
Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation

 ,
,  , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Aphid Resistance Evaluation in the Laboratory
2.3. Aphid Growth and Development
2.4. Total Phenol Compounds Content Assay
2.5. Total Flavonoid Content Assay
2.6. Statistical Analysis
3. Results
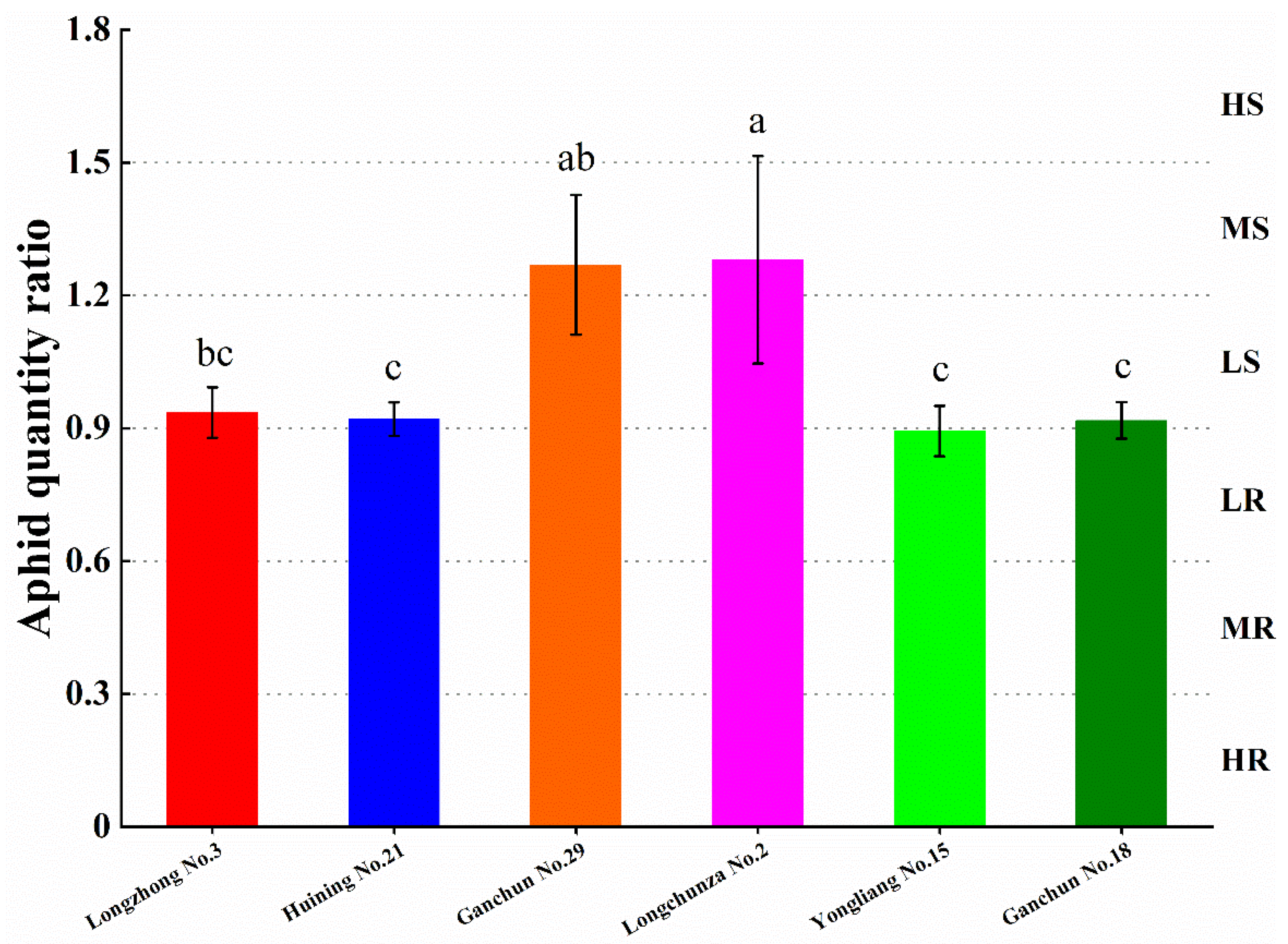
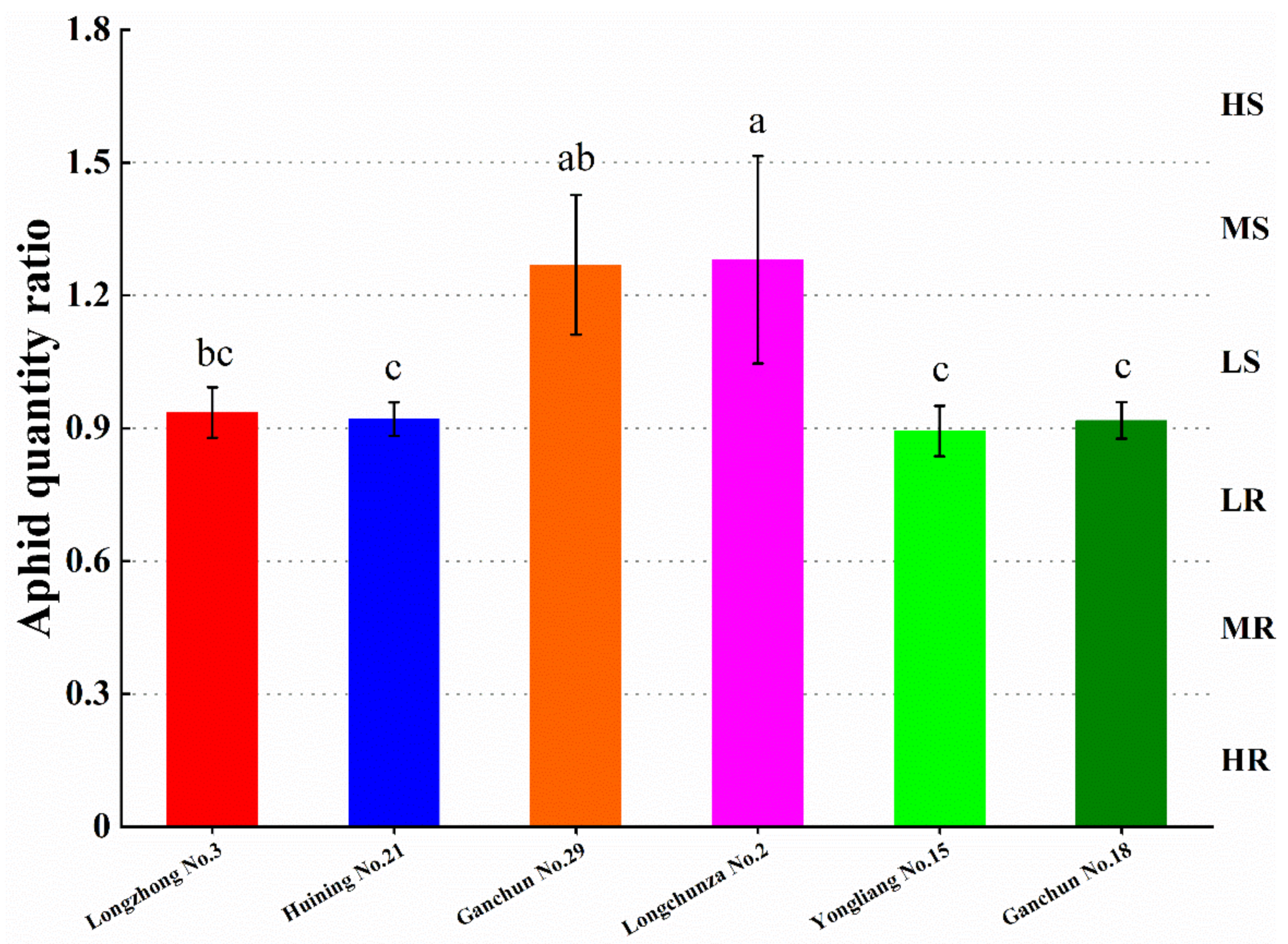
3.1. Evaluation of Wheat Resistance to S. avenae
3.2. Biological Parameters of S. avenae on Different Wheat Varieties
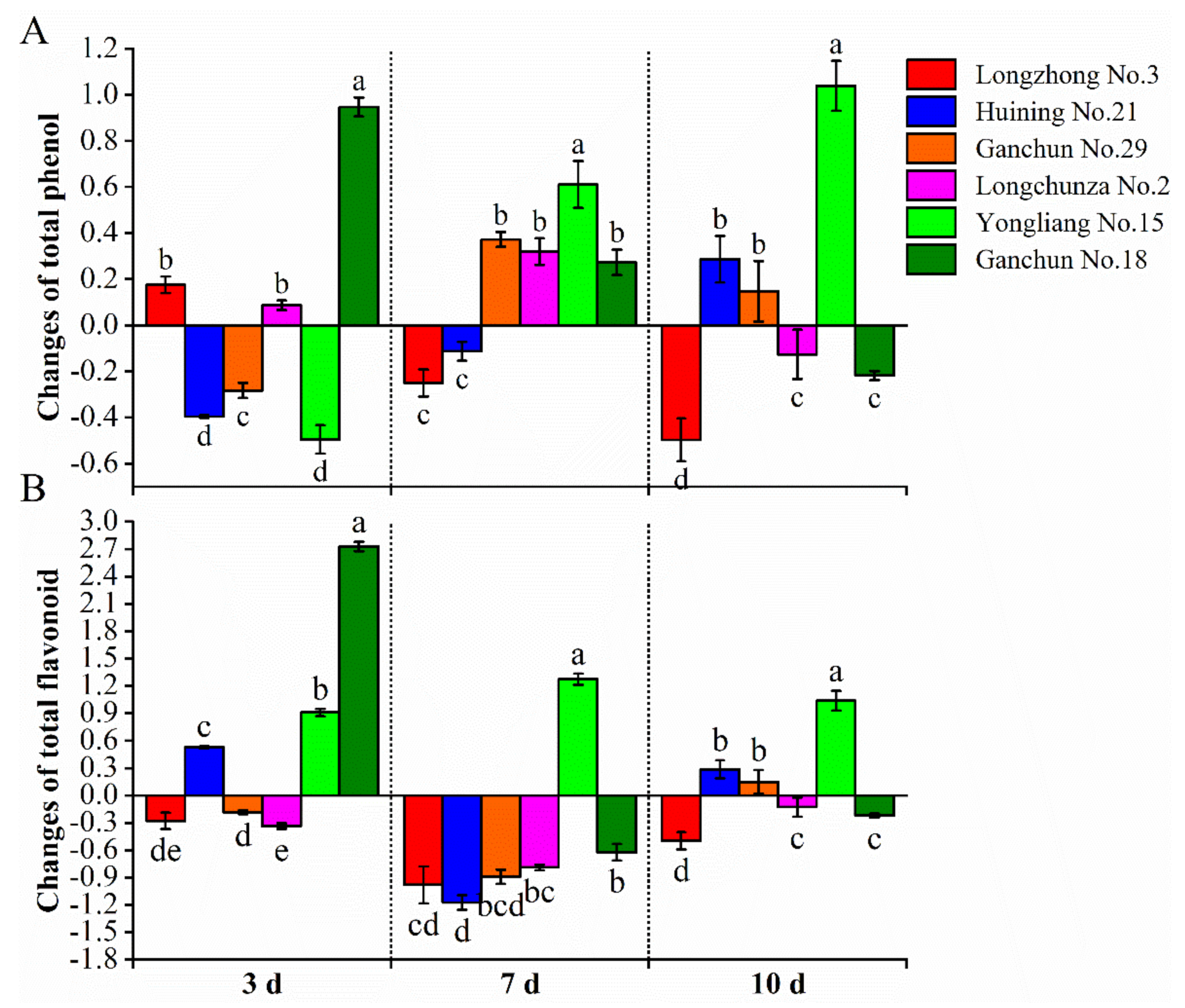
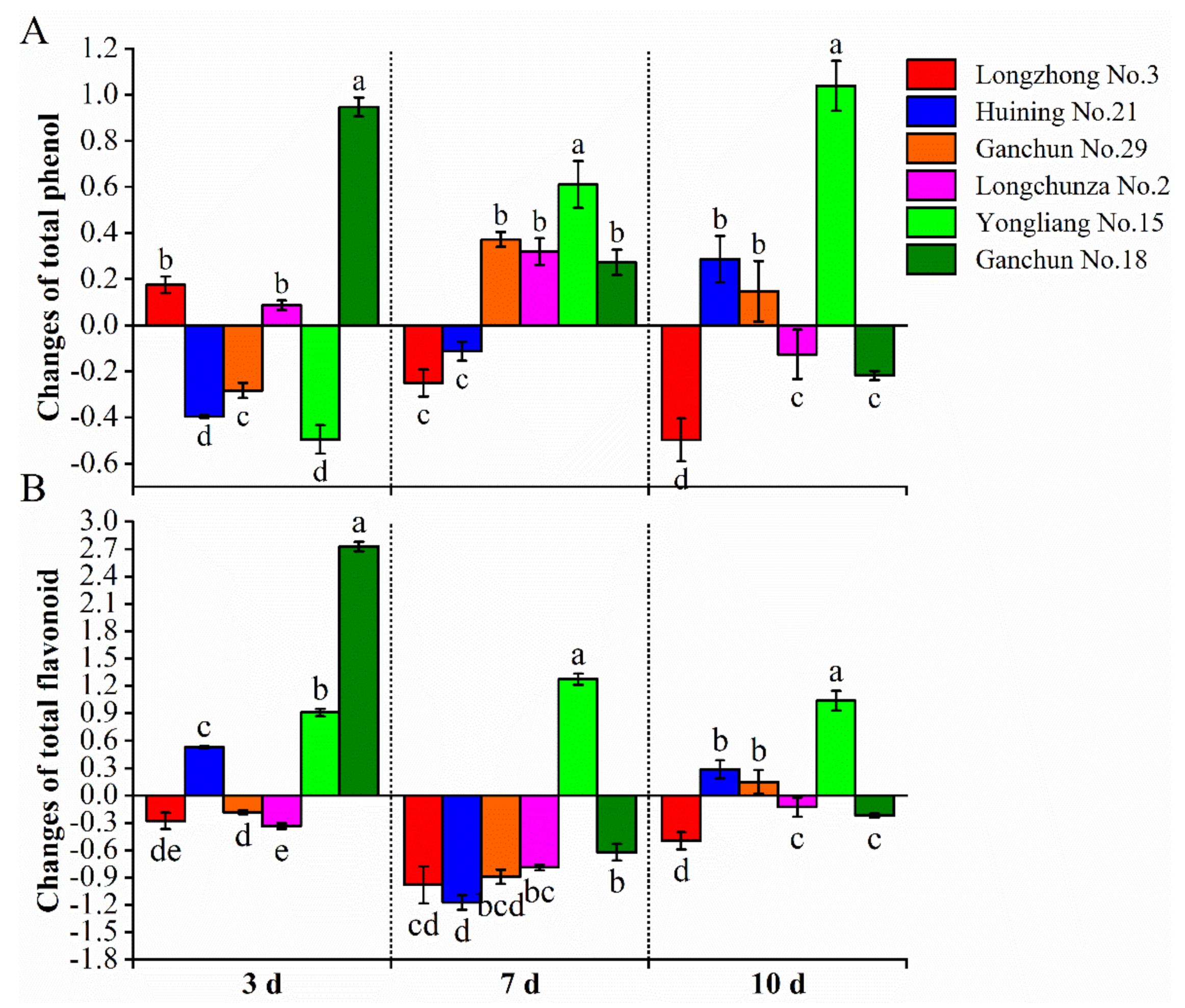
3.3. Total Phenol Compound Content
3.4. Flavonoid Content
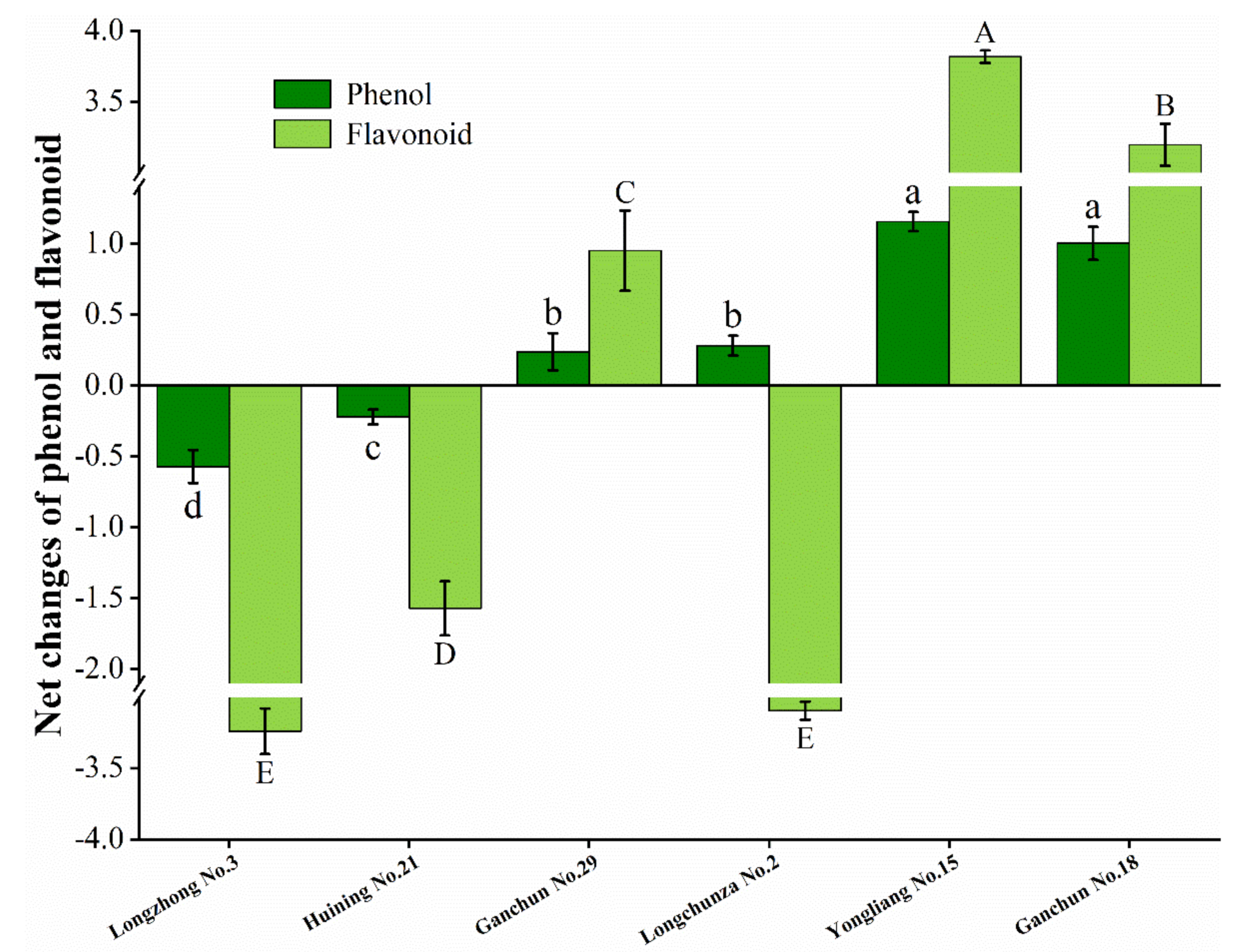
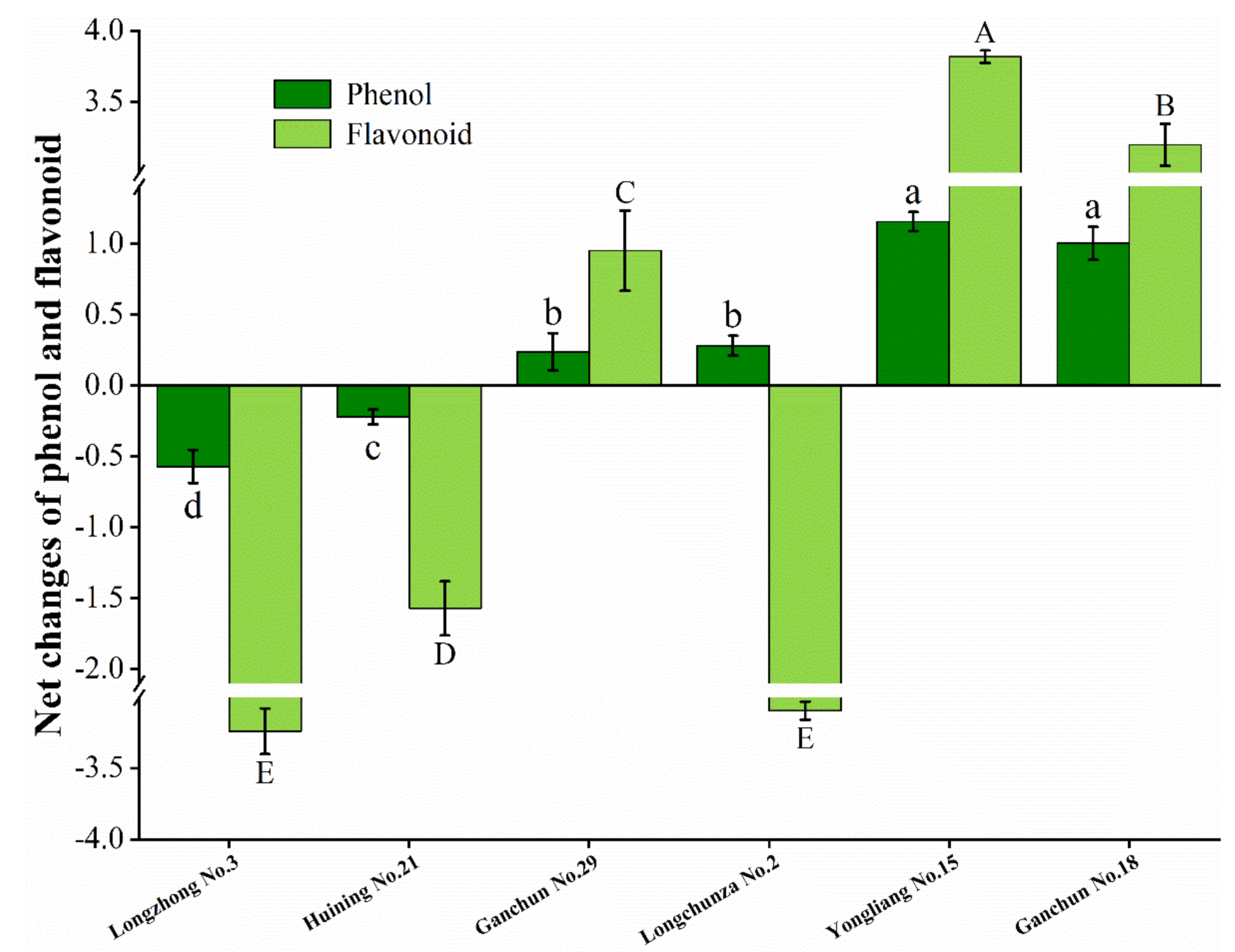
3.5. Net Changes of Total Phenol Content and Flavonoid Content
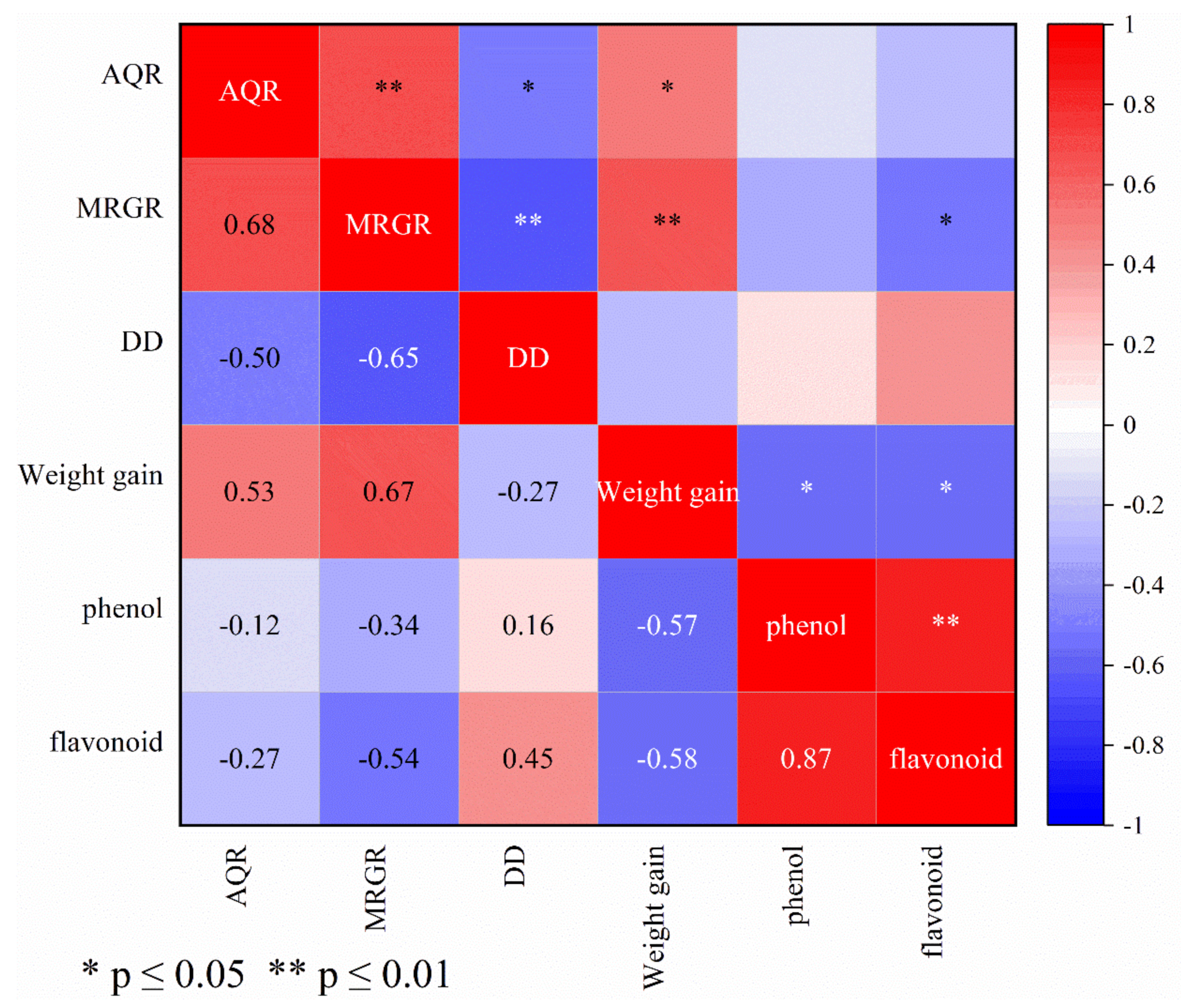
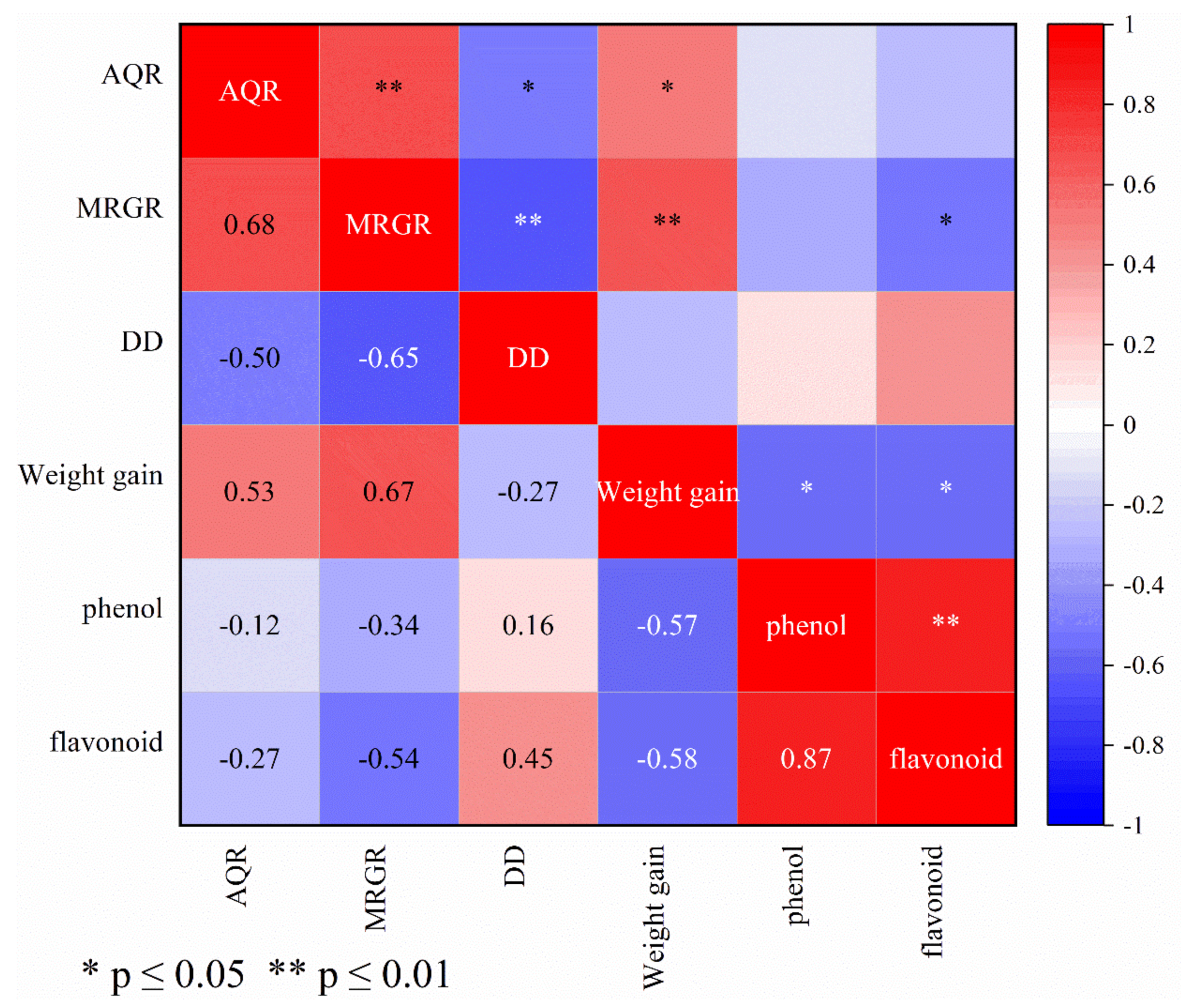
3.6. Correlation between Wheat Resistance to S. avenae and Biological Parameters of Wheat Aphid and Accumulation of Secondary Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, P.K.; Mir, R.R.; Mohan, A.; Kumar, J. Wheat genomics: Present status and future prospects. Int. J. Plant Genom. 2008, 2008, 896451. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, M.S.; Bhavani, S.; Singh, P.K.; Huerta-Espino, J.; Singh, R.P. Disease Resistance in Wheat: Present Status and Future Prospects. In Disease Resistance in Crop Plants; Wani, S.H., Ed.; Springer: Cham, Switzerland, 2019; pp. 61–81. ISBN 978-3-030-20727-4. [Google Scholar]
- Luo, K.; Zhao, H.; Wang, X.; Kang, Z. Prevalent Pest Management Strategies for Grain Aphids: Opportunities and Challenges. Front. Plant Sci. 2021, 12, 790919. [Google Scholar] [CrossRef] [PubMed]
