
Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
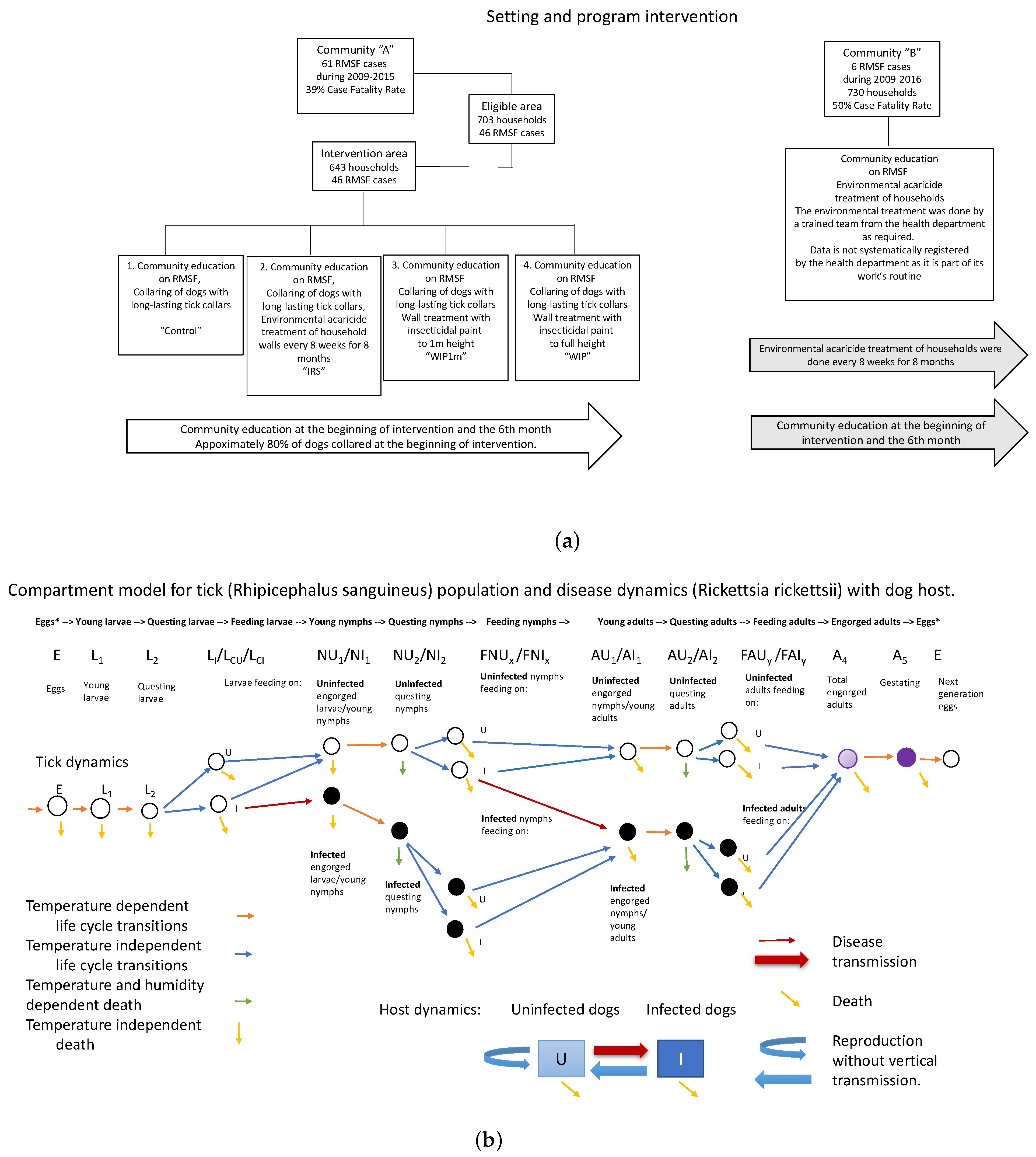
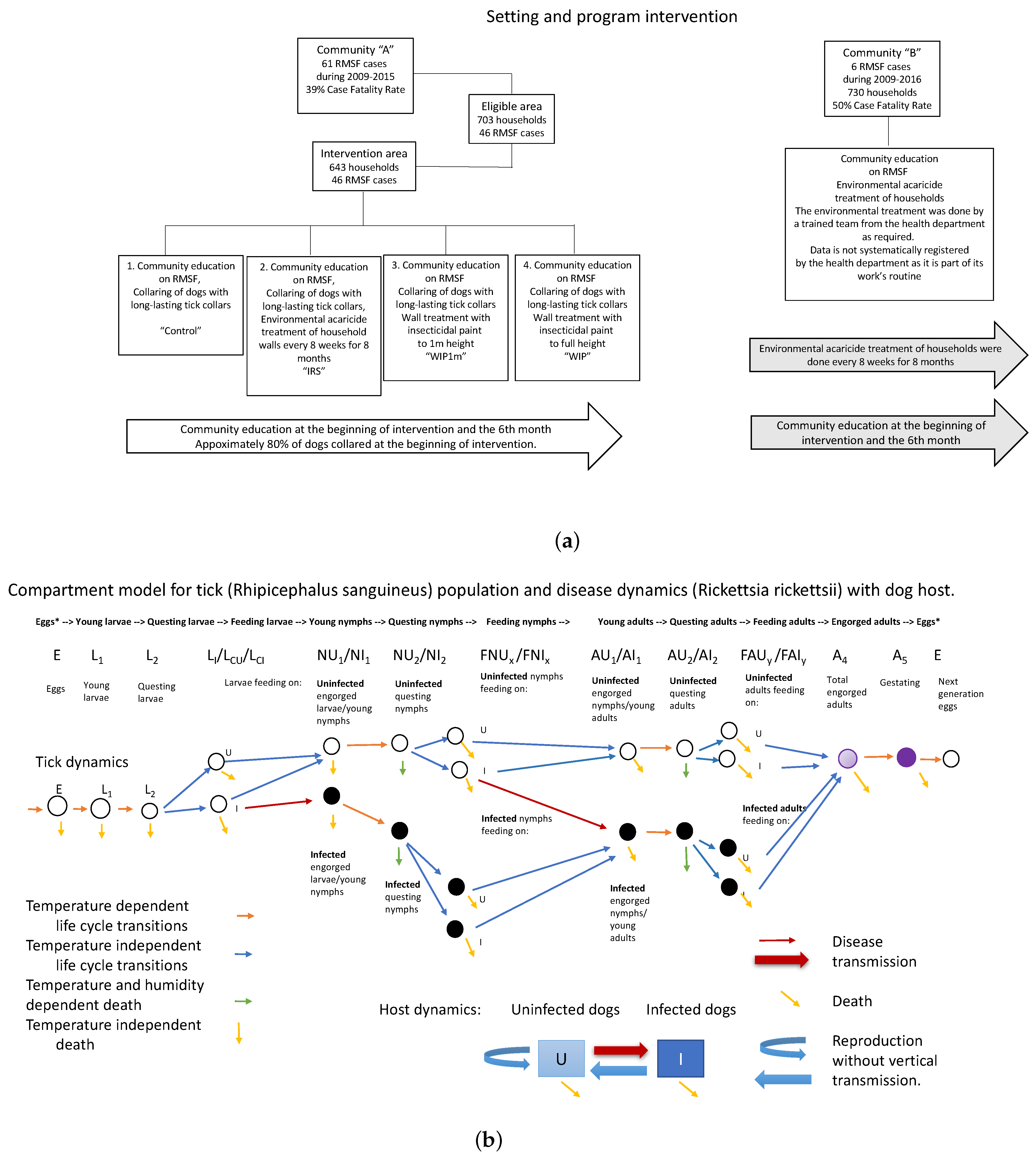
2.1. Epidemiological Intervention
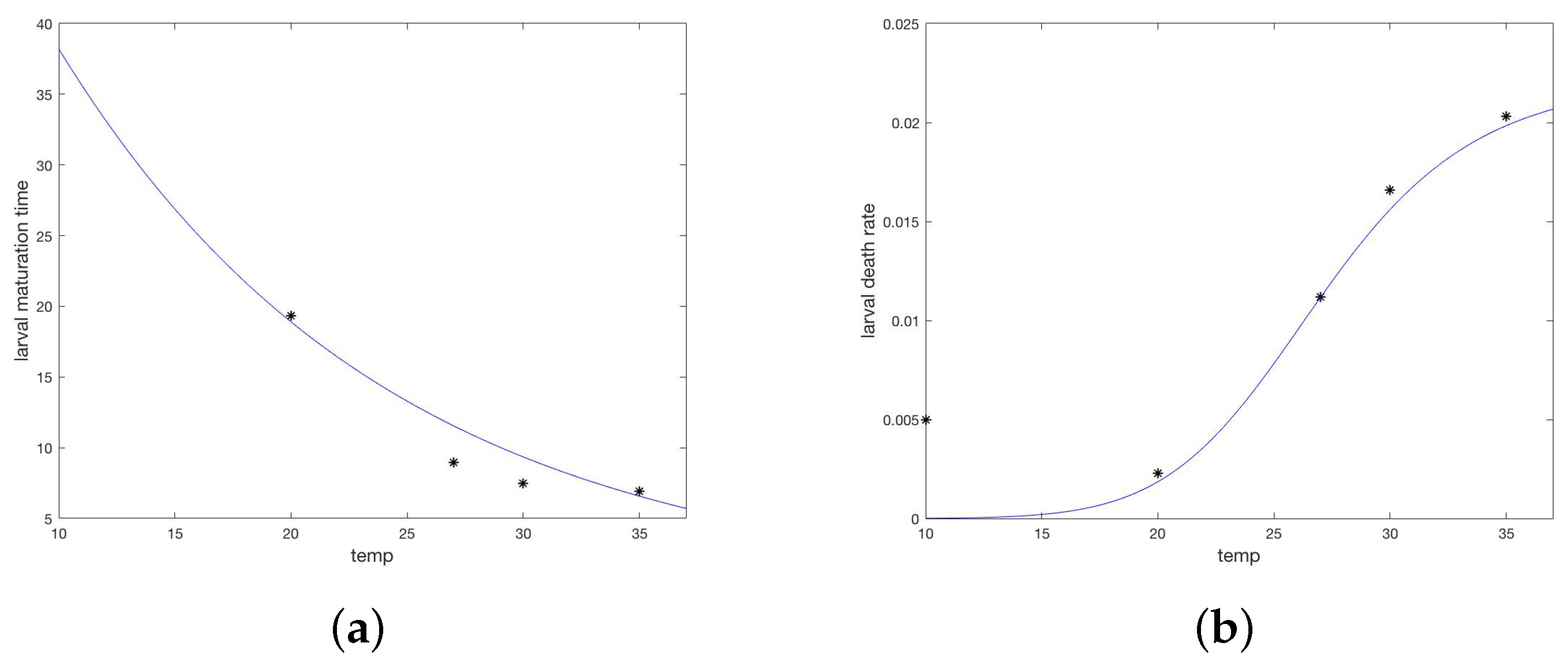
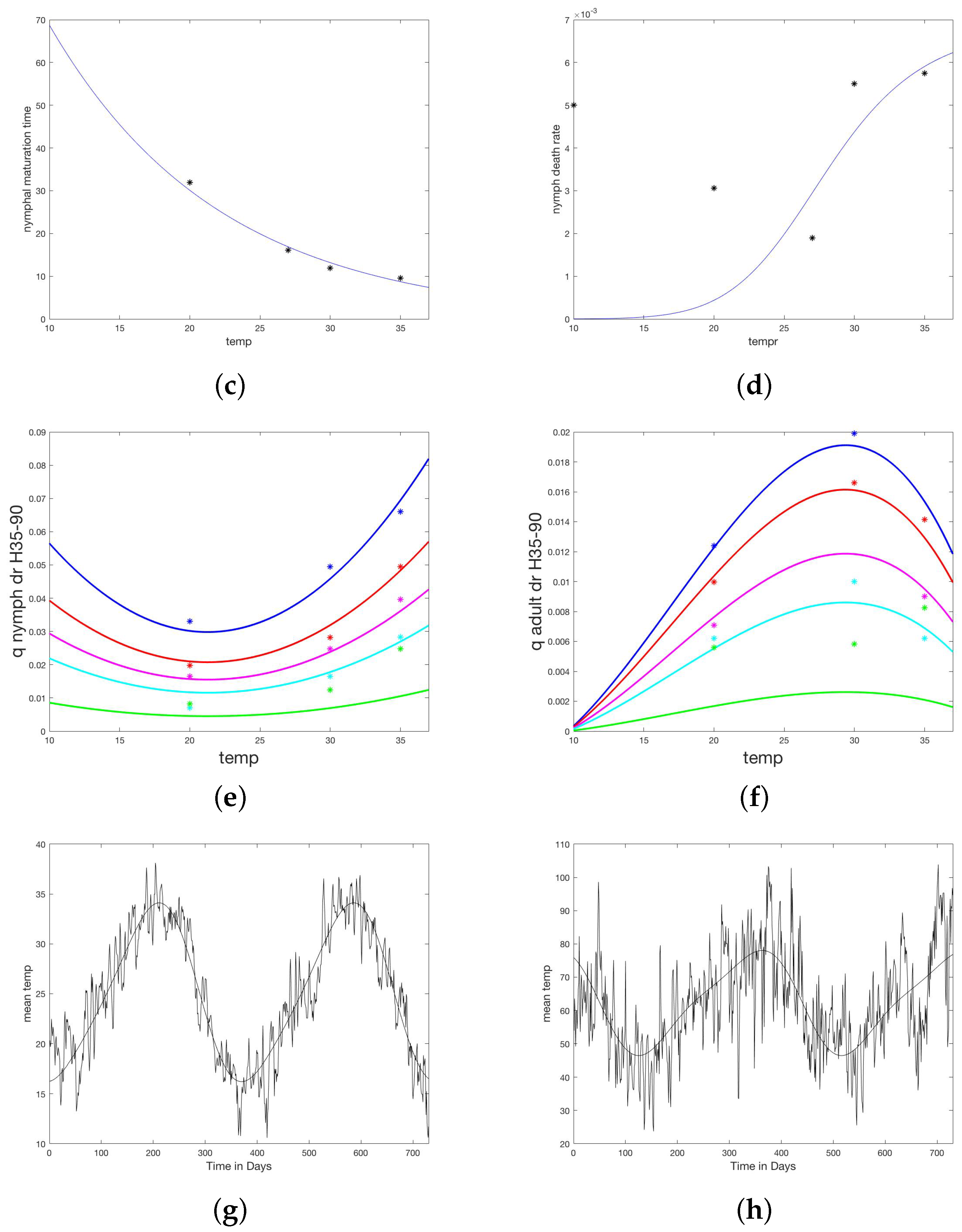
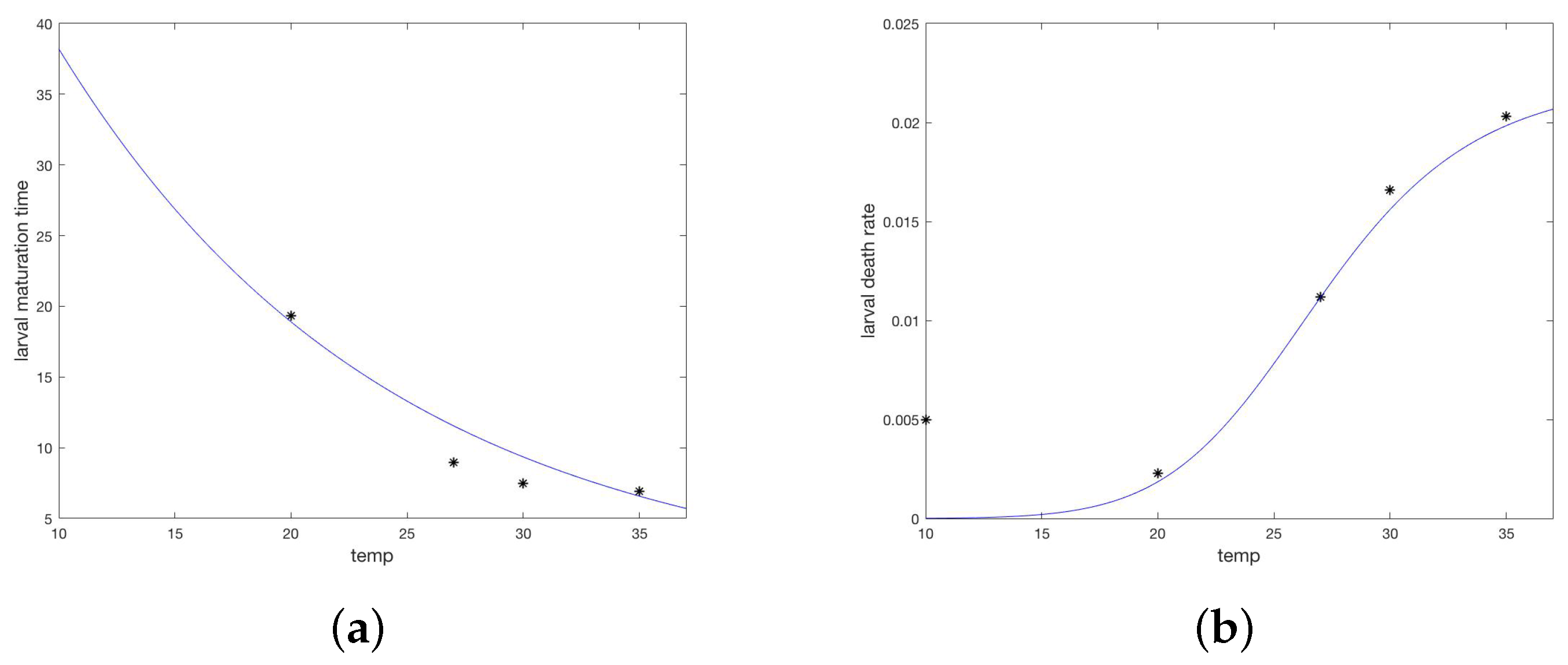
2.2. Model Development, Parameterization, and Numerical Simulations
2.3. Numerical Simulations
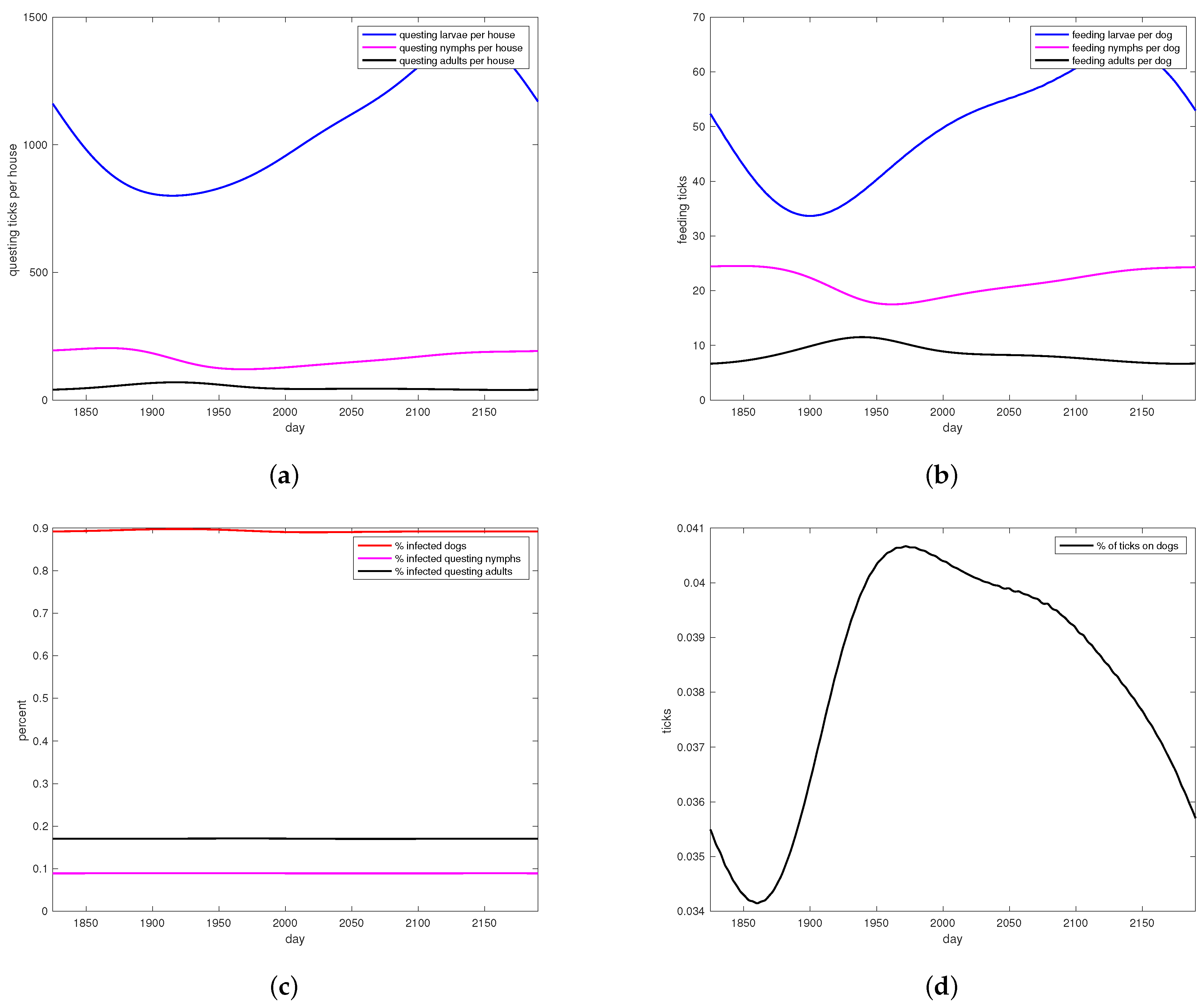
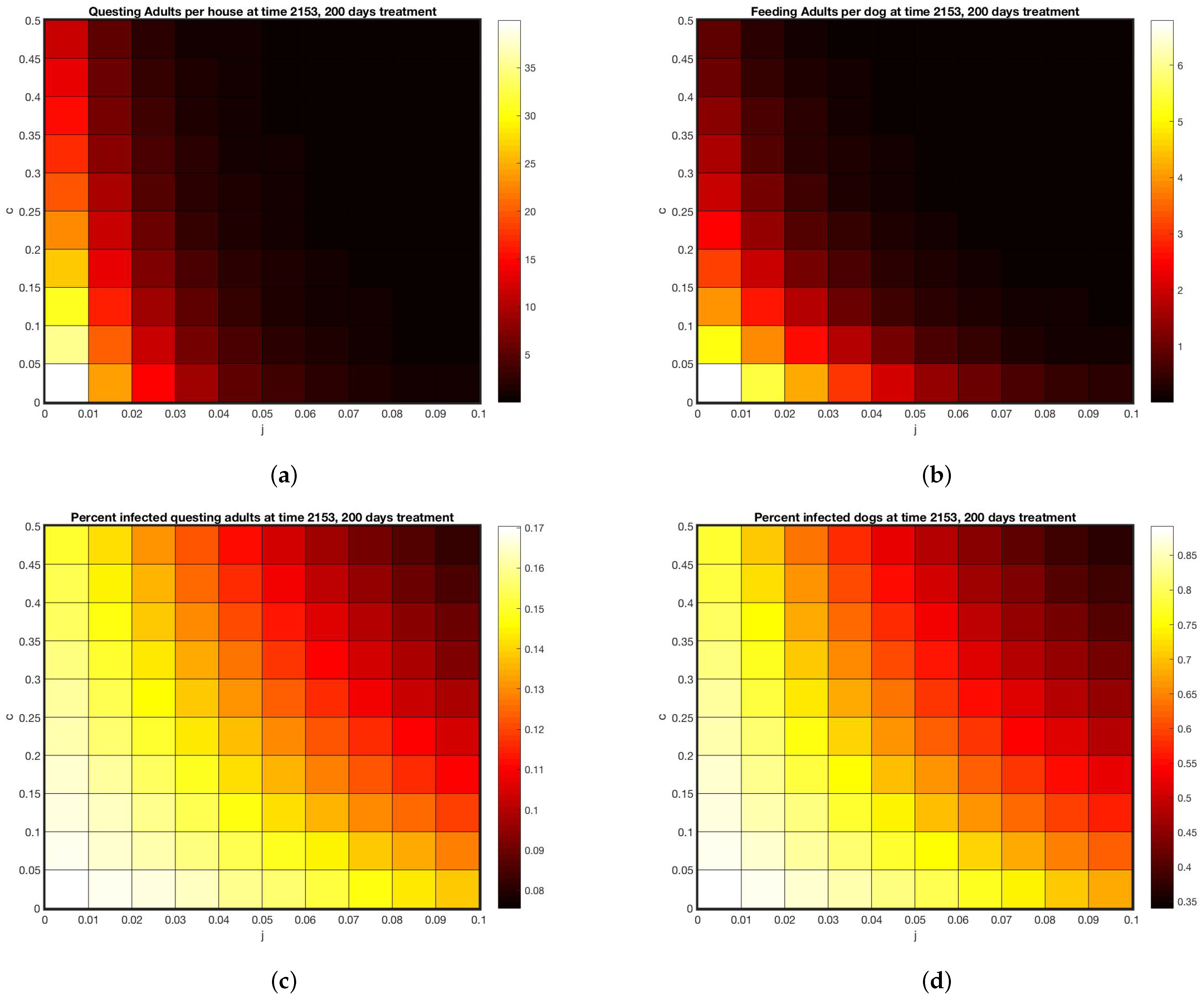
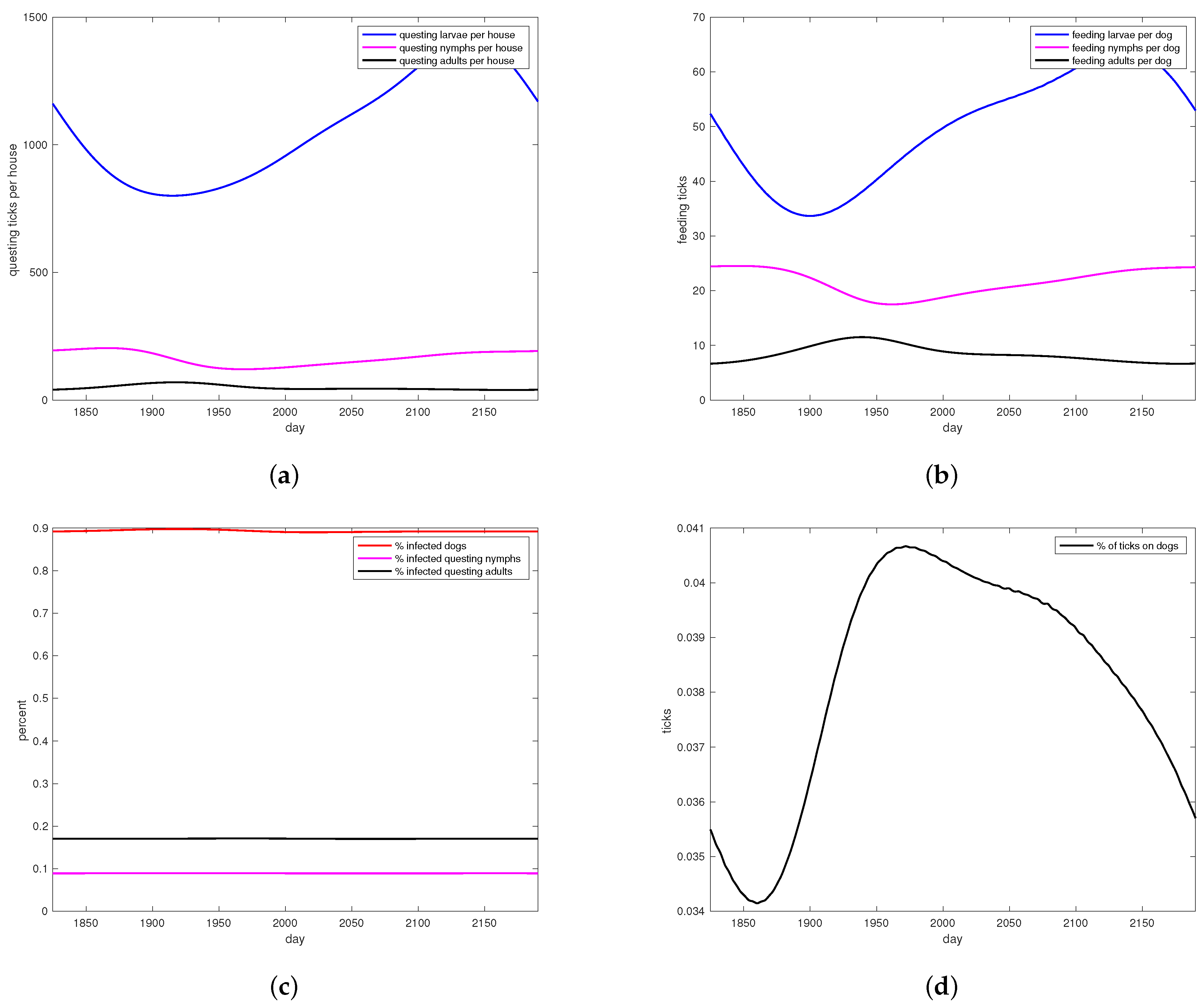
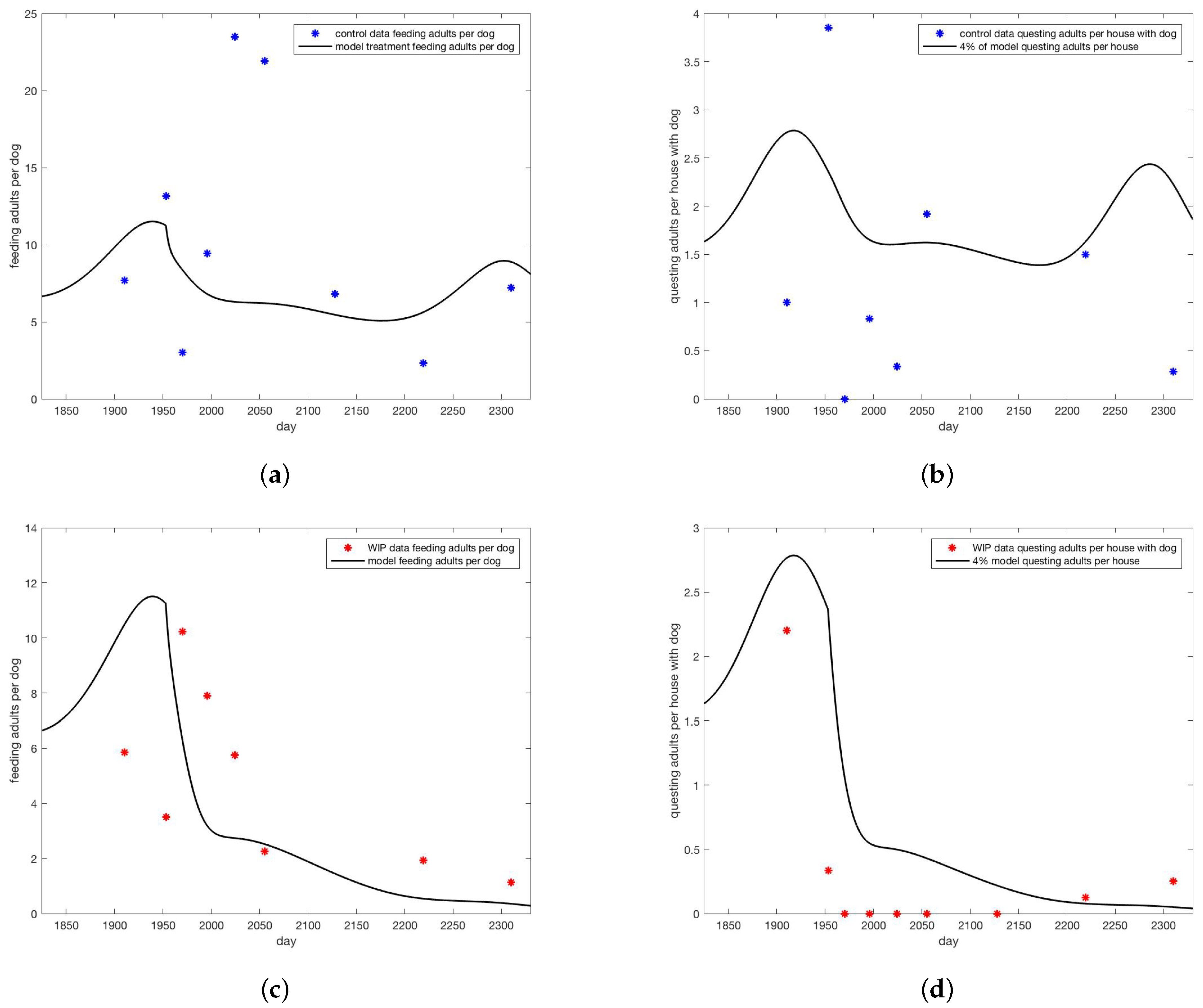
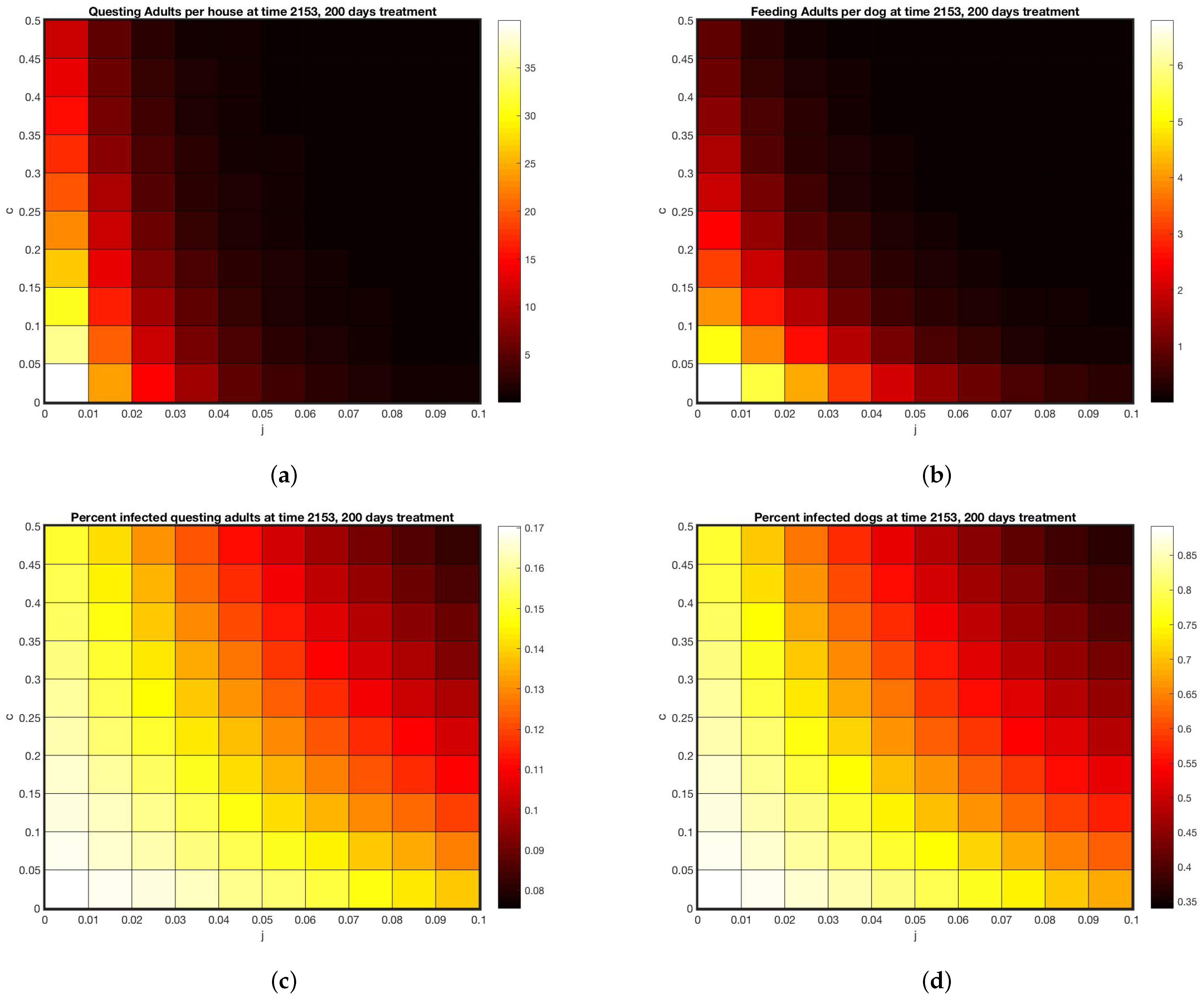
3. Results
4. Discussion
4.1. Comparison of Test Groups
4.2. Challenges in Surveying Ticks Both on and off Host
4.3. Model Performance and Results
