
Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects


Abstract
:Simple Summary
Abstract
1. Introduction
2. Defining Probiotics and Prebiotics: The Bugs That Debug the Bugs
3. The Crosstalk between the Insect and Their Intestinal Microbiota
Probiotics Applications as a Means of Decreasing Disease Occurrence in Mass Reared Insects
4. The Most Common Microorganisms Used as Probiotics in Insects
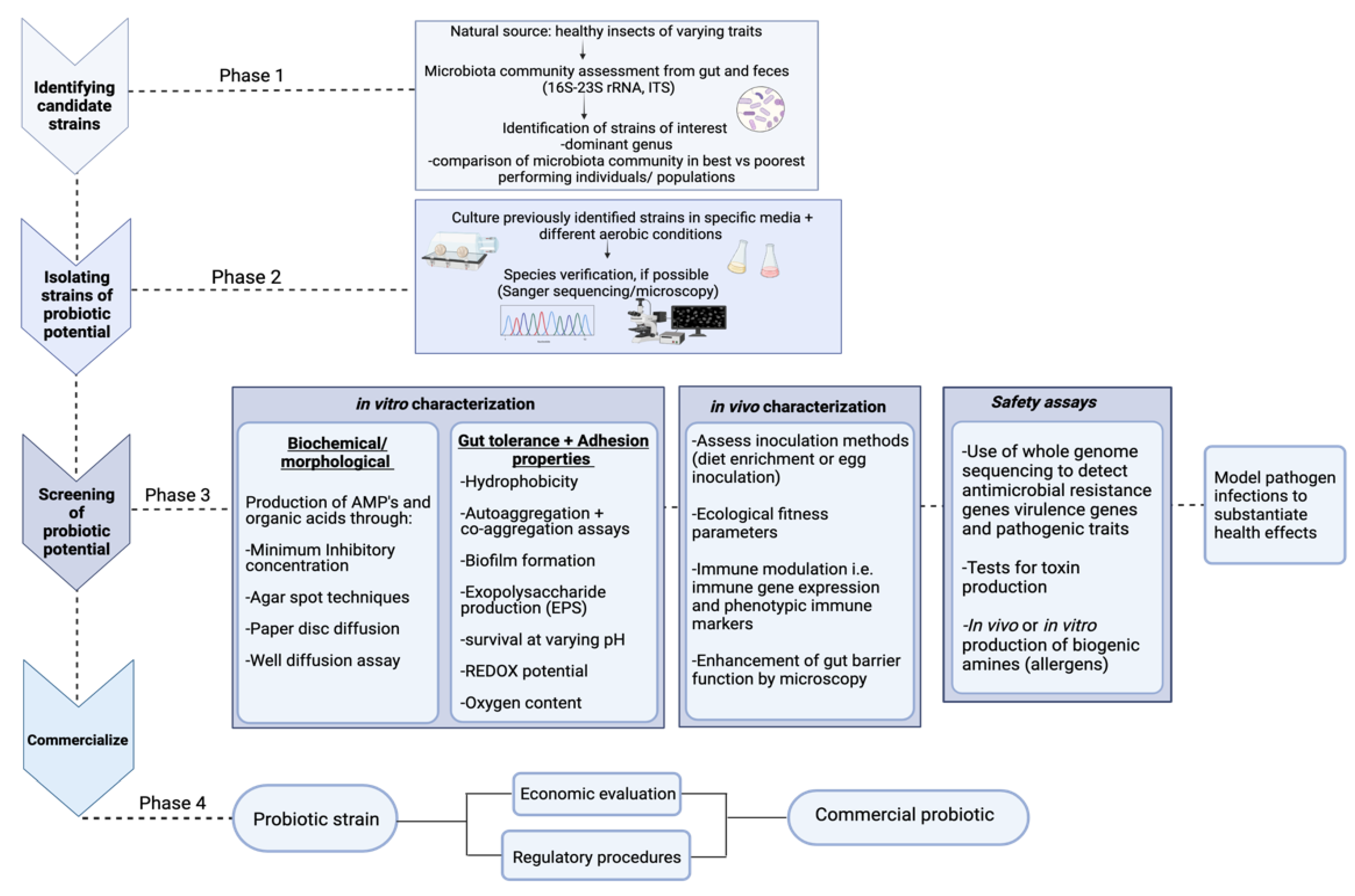
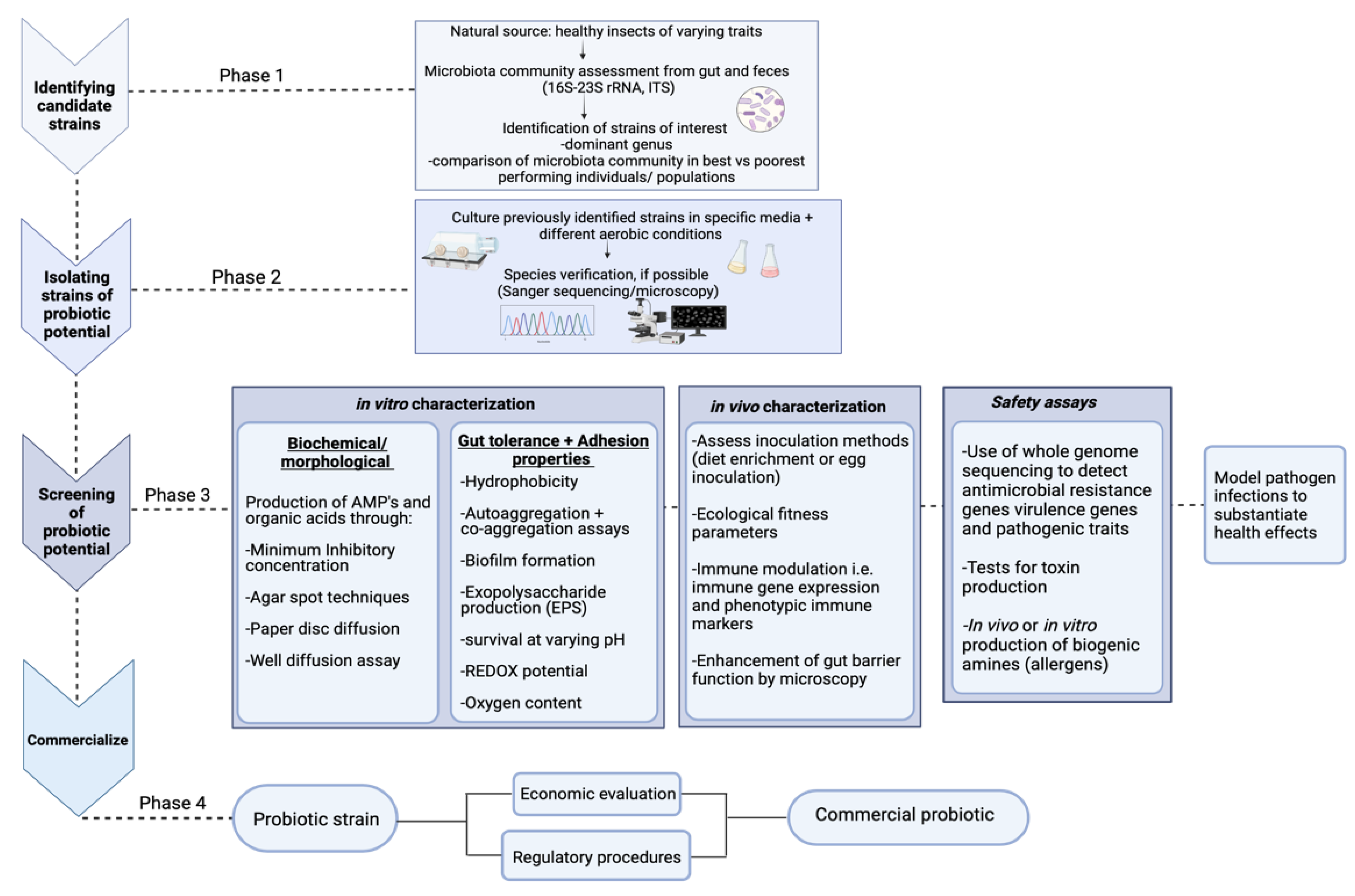
4.1. Isolating Potential Probiotic Strains and Their Characterization
4.1.1. Safety Assays
4.1.2. Analysis of Antimicrobial Potential
4.1.3. Assessing Immune Modulation
4.2. Ecological Fitness Assay
5. Improving Mass Reared Insect Fitness by Probiotic Provision
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edible Insects Market Analysis 2020–2026|Industry Forecasts. Available online: https://www.gminsights.com/industry-analysis/edible-insects-market (accessed on 1 April 2022).
- Alliedmarketresearch Edible Insects Market by Product Type (Whole Insect, Insect Powder, Insect Meal), Insect-Type (Crickets, Black Soldier Fly, Mealworms), Application (Animal Feed, Protein Bar and Shakes, Bakery, Confectionery, Beverages)- Global Forecast to 2030. 2021. Available online: https://www.marketresearch.com/Meticulous-Research-v4061/Edible-Insects-Product-Whole-Insect-13965777/ (accessed on 3 March 2022).