- Awmack, C.S.; Harrington, R.; Leather, S.R.; Lawton, J.H. The impacts of elevated CO2 on aphid-plant interactions. Asp. Appl. Biol. 1996, 45, 317–322. [Google Scholar]
- Li, Q.; Sun, J.; Qin, Y.; Fan, J.; Zhang, Y.; Tan, X.; Hou, M.; Chen, J. Reduced insecticide susceptibility of the wheat aphid Sitobion miscanthi after infection by the secondary bacterial symbiont Hamiltonella defensa. Pest Manag. Sci. 2021, 77, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, J.; Wu, X.B.; Deng, Q.Q.; Zhu, Z.Y.; Ren, M.J.; Ye, M.; Zeng, R.S. Seed priming with calcium chloride enhances wheat resistance against wheat aphid Schizaphis graminum Rondani. Pest Manag. Sci. 2021, 77, 4709–4718. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, D.; Liu, X.; Wang, B.; Shi, X. Divergence of Desiccation-Related Traits in Sitobion avenae from Northwestern China. Insects 2020, 11, 626. [Google Scholar] [CrossRef]
- He, Y.; Liu, D.; Dai, P.; Wang, D.; Shi, X. Genetic Differentiation and Structure of Sitobion avenae (Hemiptera: Aphididae) Populations from Moist, Semiarid and Arid Areas in Northwestern China. J. Econ. Entomol. 2018, 111, 603–611. [Google Scholar] [CrossRef]
- Dai, P.; Liu, D.; Shi, X. Impacts of Water Deficiency on Life History of Sitobion avenae Clones From Semi-arid and Moist Areas. J. Econ. Entomol. 2015, 108, 2250–2258. [Google Scholar] [CrossRef]
- Xu, Z.H.; Chen, J.L.; Cheng, D.F.; Sun, J.R.; Liu, Y.; Francis, F. Discovery of English grain aphid (Hemiptera: Aphididae) biotypes in China. J. Econ. Entomol. 2011, 104, 1080–1086. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, F.; Coates, B.; Wei, C.; Zhu, X.; Zhang, Y.; Zhou, X. Temporal analysis of microRNAs associated with wing development in the English grain aphid, Sitobion avenae (F.) (Homoptera: Aphidiae). Insect Biochem. Mol. Biol. 2022, 142, 103579. [Google Scholar] [CrossRef]