4.4. Factors Contributing to Discrepancies between Model and Data
4.5. Future Work
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Model Details
Appendix A.1. Differential Equations
Appendix A.2. Auxiliary Equations
Appendix A.3. Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Units |
|---|---|---|---|
| b | oviposition rate | 62.56 | eggs per tick per day |
| daily death rate of eggs | 0.015 | percent per day | |
| daily death rate of feeding larvae | =0.2 | percent per day | |
| daily death rate of feeding nymphs | =0.01 | percent per day | |
| daily death rate of feeding adults | 0.01 | percent per day | |
| daily death rate of gestating adults | 0.0351 | percent per day | |
| young larvae to questing larvae maturation rate | 0.069 | percent per day | |
| questing larvae to feeding larvae maturation | 0.1 | percent per day | |
| larvae feeding on host maturation rate | 0.232 | percent per day | |
| uesting nymph maturation rate | 0.1 | percent per day | |
| feeding nymph maturation rate | 0.142 | percent per day | |
| questing adult maturation rate | 0.1 | percent per day | |
| feeding adult maturation rate | 0.1058 | percent per day | |
| C | per host carrying capacity | 50 | maximum feeding nymphs and adults per host |
| birth rate of host | 0.0135 | per dog per day | |
| carrying capacity of hosts | 2000 | number of dogs | |
| death rate of uninfected host | 0.0002739726 | percent per day | |
| death rate of infected hosts | 0.005479 | percent per day | |