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.C. Ecology of Insect Rearing Systems: A Mini-Review of Insect Rearing Papers from 1906–2017. Adv. Entomol. 2018, 6, 86–115. [Google Scholar] [CrossRef] [Green Version]
- Maciel-Vergara, G.; Jensen, A.B.; Lecocq, A.; Eilenberg, J. Diseases in Edible Insect Rearing Systems. J. Insects Food Feed 2021, 7, 621–638. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Nair, S. Dynamics of Insect–Microbiome Interaction Influence Host and Microbial Symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef]
- Su, W.; Liu, J.; Bai, P.; Ma, B.; Liu, W. Pathogenic Fungi-Induced Susceptibility Is Mitigated by Mutual Lactobacillus Plantarum in the Drosophila Melanogaster Model. BMC Microbiol. 2019, 19, 302. [Google Scholar] [CrossRef] [Green Version]
- Liberti, J.; Engel, P. The Gut Microbiota—Brain Axis of Insects. Curr. Opin. Insect Sci. 2020, 39, 6–13. [Google Scholar] [CrossRef]
- Richard, F.-J. Symbiotic Bacteria Influence the Odor and Mating Preference of Their Hosts. Front. Ecol. Evol. 2017, 5, 143. [Google Scholar] [CrossRef] [Green Version]
- Jordan, H.R.; Tomberlin, J.K. Microbial Influence on Reproduction, Conversion, and Growth of Mass Produced Insects. Curr. Opin. Insect Sci. 2021, 48, 57–63. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E. Probiotics: Definition, Sources, Selection, and Uses. Clin. Infect. Dis. 2008, 46, S58–S61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- György, P.; Norris, R.F.; Rose, C.S. Bifidus Factor. I. A Variant of Lactobacillus Bifidus Requiring a Special Growth Factor. Arch. Biochem. Biophys. 1954, 48, 193–201. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, B.; Viswanath, B.; Sai Gopal, D.V.R. Probiotics as Antiviral Agents in Shrimp Aquaculture. J. Pathog. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hutkins, R.W.; Krumbeck, J.A.; Bindels, L.B.; Cani, P.D.; Fahey, G.; Goh, Y.J.; Hamaker, B.; Martens, E.C.; Mills, D.A.; Rastal, R.A.; et al. Prebiotics: Why Definitions Matter. Curr. Opin. Biotechnol. 2016, 37, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The Gut Microbiota of Insects—Diversity in Structure and Function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Kyritsis, G.A.; Papadopoulos, N.T.; Abd-Alla, A.M.M.; Cáceres, C.; Bourtzis, K. Exploitation of the Medfly Gut Microbiota for the Enhancement of Sterile Insect Technique: Use of Enterobacter Sp. in Larval Diet-Based Probiotic Applications. PLoS ONE 2015, 10, e0136459. [Google Scholar] [CrossRef] [Green Version]
- Somerville, J.; Zhou, L.; Raymond, B. Aseptic Rearing and Infection with Gut Bacteria Improve the Fitness of Transgenic Diamondback Moth, Plutella Xylostella. Insects 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Contreras, M.; Vlisidou, I. The Diversity of Insect-Bacteria Interactions and Its Applications for Disease Control. Biotechnol. Genet. Eng. Rev. 2008, 25, 203–244. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.-L.; Zhang, S.-X.; Chen, Z.-H.; Tao, H.; Li, X.; Qiu, J.-F.; Cui, W.-Z.; Sima, Y.-H.; Cui, W.-Z.; Xu, S.-Q. Differences in Gut Microbiota between Silkworms (Bombyx Mori) Reared on Fresh Mulberry (Morus Alba Var. Multicaulis) Leaves or an Artificial Diet. RSC Adv. 2018, 8, 26188–26200. [Google Scholar] [CrossRef] [Green Version]
- Ojha, A.; Sinha, D.K.; Padmakumari, A.P.; Bentur, J.S.; Nair, S. Bacterial Community Structure in the Asian Rice Gall Midge Reveals a Varied Microbiome Rich in Proteobacteria. Sci. Rep. 2017, 7, 9424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Xiao, G.; Zhou, W.; Gao, Y.; Li, Z.; Du, G.; Chen, B. Midgut Microbiota Diversity of Potato Tuber Moth Associated with Potato Tissue Consumed. BMC Microbiol. 2020, 20, 58. [Google Scholar] [CrossRef] [Green Version]
- Weiss, B.L.; Maltz, M.A.; Vigneron, A.; Wu, Y.; Walter, K.S.; O’Neill, M.B.; Wang, J.; Aksoy, S. Colonization of the Tsetse Fly Midgut with Commensal Kosakonia Cowanii Zambiae Inhibits Trypanosome Infection Establishment. PLOS Pathog. 2019, 15, e1007470. [Google Scholar] [CrossRef] [Green Version]
- Paniagua Voirol, L.R.; Frago, E.; Kaltenpoth, M.; Hilker, M.; Fatouros, N.E. Bacterial Symbionts in Lepidoptera: Their Diversity, Transmission, and Impact on the Host. Front. Microbiol. 2018, 9, 556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jiao, S.; Li, X.; Li, M. Bacterial and Fungal Gut Communities of Agrilus Mali at Different Developmental Stages and Fed Different Diets. Sci. Rep. 2018, 8, 15634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egert, M.; Stingl, U.; Dyhrberg Bruun, L.; Pommerenke, B.; Brune, A.; Friedrich, M.W. Structure and Topology of Microbial Communities in the Major Gut Compartments of Melolontha Melolontha Larvae (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 2005, 71, 4556–4566. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Teh, B.-S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and Activity of the Gut Microbiota across the Life History of the Insect Herbivore Spodoptera Littoralis. Sci. Rep. 2016, 6, 29505. [Google Scholar] [CrossRef]