- Dogimont, C.; Bendahmane, A.; Chovelon, V.; Boissot, N. Host plant resistance to aphids in cultivated crops: Genetic and molecular bases, and interactions with aphid populations. Comptes Rendus Biol. 2010, 333, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Ge, F.; Zhu, S.; Parajulee, M.N. Effect of Cotton Cultivar on Development and Reproduction of Aphis gossypii (Homoptera: Aphididae) and Its Predator Propylaea japonica (Coleoptera: Coccinellidae). J. Econ. Entomol. 2004, 97, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [Green Version]
- Marimuthu, M.; Smith, C.M. Barley tolerance of Russian wheat aphid (Hemiptera: Aphididae) biotype 2 herbivory involves expression of defense response and developmental genes. Plant Signal. Behav. 2012, 7, 382–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Leitner, M.; Boland, W.; Mithöfer, A. Direct and indirect defences induced by piercing-sucking and chewing herbivores in Medicago truncatula. New Phytol. 2005, 167, 597–606. [Google Scholar] [CrossRef]
- Mumm, R.; Hilker, M. Direct and indirect chemical defence of pine against folivorous insects. Trends Plant Sci. 2006, 11, 351–358. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant growth regulator mediated consequences of secondary metabolites in medicinal plants. J. Appl. Res. Med. Aroma. 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Pan, L.; Ren, L.; Chen, F.; Feng, Y.; Luo, Y. Antifeedant Activity of Ginkgo biloba Secondary Metabolites against Hyphantria cunea Larvae: Mechanisms and Applications. PLoS ONE 2016, 11, e0155682. [Google Scholar] [CrossRef]
