
C/EBPα Regulates PxTreh1 and PxTreh2 Trehalase-Related Bt Resistance in Plutella xylostella (L.)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Cell Line
2.2. Trehalase Activity Assay
2.3. Quantitative Real-Time PCR
2.4. 5’ RNA Ligase-Mediated Rapid Amplification of cDNA Ends (5’ RLM-RACE)
2.5. Cloning of the Promoter and TFs
2.6. PxTreh Promoter-Reporter Constructs
2.7. Dual-Luciferase Activity Assay
2.8. Y1H Assay
2.9. dsRNA Synthesis and RNAi
2.10. Statistical Analysis
3. Results
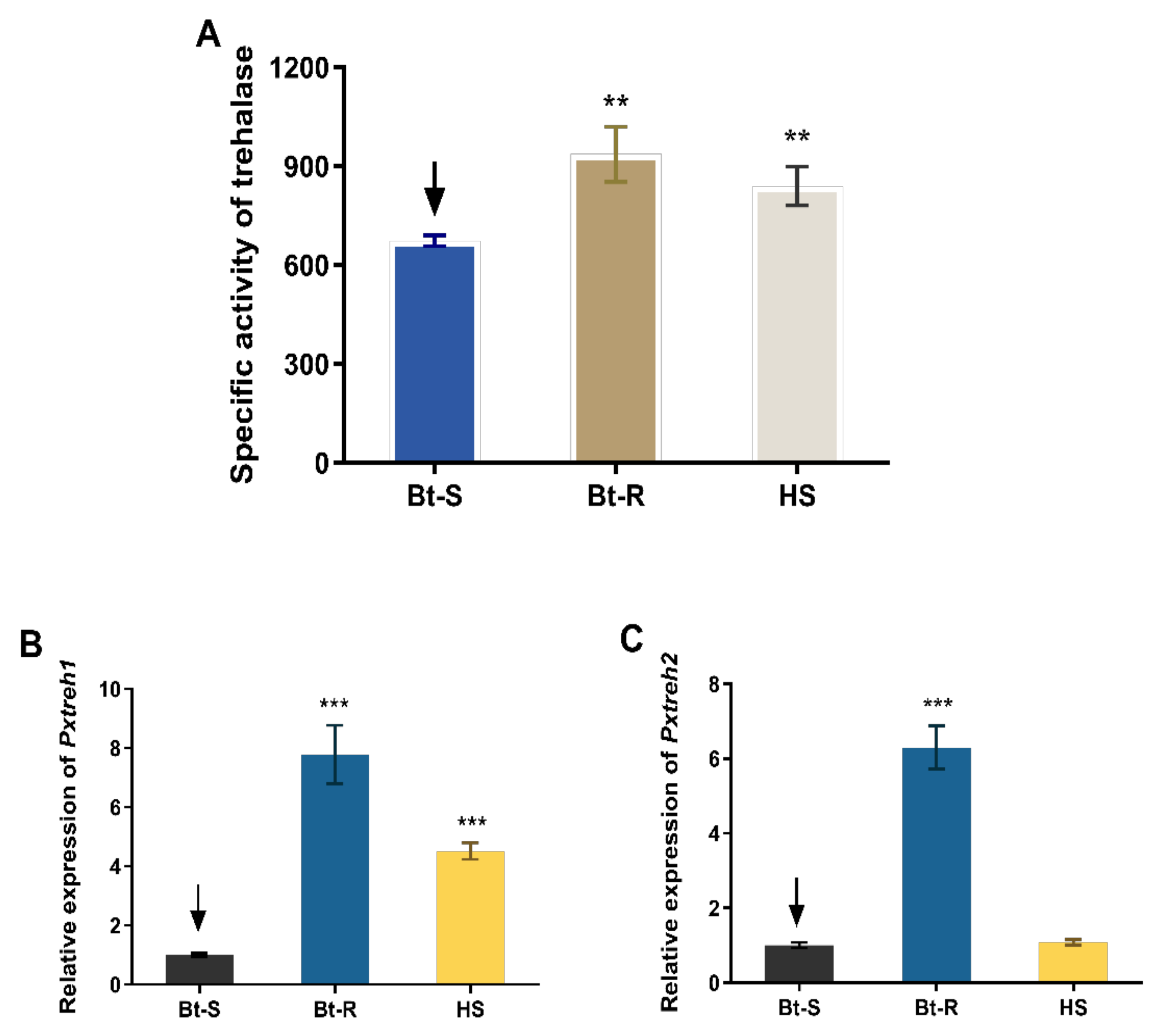
3.1. Trehalase Activity Was Significantly High in Bt-R P. xylostella Larvae
3.2. Expression of PxTreh1 and PxTreh2 Genes Were Significantly High in the HS and Bt-R P. xylostella Population
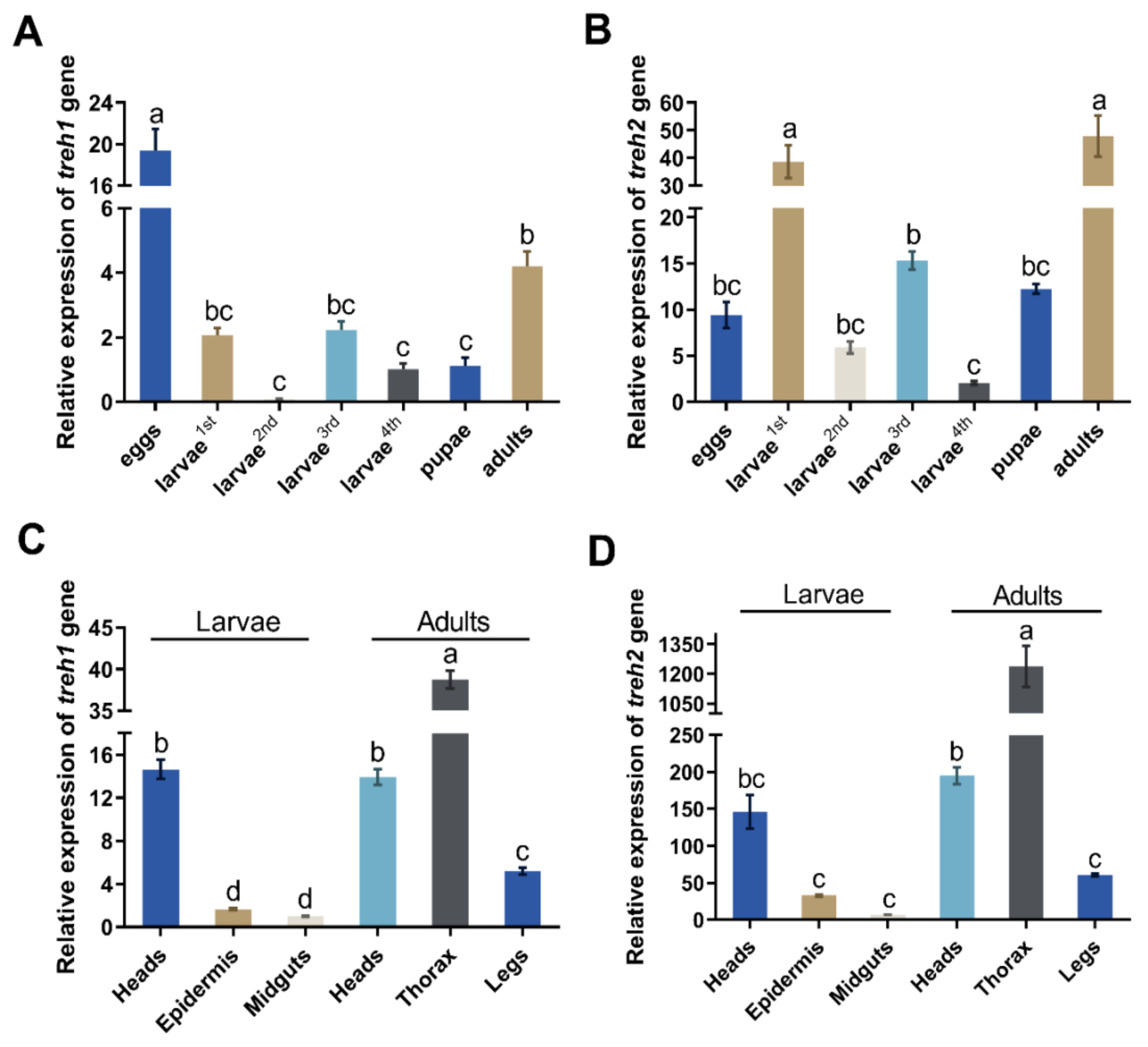
3.3. The Expression of PxTreh Gene in P. xylostella Developmental Stages
3.4. Cloning of the PxTreh Promoter
3.5. The Promoter Activity of the PxTreh Gene
3.6. The C/EBPα Transcription Factor Enhances PxTreh Expression
3.7. C/EBPα Interacts with the PxTreh2 Promoter Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, A.; Schlöder, P.; Steele, J.E.; Wegener, G. The regulation of trehalose metabolism in insects. Experientia 1996, 52, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Chen, J.; Yao, Q.; Pan, Z.Q.; Xu, W.H.; Wang, S.G.; Zhang, W.Q. Characterization of a trehalose-6-phosphate synthase gene from Spodoptera exigua and its function identification through RNA interference. J. Insect Physiol. 2010, 56, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2015, 25, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Wei, P.; Zhao, L.; Shi, Z.K.; Shen, Q.D.; Yang, M.M.; Xie, G.Q.; Wang, S.G. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathway in Tribolium castaneum. BMC Biotechnol. 2016, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wegener, G.; Tschiedel, V.; Schlöder, P.; Ando, O. The toxic and lethal effects of the trehalase inhibitor trehazolin in locusts are caused by hypoglycaemia. J. Exp. Biol. 2003, 206, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.C.P.; Ribeiro, A.F.; Terra, W.R.; Ferreira, C. Sequencing of Spodoptera frugiperda midgut trehalases and demonstration of secretion of soluble trehalase by midgut columnar cells. Insect Mol. Biol. 2009, 18, 769–784. [Google Scholar] [CrossRef]