- Mazumdar, T.; Teh, B.S.; Murali, A.; Schmidt-Heck, W.; Schlenker, Y.; Vogel, H.; Boland, W. Survival Strategies of Enterococcus Mundtii in the Gut of Spodoptera Littoralis: A Live Report; Microbiology. J. Chem. Ecol. 2020, 47, 227–241. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. T HE G UT B ACTERIA OF I NSECTS: Nonpathogenic Interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Engel, P. Mechanisms Underlying Gut Microbiota–Host Interactions in Insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef] [PubMed]
- Gandotra, S.; Bhuyan, P.M.; Gogoi, D.K.; Kumar, A.; Subramanian, S. Screening of Nutritionally Important Gut Bacteria from the Lepidopteran Insects Through Qualitative Enzyme Assays. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 329–337. [Google Scholar] [CrossRef]
- Pauchet, Y.; Heckel, D.G. The Genome of the Mustard Leaf Beetle Encodes Two Active Xylanases Originally Acquired from Bacteria through Horizontal Gene Transfer. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131021. [Google Scholar] [CrossRef]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and Evolution of Heritable Bacterial Symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Nutritional Interactions in Insect-Microbial Symbioses: Aphids and Their Symbiotic Bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Shuttleworth, L.A.; Khan, M.A.M.; Osborne, T.; Collins, D.; Srivastava, M.; Reynolds, O.L. A Walk on the Wild Side: Gut Bacteria Fed to Mass-Reared Larvae of Queensland Fruit Fly [Bactrocera Tryoni (Froggatt)] Influence Development. BMC Biotechnol. 2019, 19, 95. [Google Scholar] [CrossRef]
- van den Bosch, T.J.M.; Welte, C.U. Detoxifying Symbionts in Agriculturally Important Pest Insects. Microb. Biotechnol. 2017, 10, 531–540. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut Microbiota Mediate Insecticide Resistance in the Diamondback Moth, Plutella Xylostella (L.). Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Tago, K.; Hayatsu, M.; Kikuchi, Y. Detoxifying Symbiosis: Microbe-Mediated Detoxification of Phytotoxins and Pesticides in Insects. Nat. Prod. Rep. 2018, 35, 434–454. [Google Scholar] [CrossRef]
- Genta, F.A.; Dillon, R.J.; Terra, W.R.; Ferreira, C. Potential Role for Gut Microbiota in Cell Wall Digestion and Glucoside Detoxification in Tenebrio Molitor Larvae. J. Insect Physiol. 2006, 52, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, F.H.; Gendrin, M.; Wyer, C.A.S.; Christophides, G.K. Microbiota-Induced Peritrophic Matrix Regulates Midgut Homeostasis and Prevents Systemic Infection of Malaria Vector Mosquitoes. PLOS Pathog. 2017, 13, e1006391. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in Insects Produced for Food and Feed. J. Insects Food Feed 2015, 1, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Sikorowski, P.P.; Lawrence, A.M. Microbial Contamination and Insect Rearing. Am. Entomol. 1994, 40, 240–253. [Google Scholar] [CrossRef]
- Muhammad, A.; Habineza, P.; Ji, T.; Hou, Y.; Shi, Z. Intestinal Microbiota Confer Protection by Priming the Immune System of Red Palm Weevil Rhynchophorus Ferrugineus Olivier (Coleoptera: Dryophthoridae). Front. Physiol. 2019, 10, 1303. [Google Scholar] [CrossRef]
- Guarner, F.; Sanders, M.E.; Eliakim, R.; Garisch, J.; Gangl, A.; Thomson, A.; Kim, N. Probiotics and Prebiotics. World Gastroenterology Organization Probiotics and Prebiotics Guidelines. 2017. Available online: https://www.worldgastroenterology.org/guidelines/probiotics-and-prebiotics/probiotics-and-prebiotics-english (accessed on 4 January 2022).
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Nishida, S.; Ishii, M.; Nishiyama, Y.; Abe, S.; Ono, Y.; Sekimizu, K. Lactobacillus Paraplantarum 11-1 Isolated from Rice Bran Pickles Activated Innate Immunity and Improved Survival in a Silkworm Bacterial Infection Model. Front. Microbiol. 2017, 8, 436. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; de Barros, P.P.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O.C. Lactobacillus Rhamnosus Inhibits Candida Albicans Virulence Factors in Vitro and Modulates Immune System in Galleria Mellonella. J. Appl. Microbiol. 2017, 122, 201–211. [Google Scholar] [CrossRef]
- Jorjão, A.L.; de Oliveira, F.E.; Leão, M.V.P.; Jorge, A.O.C.; de Oliveira, L.D. Effect of Lactobacillus Rhamnosus on the Response of Galleria Mellonella against Staphylococcus Aureus and Escherichia Coli Infections. Arch. Microbiol. 2018, 200, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, T.M.; Serra, C.R.; La Ragione, R.M.; Woodward, M.J.; Henriques, A.O. Screening for Bacillus Isolates in the Broiler Gastrointestinal Tract. Appl. Environ. Microbiol. 2005, 71, 968–978. [Google Scholar] [CrossRef] [Green Version]