- Sharma, H.C.; Sujana, G.; Manohar Rao, D. Morphological and chemical components of resistance to pod borer, Helicoverpa armigera in wild relatives of pigeonpea. Arthropod Plant Interact. 2009, 3, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Sun, X.; Zhao, H.P.; Xue, M.; Wang, D. Phenolic compounds induced by Bemisia tabaci and Trialeurodes vaporariorum in Nicotiana tabacum L. and their relationship with the salicylic acid signaling pathway. Arthropod Plant Interact. 2017, 11, 659–667. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Goławska, S.; Łukasik, I. Acceptance of low-saponin lines of alfalfa with varied phenolic concentrations by pea aphid (Homoptera: Aphididae). Biologia 2009, 64, 377–382. [Google Scholar] [CrossRef]
- Jördens-Röttger, D. The Role of Phenolic Substances for Host-Selection Behaviour of the Black Bean Aphid, Aphis Fabae. Entomol. Exp. Appl. 1979, 26, 49–54. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of Polyphenols in the Resistance Mechanisms of Plants Against Fungal Pathogens and Insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Trivandrum, India, 2006; Volume 37, pp. 23–67. ISBN 81-308-0034-9. [Google Scholar]
- Moctezuma, C.; Hammerbacher, A.; Heil, M.; Gershenzon, J.; Mendez-Alonzo, R.; Oyama, K. Specific polyphenols and tannins are associated with defense against insect herbivores in the tropical oak Quercus oleoides. J. Chem. Ecol. 2014, 40, 458–467. [Google Scholar] [CrossRef]
- Painter, R.H. Resistance of Plants to Insects. Annu. Rev. Entomol. 1958, 3, 267–290. [Google Scholar] [CrossRef]
- Zhang, Z.; Lan, H.; Cao, H.; Hu, X.; Fan, Y.; Song, Y.; Wu, L.; Liu, T.X. Impacts of Constitutive and Induced Benzoxazinoids Levels on Wheat Resistance to the Grain Aphid (Sitobion avenae). Metabolites 2021, 11, 783. [Google Scholar] [CrossRef]
- Tu, X.B.; Fan, Y.L.; McNeill, M.; Zhang, Z.H. Including predator presence in a refined model for assessing resistance of alfalfa cultivar to aphids. J. Integr. Agr. 2018, 17, 397–405. [Google Scholar] [CrossRef]