- Clegg, J.S.; Evans, D.R. Blood trehalose and flight metabolism in the blowfly. Science 1961, 134, 54–55. [Google Scholar] [CrossRef]
- Takiguchi, M.; Niimi, T.; Su, Z.H.; Yaginuma, T. Trehalase from male accessory gland of an insect, Tenebrio molitor. cDNA sequencing and developmental profile of the gene expression. Biochem. J. 1992, 288, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Mitsumasu, K.; Azuma, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. Membrane-penetrating trehalase from silkworm Bombyx mori. Molecular cloning and localization in larval midgut. Insect Mol. Biol. 2005, 14, 501–508. [Google Scholar] [CrossRef]
- Tang, B.; Chen, X.F.; Liu, Y.; Tian, H.G.; Liu, J.; Hu, J.; Xu, W.H.; Zhang, W.Q. Characterization and expression patterns of a membrane-bound trehalase from Spodoptera exigua. BMC Mol. Biol. 2008, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tang, B.; Chen, H.X.; Yao, Q.; Huang, X.F.; Chen, J.; Zhang, D.W.; Zhang, W.Q. Different functions of the insect soluble and membrane-bound trehalase genes in chitin biosynthesis revealed by RNA interference. PLoS ONE 2010, 5, e10133. [Google Scholar] [CrossRef]
- Müller, J.; Aeschbacher, R.A.; Wingler, A.; Boller, T.; Wiemken, A. Trehalose and trehalase in Arabidopsis. Plant Physiol. 2001, 125, 1086–1093. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.C.P.; Terra, W.R.; Ferreira, C. The role of carboxyl, guanidine and imidazole groups in catalysis by a midgut trehalase purified from an insect larvae. Insect Biochem. Mol. Biol. 2004, 34, 1089–1099. [Google Scholar] [CrossRef]
- De Almeida, F.M.; Bonini, B.M.; Beton, D.; Jorge, J.A.; Terenzi, H.F.; Da Silva, A.M. Heterologous expression in Escherichia coli of Neurospora crassa neutral trehalase as an active enzyme. Protein Expr. Purif. 2009, 65, 185–189. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, L.Y.; Yang, H.L.; Wang, H.J.; Zhou, M.; Wang, S.G.; Tang, B. Study on the effect of wing bud chitin metabolism and its developmental network genes in the brown planthopper, Nilaparvata lugens, by knockdown of TRE gene. Front. Physiol. 