- Burnham, A.J. Scientific Advances in Controlling Nosema Ceranae (Microsporidia) Infections in Honey Bees (Apis Mellifera). Front. Vet. Sci. 2019, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ptaszyńska, A.A.; Paleolog, J.; Borsuk, G. Nosema Ceranae Infection Promotes Proliferation of Yeasts in Honey Bee Intestines. PLoS ONE 2016, 11, e0164477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruščáková, I.C.; Schusterová, P.; Bielik, B.; Toporčák, J.; Bíliková, K.; Mudroňová, D. Effect of Application of Probiotic Pollen Suspension on Immune Response and Gut Microbiota of Honey Bees (Apis Mellifera). Probiotics Antimicrob. Proteins 2020, 12, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.D.; dos Santos Velloso, M.; Figueiredo, L.M.A.; Martins, C.P.; Jorge, A.O.C.; Junqueira, J.C. Clinical Strains of Lactobacillus Reduce the Filamentation of Candida Albicans and Protect Galleria Mellonella against Experimental Candidiasis. Folia Microbiol. 2018, 63, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grau, T.; Vilcinskas, A.; Joop, G. Probiotic Enterococcus Mundtii Isolate Protects the Model Insect Tribolium Castaneum against Bacillus Thuringiensis. Front. Microbiol. 2017, 8, 1261. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Ros, V.I.D. Viruses of Insects Reared for Food and Feed. J. Invertebr. Pathol. 2017, 147, 60–75. [Google Scholar] [CrossRef]
- Jung, J.; Heo, A.; Park, Y.W.; Kim, Y.J.; Koh, H.; Park, W. Gut Microbiota of Tenebrio Molitor and Their Response to Environmental Change. J. Microbiol. Biotechnol. 2014, 24, 888–897. [Google Scholar] [CrossRef]
- Kamei, Y.; Yoshimizu, M.; Ezura, Y.; Kimura, T. Screening of Bacteria with Antiviral Activity from Fresh Water Salmonid Hatcheries. Microbiol. Immunol. 1988, 32, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, F.; Teixeira, N.; Rigottier-Gois, L.; Marujo, P.; Nielsen-LeRoux, C.; Crespo, M.T.B.; Lopes, M.d.F.S.; Serror, V. Virulence of Enterococcus Faecalis Dairy Strains in an Insect Model: The Role of FsrB and GelE. Microbiology 2009, 155, 3564–3571. [Google Scholar] [CrossRef] [Green Version]
- Al Atya, A.K.; Drider-Hadiouche, K.; Ravallec, R.; Silvain, A.; Vachee, A.; Drider, D. Probiotic Potential of Enterococcus Faecalis Strains Isolated from Meconium. Front. Microbiol. 2015, 6, 227. [Google Scholar] [CrossRef]
- La Fauce, K.; Owens, L. Suppression of Penaeus Merguiensis Densovirus Following Oral Delivery of Live Bacteria Expressing DsRNA in the House Cricket (Acheta Domesticus) Model. J. Invertebr. Pathol. 2013, 112, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Ng’ang’a, J.; Imathiu, S.; Fombong, F.; Borremans, A.; Van Campenhout, L.; Broeck, J.V.; Kinyuru, J. Can Farm Weeds Improve the Growth and Microbiological Quality of Crickets (Gryllus Bimaculatus)? J. Insects Food Feed 2020, 6, 199–209. [Google Scholar] [CrossRef]
- Taha, R.H.; Kamel, H.M. Micro-Organisms Supplementation to Mulberry Silkworm, Bombyx Mori, L. Egypt. Acad. J. Biol. Sci. Entomol. 2017, 10, 57–64. [Google Scholar] [CrossRef]
- Suraporn, S.; Sangsuk, W.; Chanhan, P.; Promma, S. Effects of Probiotic Bacteria on the Growth Parameters of the Thai Silkworm, Bombyxmori. Thai J. Agric. Sci. 2015, 48, 29–33. [Google Scholar]
- Suraporn, S.; Terenius, O. Supplementation of Lactobacillus Casei Reduces the Mortality of Bombyx Mori Larvae Challenged by Nosema Bombycis. BMC Res. Notes 2021, 14, 398. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Chauhan, R.M.; Pande, A.B.; Gokhale, S.B.; Hegde, N.G. Effect of Use of Lactobacillus Plantarumas a Probiotics to Improve Cocoon Production of Mulberry Silkworm, Bombyxmori (L.). J. Basic Appl. Sci. 2005, 1, 1–8. [Google Scholar]
- Saranya, M.; Krishnamoorthy, S.; Murugesh, K. Fortification of Mulberry Leaves with Indigenous Probiotic Bacteria on Larval Growth and Economic Traits of Silkworm (Bombyx Mori, L.). J. Entomol. Zool. Stud. 2019, 7, 780–784. [Google Scholar]
- Esaivani, C.; Vasanthi, K.; Bharathi, R.; Chairman, K. Impact of Probiotic Saccharomyces Cerevisiae on the Enzymatic Profile and the Economic Parameters of Sylkworm Bombyx Mori, L. Adv. Biol. Biomed. 2014, 1, 1–7. [Google Scholar]
- Alcosaba, D.M. Effect of Fungus, Trichoderma Harzianumas Probiotic on the Growth, Cocoon Parameters, Silk Characters and Resistance of Silkworms (Bombyx Mori) Challenged by Muscardine Disease-Causing Metarhizium. Ascendens Asia J. Multidiscip. Res. Abstr. 2019, 3. [Google Scholar]
- Rajakumari, D.V.S.; Padmalatha, C.; Das, S.S.M.; Ranjitsingh, A.J.A. Efficacy of Probiotic and Neutraceutical Feed Supplements against Flacherie Disease in Mulberry Silkworm, Bombyx Mori, L. Indian J. Sericult. 2007, 46, 179–182. [Google Scholar]