- Leather, S.R.; Dixon, A.F.G. Aphid growth and reproductive rates. Entomol. Exp. Appl. 1984, 35, 137–140. [Google Scholar] [CrossRef]
- Chlopicka, J.; Pasko, P.; Gorinstein, S.; Jedryas, A.; Zagrodzki, P. Total phenolic and total flavonoid content, antioxidant activity and sensory evaluation of pseudocereal breads. LWT-Food Sci. Technol. 2012, 46, 548–555. [Google Scholar] [CrossRef]
- Wu, J.X.; Jiang, J.H.; Shen, B.C.; Su, X.J. Effects of wheat varieties on main life parameters of wheat aphids. Chin. J. Appl. Ecol. 1999, 10, 447–451. [Google Scholar]
- Daniels, N.E.; Porter, K.B. Greenbug Resistance Studies in Winter Wheat. J. Econ. Entomol. 1958, 51, 702–704. [Google Scholar] [CrossRef]
- Niraz, S.; Leszczynski, B.; Ciepiela, A.; Urbańska, A. The importance of various plant chemical compounds to constitutive aphid resistance in winter wheat. Rozczniki Nauk. Rol. E. (Ochr. Rosl.) 1987, 17, 61–75. [Google Scholar]
- Hu, X.S.; Liu, X.F.; Thieme, T.; Zhang, G.S.; Liu, T.X.; Zhao, H.Y. Testing the fecundity advantage hypothesis with Sitobion avenae, Rhopalosiphum padi, and Schizaphis graminum (Hemiptera: Aphididae) feeding on ten wheat accessions. Sci. Rep. 2015, 5, 18549. [Google Scholar] [CrossRef] [Green Version]
- Caillaud, C.M.; Dedryver, C.A.; Simon, J.C. Development and reproductive potential of the cereal aphid Sitobion avenae on resistant wheat lines (Triticum monococcum). Ann. Appl. Biol. 1994, 125, 219–232. [Google Scholar] [CrossRef]
- Hong, F.; Han, H.L.; Pu, P.; Wei, D.; Wang, J.; Liu, Y. Effects of Five Host Plant Species on the Life History and Population Growth Parameters of Myzus persicae (Hemiptera: Aphididae). J. Insect Sci. 2019, 19, 15. [Google Scholar] [CrossRef] [Green Version]