| probability of host infection by one feeding tick | 0.0001 | per infective tick per day | |
| percent of feeding larvae infected | 0.1 | percent per day | |
| percent of feeding nymphs infected | 0.1 | percent per day | |
| numerical feature | 0.01 | no units |
Appendix B. Rhipicephalus sanguineus Ticks Collected from Dogs during Home Inspections by Treatment (Pre and Post Intervention) for the Study Period, 2018–2019
| Treatment | # Time at Treatment | # Dogs Inspected | # Inspected Infested with Ticks | R. sanguineus Larvae | R. sanguineus Nymphs | R. sanguineus Adults | Total Ticks Found per Inspection Date |
| Pre Intervention | |||||||
| Control | Week 0 | 35 | 15 | 55 | 76 | 269 | 400 |
| WIP1m | Week 0 | 20 | 14 | 22 | 17 | 885 | 924 |
| WIP | Week 0 | 34 | 15 | 5 | 9 | 199 | 213 |
| IRS-PPX | Week 0 | 28 | 4 | 0 | 0 | 17 | 17 |
| Post-Intervention | |||||||
| Control | Week 4 | 20 | 18 | 19 | 18 | 263 | 300 |
| Control | Month 1 | 9 | 8 | 4 | 2 | 27 | 33 |
| Control | Month 2 | 9 | 8 | 0 | 3 | 85 | 88 |
| Control | Month 3 | 2 | 2 | 1 | 1 | 47 | 49 |
| Control | Month 4 | 21 | 19 | 2 | 115 | 460 | 577 |
| Control | Month 6 | 5 | 4 | 0 | 0 | 34 | 34 |
| Control | Month 9 | 6 | 4 | 0 | 12 | 14 | 26 |
| Control | Month 12 | 5 | 3 | 0 | 3 | 36 | 39 |
| Total | 77 | 66 | 26 | 154 | 966 | ||
| WIP1m | Week 4 | 16 | 6 | 0 | 0 | 11 | 11 |
| WIP1m | Month 1 | 19 | 17 | 226 | 200 | 685 | 1111 |
| WIP1m | Month 2 | 7 | 7 | 35 | 42 | 103 | 180 |
| WIP1m | Month 3 | 10 | 10 | 181 | 498 | 150 | 829 |
| WIP1m | Month 4 | 6 | 6 | 16 | 77 | 237 | 330 |