- Scalfaro, C.; Iacobino, A.; Nardis, C.; Franciosa, G. Galleria Mellonella as an in Vivo Model for Assessing the Protective Activity of Probiotics against Gastrointestinal Bacterial Pathogens. FEMS Microbiol. Lett. 2017, 364, fnx225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela, S.F.; Barbosa, J.O.; Rossoni, R.D.; Santos, J.D.; Prata, M.C.; Anbinder, A.L.; Jorge, A.O.; Junqueira, J.C. Lactobacillus Acidophilus ATCC 4356 Inhibits Biofilm Formation by C. Albicansand Attenuates the Experimental Candidiasis in Galleria Mellonella. Virulence 2015, 6, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berríos, P.; Fuentes, J.A.; Salas, D.; Carreño, A.; Aldea, P.; Fernández, F.; Trombert, A.N. Inhibitory Effect of Biofilm-Forming Lactobacillus Kunkeei Strains against Virulent Pseudomonas Aeruginosa in Vitro and in Honeycomb Moth (Galleria Mellonella) Infection Model. Benef. Microbes 2018, 9, 257–268. [Google Scholar] [CrossRef]
- Rizou, E.; Kalogiouri, N.; Bisba, M.; Papadimitriou, A.; Kyrila, G.; Lazou, A.; Andreadis, S.; Hatzikamari, M.; Mourtzinos, I.; Touraki, M. Amelioration of Growth, Nutritional Value, and Microbial Load of Tenebrio Molitor (Coleoptera: Tenebrionidae) through Probiotic Supplemented Feed. Eur. Food Res. Technol. 2022, 248, 727–739. [Google Scholar] [CrossRef]
- Lecocq, A.; Natsopoulou, M.E.; Berggreen, I.E.; Eilenberg, J.; Heckmann, L.-H.L.; Nielsen, H.V.; Stensvold, C.R.; Jensen, A.B. Probiotic Properties of an Indigenous Pediococcus Pentosaceus Strain on Tenebrio Molitor Larval Growth and Survival. J. Insects Food Feed 2021, 7, 975–986. [Google Scholar] [CrossRef]
- Kooienga, E.M.; Baugher, C.; Currin, M.; Tomberlin, J.K.; Jordan, H.R. Effects of Bacterial Supplementation on Black Soldier Fly Growth and Development at Benchtop and Industrial Scale. Front. Microbiol. 2020, 11, 587979. [Google Scholar] [CrossRef]
- Yu, G.; Cheng, P.; Chen, Y.; Li, Y.; Yang, Z.; Chen, Y.; Tomberlin, J.K. Inoculating Poultry Manure With Companion Bacteria Influences Growth and Development of Black Soldier Fly (Diptera: Stratiomyidae) Larvae. Environ. Entomol. 2011, 40, 30–35. [Google Scholar] [CrossRef]
- Franks, K.; Kooienga, E.; Sanders, M.; Pendarvis, K.; Yang, F.; Tomberlin, J.K.; Jordan, H.R. The Effect of Rhodococcus Rhodochrous Supplementation on Black Soldier Fly (Diptera: Stratiomyidae) Development, Nutrition, and Waste Conversion. J. Insects Food Feed 2021, 7, 397–408. [Google Scholar] [CrossRef]
- Richard, N.; Rabot, R.; Beutin, C.; van Loon, J.J.A. Live Yeast Probiotic Can Boost Growth Performances of Yellow Mealworm and Black Soldier Fly Larvae; Wageningen University: Wageningen, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, S.; Zhang, X.; Zhang, K.; Liu, W.; Zhang, R.; Zhang, Z. Enterobacter Hormaechei in the Intestines of Housefly Larvae Promotes Host Growth by Inhibiting Harmful Intestinal Bacteria. Parasit. Vectors 2021, 14, 598. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and Immune Health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Yao, Z.; Li, Y.; Xi, Z.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H. Intestinal Probiotics Restore the Ecological Fitness Decline of Bactrocera Dorsalis by Irradiation. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef]
- Dierking, K.; Pita, L. Receptors Mediating Host-Microbiota Communication in the Metaorganism: The Invertebrate Perspective. Front. Immunol. 2020, 11, 1251. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E. Impact of 16S RRNA Gene Sequence Analysis for Identification of Bacteria on Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winand, R.; Bogaerts, B.; Hoffman, S.; Lefevre, L.; Delvoye, M.; Van Braekel, J.; Fu, Q.; Roosens, N.H.; De Keersmaecker, S.C.; Vanneste, K. Targeting the 16S RRNA Gene for Bacterial Identification in Complex Mixed Samples: Comparative Evaluation of Second (Illumina) and Third (Oxford Nanopore Technologies) Generation Sequencing Technologies. Int. J. Mol. Sci. 2019, 21, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeruva, T.; Vankadara, S.; Ramasamy, S.; Lingaiah, K. Identification of Potential Probiotics in the Midgut of Mulberry Silkworm, Bombyx Mori Through Metagenomic Approach. Probiotics Antimicrob. Proteins 2020, 12, 635–640. [Google Scholar] [CrossRef]
- World Health Organization. Food and Agriculture Organization of the United Nations Probiotics in Food, Health Nutritional Proprieties and Guidelines for Evaluation; FAO/WHO: Geneva, Switzerland, 2016; ISBN 02544725. [Google Scholar]
- Papadimitriou, K.; Zoumpopoulou, G.; FolignÃ, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering Probiotic Microorganisms: In Vitro, in Vivo, Genetic and Omics Approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byakika, S.; Mukisa, I.M.; Byaruhanga, Y.B.; Muyanja, C. A Review of Criteria and Methods for Evaluating the Probiotic Potential of Microorganisms. Food Rev. Int. 2019, 35, 427–466. [Google Scholar] [CrossRef]
- Kumura, H.; Tanoue, Y.; Tsukahara, M.; Tanaka, T.; Shimazaki, K. Screening of Dairy Yeast Strains for Probiotic Applications. J. Dairy Sci. 2004, 87, 4050–4056. [Google Scholar] [CrossRef]
- Chomchai, S.; Chomchai, C. Histamine Poisoning from Insect Consumption: An Outbreak Investigation from Thailand. Clin. Toxicol. 2018, 56, 126–131. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740.