- Abbas, N.; Shad, S.A.; Razaq, M. Fitness cost, cross resistance and realized heritability of resistance to imidacloprid in Spodoptera litura (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2012, 103, 181–188. [Google Scholar] [CrossRef]
- Gonzáles, W.L.; Ramirez, C.C.; Olea, N.; Niemeyer, H.M. Host plant changes produced by the aphid Sipha flava: Consequences for aphid feeding behaviour and growth. Entomol. Exp. Appl. 2002, 103, 107–113. [Google Scholar] [CrossRef]
- Golizadeh, A.; Abedi, Z.; Borzoui, E.; Golikhajeh, N.; Jafary, M. Susceptibility of Five Sugar Beet Cultivars to the Black Bean Aphid, Aphis fabae Scopoli (Hemiptera: Aphididae). Neotrop. Entomol. 2016, 45, 427–432. [Google Scholar] [CrossRef]
- Ongaratto, S.; Silveira, C.M.; Santos, M.C.; Gorri, J.E.R.; Sartori, M.M.P.; Hunt, T.E.; Lourencao, A.L.; Baldin, E.L.L. Resistance of Soybean Genotypes to Anticarsia gemmatalis (Lepidoptera: Erebidae): Antixenosis and Antibiosis Characterization. J. Econ. Entomol. 2021, 114, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.Y.; Liu, J.; Duan, X.Y.; Zhang, N.Z.; Gao, F. Effects of different wheat varieties on the development of Sitobion avenae. Chin. J. Appl. Entomol. 2020, 57, 134–141. [Google Scholar] [CrossRef]
- Lan, H.; Zhang, Z.-F.; Wu, J.; Cao, H.-H.; Liu, T.-X. Performance and transcriptomic response of the English grain aphid, Sitobion avenae, feeding on resistant and susceptible wheat cultivars. J. Integr. Agr. 2021, 20, 178–190. [Google Scholar] [CrossRef]
- Rani, P.U.; Pratyusha, S. Defensive role of Gossypium hirsutum L. anti-oxidative enzymes and phenolic acids in response to Spodoptera litura F. feeding. J. Asia Pac. Entomol. 2013, 16, 131–136. [Google Scholar] [CrossRef]
