | Vitaceae | Ampelopsis glandulosa | Amur peppervine | − | 2/11/2022 | OM673000 |
| p024 | Sapindaceae | Acer negundo | Boxelder maple | − | 2/11/2022 | OM673001 |
| p025 | Magnoliaceae | Liriodendron tulipifera | Tulip tree | − | 2/11/2022 | OM673002 |
| p026 | Rosaceae | Prunus serotina | Black cherry | − | 2/11/2022 | OM673003 |
| p027 | Rosaceae | Rubus idaeus | Red raspberry | − | 2/11/2022 | OM673004 |
| p028 | Boraginaceae | Lappula squarrosa | Bristly sheepburr | − | 2/11/2022 | OM673005 |
| Primers and Other PCR Components | Concentration/Volume (for 20 μL of PCR Reaction) | PCR Conditions |
|---|---|---|
| rbcLaF (5′-ATG TCA CCA CAA ACA GAG ACT AAA GC-3′) | 2 μM/2 μL | initial denaturation: 94 °C for 4 min 35 cycles of 94 °C for 30 s, 57 °C for 30 s, and 72 °C for 30 s final extension: 72 °C for 2 min |
| rbcLaR (5′-GTA AAA TCA AGT CCA CCR CG-3′) | 2 μM/2 μL | |
| 2X PCR PreMix with Dye (Syd Labs Inc., Natick, MA, USA) | 10 μL | |
| ddH2O | 5.2 μL | |
| template DNA | 0.8 μL |
| Sample ID | Total Number of Sequence Reads | Sequence Read Length (bp) | Sequence Length Screened (bp; Consensus Sequences) | Number of Unique Readable Sequences Passed the Screening | Number of Unique Plant Species | Sequence Quality Score (Mean ± SE) * | |
|---|---|---|---|---|---|---|---|
| Forward | Reverse | ||||||
| 1n1a-ref | 347,901 | 347,901 | 35–250 | >150 | 65 | 11 | 34.37 ± 0.11 ac |
| 1n1b | 223,629 | 223,629 | 35–250 | >100 | 3 | 3 | 34.11 ± 0.12 ac |
| 1n2 | 117,181 | 117,181 | 35–250 | >100 | 1 | 1 | 33.53 ± 0.12 bc |
| 1n3 | 176,223 | 176,223 | 35–250 | >100 | 9 | 3 | 33.59 ± 0.11 c |
| 1n4 | 157,468 | 157,468 | 35–250 | >100 | 19 | 12 | 33.95 ± 0.10 c |
| 1n5 | 169,266 | 169,266 | 35–250 | >100 | 7 | 3 | 33.67 ± 0.09 bc |
| 1n6 | 138,802 | 138,802 | 35–250 | >100 | 5 | 3 | 33.84 ± 0.10 bc |
| 1n7 | 163,404 | 163,404 | 35–250 | >100 | 11 | 6 | 34.11 ± 0.08 ac |
| 1n8 | 160,706 | 160,706 | 35–250 | >100 | 3 | 1 | 34.53 ± 0.11 a |
| 1n9 | 195,474 | 195,474 | 35–250 | >100 | 7 | 5 | 34.46 ± 0.14 a |
| 1n10 | 198,622 | 198,622 | 35–250 | >100 | 4 | 3 | 34.43 ± 0.16 ac |
| Plant Family | Plant Species | Common Name | Seq. Length (bp) | Highest Match, % | Plant Origin | Plant Life Form | Presence at the Site | Reported As a Host * | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Asteraceae | Ageratina altissima | White snakeroot | 285 | 99.65 | Native | Herbaceous Perennial | − | − |
| 2 | Asteraceae | Mikania scandens | Climbing hempvine | 225 | 99.55 | Native | Woody Vine | − | − |
| 3 | Betulaceae | Betula sp. | Birch | 406 | 100 | Both | Tree | − | + |
| 4 | Brassicaceae | Cardamine impatiens | Narrowleaf bittercress | 258 | 100 | Introduced | Herbaceous Biennial | − | − |
| 5 | Caprifoliaceae | Lonicera maackii | Amur honeysuckle | 247 | 100 | Introduced | Shrub | − | + |