- Tagg, J.; McGiven, A. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef] [PubMed]
- Cabo, M.L.; Murado, M.A.; Gonzalez, M.P.; Pastoriza, L. A Method for Bacteriocin Quantification. J. Appl. Microbiol. 1999, 87, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Microbial Production of Bacteriocins: Latest Research Development and Applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of Culture Media, PH and Temperature on Growth and Bacteriocin Production of Bacteriocinogenic Lactic Acid Bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Measurement of Aggregation Properties between Probiotics and Pathogens: In Vitro Evaluation of Different Methods. J. Microbiol. Methods 2007, 71, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Ahn, J. Probiotic-Mediated Competition, Exclusion and Displacement in Biofilm Formation by Food-Borne Pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef]
- Hamden, H.; MSaad Guerfali, M.; Charaabi, K.; Djobbi, W.; Fadhl, S.; Mahjoubi, M.; Mnasri, K.; Najjari, A.; Saidi, M.; Chevrier, C.; et al. Screening and Selection of Potential Probiotic Strains from the Mediterranean Fruit Fly (Ceratitis Capitata) Guts Origin Based on SIT Application. J. Radiat. Res. Appl. Sci. 2020, 13, 776–791. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and Aggregation Properties of Probiotic and Pathogen Strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- De Gregorio, E.; Han, S.-J.; Lee, W.-J.; Baek, M.-J.; Osaki, T.; Kawabata, S.-I.; Lee, B.-L.; Iwanaga, S.; Lemaitre, B.; Brey, P.T. An Immune-Responsive Serpin Regulates the Melanization Cascade in Drosophila. Dev. Cell 2002, 3, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Tlak Gajger, I.; Vlainić, J.; Šoštarić, P.; Prešern, J.; Bubnič, J.; Smodiš Škerl, M.I. Effects on Some Therapeutical, Biochemical, and Immunological Parameters of Honey Bee (Apis Mellifera) Exposed to Probiotic Treatments, in Field and Laboratory Conditions. Insects 2020, 11, 638. [Google Scholar] [CrossRef]
- Peacock, K.A. The Three Faces of Ecological Fitness. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2011, 42, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.; Bouchard, F. Matthen and Ariew’s Obituary for Fitness: Reports of Its Death Have Been Greatly Exaggerated. Biol. Philos. 2005, 20, 343–353. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Chapman, T.A.; Shuttleworth, L.A.; Riegler, M.; Reynolds, O.L. Tephritid-Microbial Interactions to Enhance Fruit Fly Performance in Sterile Insect Technique Programs. BMC Microbiol. 2019, 19, 287. [Google Scholar] [CrossRef] [PubMed]
- Kyritsis, G.A.; Augustinos, A.A.; Cáceres, C.; Bourtzis, K. Medfly Gut Microbiota and Enhancement of the Sterile Insect Technique: Similarities and Differences of Klebsiella Oxytoca and Enterobacter Sp. AA26 Probiotics during the Larval and Adult Stages of the VIENNA 8D53+ Genetic Sexing Strain. Front. Microbiol. 2017, 8, 2064. [Google Scholar] [CrossRef] [PubMed]
- Gavriel, S.; Jurkevitch, E.; Gazit, Y.; Yuval, B. Bacterially Enriched Diet Improves Sexual Performance of Sterile Male Mediterranean Fruit Flies: Bacteria and Medfly Sexual Performance. J. Appl. Entomol. 2011, 135, 564–573. [Google Scholar] [CrossRef]
- Daisley, B.A.; Pitek, A.P.; Chmiel, J.A.; Al, K.F.; Chernyshova, A.M.; Faragalla, K.M.; Burton, J.P.; Thompson, G.J.; Reid, G. Novel Probiotic Approach to Counter Paenibacillus Larvae Infection in Honey Bees. ISME J. 2020, 14, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P. Antibacterial Particles and Predatory Bacteria as Alternatives to Antibacterial Chemicals in the Era of Antibiotic Resistance. Curr. Opin. Microbiol. 2021, 64, 109–116. [Google Scholar] [CrossRef]
- Consuegra, J.; Grenier, T.; Akherraz, H.; Rahioui, I.; Gervais, H.; da Silva, P.; Leulier, F. Metabolic Cooperation among Commensal Bacteria Supports Drosophila Juvenile Growth under Nutritional Stress. iScience 2020, 23, 101232. [Google Scholar] [CrossRef]
- Zhong, J.; Zhang, F.; Peng, Y.; Ji, Z.; Li, H.; Li, S.; Zhang, X.; Shi, Q.; Zhang, J. Mixed Culture of Probiotics on a Solid-State Medium: An Efficient Method to Produce an Affordable Probiotic Feed Additive. Biotechnol. Bioprocess. Eng. 2017, 22, 758–766. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Insect Species | Probiotics | Effects on Performance and Yield | Ref. |
|---|---|---|---|
| Silkworm | Bacteria | ||
| Bombyx mori | Bifidobacteria | ||
| Bifidobacterium bifidum | Found to be an immunomodulating agent (increase in the activity of protease, amylase and invertase); increased raw silk production with fewer cocoons | [66] | |
| Lactobacilli | |||
| Lactobacillus acidophilus | Stimulated growth factors leading to an increase in the silk yield and to an improvement of the silk harvest | [67] | |
| L. casei | Improved larval weight, cocooning ratio, pupation ratio, and economic characters (cocoon weight and size) when larvae were infected with microsporidium Nosema bombycis | [68] | |