- Haviola, S.; Kapari, L.; Ossipov, V.; Rantala, M.J.; Ruuhola, T.; Haukioja, E. Foliar phenolics are differently associated with Epirrita autumnata growth and immunocompetence. J. Chem. Ecol. 2007, 33, 1013–1023. [Google Scholar] [CrossRef]
- Ruuhola, T.; Salminen, J.P.; Haviola, S.; Yang, S.; Rantala, M.J. Immunological memory of mountain birches: Effects of phenolics on performance of the autumnal moth depend on herbivory history of trees. J. Chem. Ecol. 2007, 33, 1160–1176. [Google Scholar] [CrossRef]
- Wang, Z.; Nur, F.A.; Ma, J.; Wang, J.; Cao, C. Effects of poplar secondary metabolites on performance and detoxification enzyme activity of Lymantria dispar. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 225, 108587. [Google Scholar] [CrossRef]
- Zhu, Y.F.; Chen, M.; Liu, C.Z. Variations of several substances in the seedlings of five oat cultivars infested by aphids. Plant. Prot. 2011, 37, 55–58. [Google Scholar] [CrossRef]
- Green, P.W.; Stevenson, P.C.; Simmonds, M.S.; Sharma, H.C. Phenolic compounds on the pod-surface of pigeonpea, Cajanus cajan, mediate feeding behavior of Helicoverpa armigera larvae. J. Chem. Ecol. 2003, 29, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Piesik, D.; Wenda-Piesik, A. Sitophilus granarius responses to blends of five groups of cereal kernels and one group of plant volatiles. J. Stored Prod. Res. 2015, 63, 63–66. [Google Scholar] [CrossRef]
- Piesik, D.; Pańka, D.; Jeske, M.; Wenda-Piesik, A.; Delaney, K.J.; Weaver, D.K. Volatile induction of infected and neighbouring uninfected plants potentially influence attraction/repellence of a cereal herbivore. J. Appl. Entomol. 2013, 137, 296–309. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Level | AQR | Resistance Type |
|---|---|---|
| 0 | 0 | Immune (I) |