2017, 8, 750. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Li, Y.; Ren, B.; Natarajan, R. PU.1 and C/EBPα synergistically program distinct response to NF-κB activation through establishing monocyte specific enhancers. Proc. Natl. Acad. Sci. USA 2011, 108, 5290–5295. [Google Scholar] [CrossRef] [Green Version]
- Nerlov, C. The C/EBP family of transcription factors: A paradigm for interaction between gene expression and proliferation control. Trends Cell Biol. 2007, 17, 318–324. [Google Scholar] [CrossRef]
- Yang, J.Q.; Croniger, C.M.; Lekstrom-Himes, J.; Zhang, P.; Fenyus, M.; Tenen, D.G.; Darlington, G.J.; Hanson, R.W. Metabolic response of mice to a postnatal ablation of CCAAT/enhancer-binding protein α. J. Biol. Chem. 2005, 280, 38689–38699. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Qiao, L.P.; Lu, M.; Yoo, H.S.; Cheung, W.; Mak, R.; Schaack, J.; Feng, G.S.; Chi, N.-W.; Olefsky, J.M.; et al. C/EBPα regulates macrophage activation and systemic metabolism. Am. J. Physiol.-Endocrinol. Metab. 2014, 306, E1144–E1154. [Google Scholar] [CrossRef] [Green Version]
- Kagita, S.; Uppalapati, S.; Gundeti, S.; Digumarti, R. Correlation of C/EBPα expression with response and resistance to imatinib in chronic myeloid leukaemia. Jpn. J. Clin. Oncol. 2015, 45, 749–754. [Google Scholar] [CrossRef]
- Zhang, H.H.; Chen, A.Q.; Shan, T.S.; Dong, W.Y.; Shi, X.Y.; Gao, X. Cross-resistance and fitness cost analysis of resistance to thiamethoxam in melon and cotton aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2020, 113, 1946–1954. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Ma, K.S.; Tang, Q.L.; Xia, J.; Lv, N.N.; Gao, X.W. Fitness costs of sulfoxaflor resistance in the cotton aphid, Aphis gossypii Glover. Pestic. Biochem. Physiol. 2019, 158, 40–46. [Google Scholar] [CrossRef]
- Shen, G.M.; Dou, W.; Huang, Y.; Jiang, X.Z.; Smagghe, G.; Wang, J.J. In silico cloning and annotation of genes involved in the digestion, detoxification and RNA interference mechanism in the midgut of Bactrocera dorsalis [Hendel (Diptera: Tephritidae)]. Insect Mol. Biol. 2013, 22, 354–365. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.Y.; Luo, C.; Yang, G.F. Characterization of pyridalyl resistance in a laboratory-selected strain of Frankliniella occidentalis. Pestic. Biochem. Physiol. 2020, 166, 104564. [Google Scholar] [CrossRef]