| WIP1m | Month 6 | 9 | 6 | 9 | 33 | 208 | 250 |
| WIP1m | Month 9 | 10 | 4 | 4 | 0 | 14 | 18 |
| WIP1m | Month 12 | 9 | 7 | 24 | 107 | 154 | 285 |
| Total | 86 | 63 | 495 | 957 | 1562 | ||
| WIP1m | Week 4 | 12 | 6 1 | 1 | 42 | 44 | |
| WIP | Month 1 | 9 | 8 | 15 | 4 | 92 | 111 |
| WIP | Month 2 | 10 | 7 | 0 | 11 | 79 | 90 |
| WIP | Month 3 | 12 | 9 | 20 | 20 | 69 | 109 |
| WIP | Month 4 | 12 | 7 | 0 | 4 | 27 | 31 |
| WIP | Month 6 | 11 | 7 | 0 | 39 | 140 | 179 |
| WIP | Month 9 | 16 | 13 | 3 | 40 | 31 | 74 |
| WIP | Month 12 | 23 | 7 | 0 | 65 | 28 | 93 |
| Total | 105 | 64 | 39 | 184 | 508 | ||
| IRS-PPX | Week 4 | 13 | 1 | 0 | 0 | 2 | 2 |
| IRS-PPX | Month 1 | 14 | 2 | 0 | 0 | 2 | 2 |
| IRS-PPX | Month 2 | 13 | 10 | 0 | 1 | 67 | 68 |