| 1 | 0.01–0.30 | Highly resistant (HR) |
| 2 | 0.31–0.60 | Moderately resistant (MR) |
| 3 | 0.61–0.90 | Low resistant (LR) |
| 4 | 0.91–1.20 | Low susceptible (LS) |
| 5 | 1.21–1.50 | Moderately susceptible (MS) |
| 6 | >1.50 | Highly susceptible (HS) |
| Wheat Varieties | Development Duration/d | Weight Gain/μg | MRGR |
|---|---|---|---|
| Longzhong No.3 | 6.61 ± 0.16 ab | 777.42 ± 26.18 a | 0.418 ± 0.007 ab |
| Huining No.21 | 6.68 ± 0.01 ab | 667.51 ± 17.76 ab | 0.400 ± 0.012 b |
| Ganchun No.29 | 6.54 ± 0.08 ab | 760.89 ± 41.41 ab | 0.423 ± 0.010 ab |
| Longchunza No.2 | 6.19 ± 0.04 b | 749.68 ± 63.07 ab | 0.433 ± 0.017 a |
| Yongliang No.15 | 6.63 ± 0.13 ab | 622.98 ± 102.59 b | 0.395 ± 0.001 b |
| Ganchun No.18 | 6.83 ± 0.40 a | 660.50 ± 13.76 ab | 0.403 ± 0.015 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.-X.; Li, H.-Y.; Quandahor, P.; Gou, Y.-P.; Li, C.-C.; Zhang, Q.-Y.; Haq, I.U.; Ma, Y.; Liu, C.-Z. Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation. Insects 2022, 13, 508. https://doi.org/10.3390/insects13060508
Zhang K-X, Li H-Y, Quandahor P, Gou Y-P, Li C-C, Zhang Q-Y, Haq IU, Ma Y, Liu C-Z. Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation. Insects. 2022; 13(6):508. https://doi.org/10.3390/insects13060508
Chicago/Turabian StyleZhang, Ke-Xin, Hong-Yan Li, Peter Quandahor, Yu-Ping Gou, Chun-Chun Li, Qiang-Yan Zhang, Inzamam Ul Haq, Yue Ma, and Chang-Zhong Liu. 2022. "Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation" Insects 13, no. 6: 508. https://doi.org/10.3390/insects13060508
APA StyleZhang, K.-X., Li, H.-Y., Quandahor, P., Gou, Y.-P., Li, C.-C., Zhang, Q.-Y., Haq, I. U., Ma, Y., & Liu, C.-Z. (2022). Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation. Insects, 13(6), 508. https://doi.org/10.3390/insects13060508