- Bird, L.J.; Drynan, L.J.; Walker, P.W. Relative fitness and stability of resistance in a near-isogenic strain of indoxacarb resistant Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Manag. Sci. 2020, 76, 4077–4085. [Google Scholar] [CrossRef]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Tótola, M.R.; Guedes, R.N.C. Phosphine resistance, respiration rate and fitness consequences in stored-product insects. Pest Manag. Sci. Former. Pestic. Sci. 2007, 63, 876–881. [Google Scholar] [CrossRef] [Green Version]
- Guedes, R.N.C.; Oliveira, E.E.; Guedes, N.M.P.; Ribeiro, B.; Serrão, J.E. Cost and mitigation of insecticide resistance in the maize weevil, Sitophilus zeamais. Physiol. Entomol. 2006, 31, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Lai, X.Y.; Xu, J.; Ma, H.H.; Liu, Z.M.; Zheng, W.; Liu, J.; Zhu, H.; Zhou, Y.; Zhou, X.M. Identification and expression of inward-rectifying potassium channel subunits in Plutella xylostella. Insects 2020, 11, 461. [Google Scholar] [CrossRef]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.L.; Wu, Q.J.; Liu, Y.; Zhou, X.M.; Zhou, X.G.; Zhang, Y.J. Exploring valid reference genes for quantitative real-time PCR analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- You, Y.C.; Xie, M.; Vasseur, L.; You, M.S. Selecting and validating reference genes for quantitative real-time PCR in Plutella xylostella (L.). Genome 2018, 61, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, J.F.; Zhang, S.; Cui, J.J.; Wang, D.J.; Wang, C.Y.; Luo, J.Y.; Lv, L.M.; Ma, Y. Functional characterizations of one odorant binding protein and three chemosensory proteins from Apolygus lucorum (Meyer-Dur)(Hemiptera: Miridae) legs. J. Insect Physiol. 2013, 59, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Horváth, A.; Batki, J.; Henn, L.; Lukacsovich, T.; Róna, G.; Erdélyi, M.; Vértessy, B.G. dUTP ase expression correlates with cell division potential in Drosophila melanogaster. FEBS J. 2015, 282, 1998–2013. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Meng, X.K.; Zhang, N.; Jiang, H.; Ge, H.C.; Qian, K.; Wang, J.J. The cytosolic sulfotransferase gene TcSULT1 is involved in deltamethrin tolerance and regulated by CncC in Tribolium castaneum. Pestic. Biochem. Physiol. 2021, 177, 104905. [Google Scholar] [CrossRef]