| L. plantarum | Helped to increase body weight, cocoon, shell, and pupation rate | [69] | |
| Staphylococci | |||
| Staphylococcus gallinarum strain SWGB 7 & S. arlettae strain SWGB 16 | Increased larval growth and cocoon characters (filament length and weight, finer denier) | [70] | |
| Yeast | |||
| Saccharomyces cerevisiae | Immunomodulating agent; increased raw silk production with fewer cocoons; increased protein content | [66,71] | |
| Fungi | |||
| Trichoderma harzianumas | Improved food digestion leading to increased growth and resistance to mortality by Metarhizium anisopliae and Beauveria bassiana | [72] | |
| Commercial products | |||
| Lact-Act a | Larvae reared on leaves sprayed with Lact-Act had increased survival when exposed to bacterial pathogens (Bacillus thuringiensis var. sotto. and Staphylococcus aureus) | [73] | |
| Insect species | Probiotics | Effects on performance and yield | Ref. |
| Greater wax moth | Bacteria | ||
| Galleria mellonella | Clostridiaceae | ||
| Clostridium butyricum Miyairi 588 | Induced immune response and increased survival rates against Salmonella enterica serovar Typhimurium, enteropathogenic Escherichia coli or Listeria monocytogenes. | [74] | |
| Lactobacilli | |||
| Lactobacillus acidophilus ATCC 4356 | Increased survival from Candida albicans infection | [75] | |
| L. kunkeei b | Reduces infection of Pseudomonas aeruginosa through biofilm formation and affecting their stability | [76] | |
| L. rhamnosus ATCC 7469 | Promoted greater protection in larvae infected with Staphylococcus aureus or Escherichia coli. | [52] | |
| L. rhamnosus ATCC 9595 | Reduces infection of Pseudomonas aeruginosa through biofilm formation and affecting their stability | [51] | |
| L. rhamnosus GG | Induced immune response and increased survival rates against Salmonella enterica serovar Typhimurium, enteropathogenic Escherichia coli or Listeria monocytogenes. | [74] | |
| Yellow mealworm | Bacteria | ||
| Tenebrio molitor | Bacilli | ||
| Bacillus subtilis | Enhanced growth and nutritional fortification | [77] | |
| B. toyonensis | Enhanced growth and increased dry matter weight of produced feed | [77] | |
| Enterococcaceae | |||
| Enterococcus faecalis | Increased larval weight gain and overall size and shorter time to pupation, also increased the crude protein content | [77] | |
| Lactobacilli | |||
| Pediococcus pentosaceus (Isolated from the gut of Tenebrio larvae) | Reduces mortality in larvae and accelerates the rate of development. The strain has antimicrobial activity towards a number of pathogenic bacteria including several Bacillus thuringiensis, Serratia, and Pseudomonas spp. | [78] | |
| Insect species | Probiotics | Effects on performance and yield | Ref. |
| Black soldier fly | Bacteria | ||
| Hermetia illucens | Actinomycetia | ||
| Arthrobacter AK19 | Enhanced growth rate at early life stages culminating in larger larvae than control | [79] | |
| Bacilli | |||
| Bacillus subtilis S15 S16 S19; B. subtilis natto D1 | Increased larval weight and total development time compared to control larvae | [80] | |
| Bifidobacteria | |||
| Bifidobacterium breve | Larvae had lower weights and appeared weak/slow/discolored compared to control | [79] | |
| Nocardiaceae | |||
| Rhodococcus rhodochrous | Increased conversion rate, which could result in larger larvae with less feed. Larvae had increased proteins content related to energy production and storage. Larvae without the probiotic which had higher content of proteins related to stress responses. | [81] | |
| Commercial product | |||
| Actisaf® Sc47 c | Increased bioconversion rate, lipid and protein yield in processed larvae | [82] | |
| House fly | Bacteria | ||
| Musca domestica | Enterobacteriaceae | ||
| Enterobacter hormaechei | Increased body length and weight, pupal weight, and shortened growth cycle, which is a considerable advantage that can contribute to cost savings and boost production in large-scale feeding facilities. | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savio, C.; Mugo-Kamiri, L.; Upfold, J.K. Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects. Insects 2022, 13, 376. https://doi.org/10.3390/insects13040376
Savio C, Mugo-Kamiri L, Upfold JK. Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects. Insects. 2022; 13(4):376. https://doi.org/10.3390/insects13040376
Chicago/Turabian StyleSavio, Carlotta, Loretta Mugo-Kamiri, and Jennifer K. Upfold. 2022. "Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects" Insects 13, no. 4: 376. https://doi.org/10.3390/insects13040376
APA StyleSavio, C., Mugo-Kamiri, L., & Upfold, J. K. (2022). Bugs in Bugs: The Role of Probiotics and Prebiotics in Maintenance of Health in Mass-Reared Insects. Insects, 13(4), 376. https://doi.org/10.3390/insects13040376