| 6 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | 404 | 100 | Introduced | Woody Vine | + | + |
| 7 | Cornaceae | Nyssa sylvatica | Blackgum | 362 | 100 | Native | Tree | − | + |
| 8 | Fabaceae | Gleditsia triacanthos | Honeylocust | 260 | 99.2 | Native | Shrub | − | − |
| 9 | Fabaceae | Medicago sativa | Alfalfa | 184 | 100 | Introduced | Herbaceous Perennial | − | + |
| 10 | Fagaceae | Quercus coccinea | Scarlet oak | 336 | 99.11 | Native | Tree | − | + |
| 11 | Hydrangeaceae | Hydrangea sp. | Hydrangea | 115 | 100 | Both | Shrub | − | − |
| 12 | Juglandaceae | Carya illinoinensis | Pecan | 367 | 100 | Native | Tree | − | + |
| 13 | Lauraceae | Litsea sp. | Litsea | 260 | 99.6 | Native | Shrub | − | − |
| 14 | Magnoliaceae | Liriodendron tulipifera | Tuliptree | 292 | 99 | Native | Tree | + | + |
| 15 | Moraceae | Maclura pomifera | Osage-orange | 190 | 100 | Native | Tree | − | − |
| 16 | Moraceae | Morus alba | White mulberry | 357 | 99.72 | Introduced | Tree | − | + |
| 17 | Pinaceae | Pinus strobus | Eastern white pine | 239 | 100 | Native | Tree | − | + |
| 18 | Poaceae | Agrostis stolonifera | Creeping bentgrass | 158 | 100 | Introduced | Herbaceous perennial | − | − |
| 19 | Poaceae | Anthoxanthum odoratum | Sweet vernalgrass | 150 | 100 | Introduced | Herbaceous Perennial | − | − |
| 20 | Poaceae | Festuca sp. | Fescue | 206 | 100 | Introduced | Herbaceous Perennial | + | + |
| 21 | Poaceae | Hierochloe odorata | Sweetgrass | 218 | 100 | Native | Herbaceous Perennial | − | − |
| 22 | Poaceae | Lolium arundinaceum | Tall fescue | 346 | 99.7 | Introduced | Herbaceous Annual | − | − |
| 23 | Poaceae | Milium effusum | American milletgrass | 110 | 100 | Native | Herbaceous Perennial | − | − |
| 24 | Poaceae | Polypogon monspeliensis | Annual rabbitsfoot grass | 127 | 99.21 | Introduced | Herbaceous Annual | − | − |
| 25 | Rosaceae | Prunus sp. | Plum | 200 | 100 | Native | Tree/Shrub | + | + |
| 26 | Rosaceae | Rosa sp. | Rose | 259 | 100 | Introduced | Woody Vine | + | + |
| 27 | Simaroubaceae | Ailanthus altissima | Tree of heaven | 300 | 99.67 | Introduced | Tree | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McPherson, C.; Avanesyan, A.; Lamp, W.O. Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding. Insects 2022, 13, 534. https://doi.org/10.3390/insects13060534
McPherson C, Avanesyan A, Lamp WO. Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding. Insects. 2022; 13(6):534. https://doi.org/10.3390/insects13060534
Chicago/Turabian StyleMcPherson, Cameron, Alina Avanesyan, and William O. Lamp. 2022. "Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding" Insects 13, no. 6: 534. https://doi.org/10.3390/insects13060534
APA StyleMcPherson, C., Avanesyan, A., & Lamp, W. O. (2022). Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding. Insects, 13(6), 534. https://doi.org/10.3390/insects13060534