- Araújo, R.A.; Guedes, R.N.C.; Oliveira, M.G.A.; Ferreira, G.H. Enhanced activity of carbohydrate-and lipid-metabolizing enzymes in insecticide-resistant populations of the maize weevil, Sitophilus zeamais. Bull. Entomol. Res. 2008, 98, 417–424. [Google Scholar] [CrossRef]
- ŽĎÁREK, J.; ČTVRTEČKA, R.; HOVORKA, O.; KOŠŤÁL, V. Activation of gonads and disruption of imaginal diapause in the apple blossom weevil, Anthonomus pomorum (Coleoptera: Curculionidae), with juvenoids in laboratory and field trials. EJE 2013, 97, 25–31. [Google Scholar]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316–320. [Google Scholar] [CrossRef]
- Majerowicz, D.; Gondim, K.C. Insect lipid metabolism: Insights into gene expression regulation. In Recent Trends in Gene Expression; Subhra Ngsu, S.M., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 147–189. [Google Scholar]
- Jiang, M.S.; Lane, M.D. Sequential repression and activation of the CCAAT enhancer-binding protein-α (C/EBPα) gene during adipogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 12519–12523. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) | Description |
|---|---|---|
| PxTreh1-F | CCGAGAAGGCATCAAGAAC | Promoter cloning |
| PxTreh1-R | GAGTCGTGGAAGATGCGC | |
| P(−1188/+402) | GGCTCGAGATCTGCGATCTAACTCGTTCGTTCGTTCGTG | Recombinant upstream |
| P(−610/+402) | GGCTCGAGATCTGCGATCTAACGTACGATACCGATGTAGG | |
| P(−545/+402) | GGCTCGAGATCTGCGATCTAACTGCTAACGCCATCAGTG | |
| P(−487/+402) | GGCTCGAGATCTGCGATCTAACTGTCGCAATTTATTTCCT | |
| P(−426/+402) | GGCTCGAGATCTGCGATCTAACCTAGATGGCGCTGCTAT | |
| Promoter-R | TTAGATCGCAGATCTCGAGCC | Recombinant downstream |
| PxTreh2-F | GTGAGTTTTCCCACCCAT | Promoter cloning |
| PxTreh2-R | CGAGACCGAAATTAGAAGGAA | |
| P(−1892/+196) | GGCTCGAGATCTGCGATCTAAGTAGTGAGTTTTCCCACCCAT | Recombinant upstream |
| P(−1452/+196) | GGCTCGAGATCTGCGATCTGGTTTATTATCGTCCTATCAATG | |
| P(−925/+196) | GGCTCGAGATCTGCGATCTGAGTGGAGCCAGTCCATG | |
| P(−315/+196) | GGCTCGAGATCTGCGATCTGTGACACTGCATTTGAGCG | |
| P(−254/+196) | GGCTCGAGATCTGCGATCTCATCTAGCCAACCTGGGC | |
| P(−203/+196) | GGCTCGAGATCTGCGATCTAACACGTAAGGGGACGC | |
| P(−141/+196) | GGCTCGAGATCTGCGATCTAACGACACAGATCTAAAC | |
| P(−90/+196) | GGCTCGAGATCTGCGATCTGGCAACACTGAAATCAAGGC | |
| Promoter-R | TTAGATCGCAGATCTCGAGCC | Recombinant downstream |
| C/EBPα-F | ATGGAGTCCCCACAGATGT | TF cloning |
| C/EBPα-R | AGCGCGTGGTCGCCGTGC | |
| C/EBPα-Y1H-F | ATGGAGTCCCCACAGATGT | pGADT7-PxC/EBPα |
| C/EBPα-Y1H-R | AGCGCGTGGTCGCCGTGC | |
| qC/EBPα-F | TCAAGAAGAATGGCGAAGAC | qPCR analysis |
| qC/EBPα-R | CAGGTCCAGGCTGATCTC | |
| qTreh1-F | TTCCACGACTCCAAGACCTT | |
| qTreh1-R | CAGCAGCACAGCGAAGTC | |
| qTreh2-F | ATGCTCTCCAACTTCCTCAAC | |
| qTreh2-R | GCGGCTGTGACCTCATAG | |
| EF1-F | GCCTCCCTACAGCGAATC | |
| EF1-R | CCTTGAACCAGGGCATCT | |
| RSP4-F | ATGGATGTTGTGTCGATTGAAAAGA | |
| RSP4-R | GAGTGATGCGGTGGATGGTGA | |
| dsC/EBPα-F | T7-TGGACGAGCTCAACGGCCAG | dsRNA synthesis |
| dsC/EBPα-R | T7-TGAGCAGAGGAGGCAGCAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, Z.; Ma, H.; Yu, Y.; Zhang, C.; Zheng, W.; Man, Y.; Zhu, H.; Zhou, Y.; Chen, X.; et al. C/EBPα Regulates PxTreh1 and PxTreh2 Trehalase-Related Bt Resistance in Plutella xylostella (L.). Insects 2022, 13, 340. https://doi.org/10.3390/insects13040340
Liu J, Liu Z, Ma H, Yu Y, Zhang C, Zheng W, Man Y, Zhu H, Zhou Y, Chen X, et al. C/EBPα Regulates PxTreh1 and PxTreh2 Trehalase-Related Bt Resistance in Plutella xylostella (L.). Insects. 2022; 13(4):340. https://doi.org/10.3390/insects13040340
Chicago/Turabian StyleLiu, Jia, Zheming Liu, Haihao Ma, Yaying Yu, Chengjia Zhang, Wei Zheng, Yilong Man, Hang Zhu, Yong Zhou, Xi Chen, and et al. 2022. "C/EBPα Regulates PxTreh1 and PxTreh2 Trehalase-Related Bt Resistance in Plutella xylostella (L.)" Insects 13, no. 4: 340. https://doi.org/10.3390/insects13040340
APA StyleLiu, J., Liu, Z., Ma, H., Yu, Y., Zhang, C., Zheng, W., Man, Y., Zhu, H., Zhou, Y., Chen, X., Zhou, X., & Zeng, A. (2022). C/EBPα Regulates PxTreh1 and PxTreh2 Trehalase-Related Bt Resistance in Plutella xylostella (L.). Insects, 13(4), 340. https://doi.org/10.3390/insects13040340