| IRS-PPX | Month 3 | 14 | 8 | 8 | 16 | 78 | 102 |
| IRS-PPX | Month 4 | 7 | 7 | 8 | 1 | 28 | 37 |
| IRS-PPX | Month 6 | 15 | 10 | 39 | 38 | 168 | 245 |
| IRS-PPX | Month 9 | 21 | 8 | 0 | 1 | 25 | 26 |
| IRS-PPX | Month 12 | 13 | 4 | 1 | 11 | 5 | 17 |
| Total | 110 | 50 | 56 | 68 | 375 |
References
- Álvarez-Hernández, G.; Roldán, J.F.G.; Milan, N.S.H.; Lash, R.R.; Behravesh, C.B.; Paddock, C.D. Rocky Mountain spotted fever in Mexico: Past, present, and future. Lancet Infect. Dis. 2017, 17, e189–e196. [Google Scholar] [CrossRef]
- Kirkland, K.B.; Wilkinson, W.E.; Sexton, D.J. Therapeutic delay and mortality in cases of Rocky Mountain spotted fever. Clin. Infect. Dis. 1995, 20, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Biggs, H.M.; Behravesh, C.B.; Bradley, K.K.; Dahlgren, F.S.; Drexler, N.A.; Dumler, J.S.; Folk, S.M.; Kato, C.Y.; Lash, R.R.; Levin, M.L.; et al. Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever and other spotted fever group rickettsioses, ehrlichioses, and anaplasmosisnited States: A practical guide for health care and public health professionals. Morb. Mortal. Wkly. Rep. Recomm. Rep. 2016, 65, 1–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paddock, C.D.; Alvarez-Hernández, G. Rickettsia rickettsii (Rocky Mountain spotted fever). In Principles and Practice of Pediatric Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2018; pp. 952–957. [Google Scholar]
- Alvarez, D.; Ochoa, E.; Nichols Heitman, K.; Binder, A.M.; Alvarez, G.; Armstrong, P.A. Epidemiology and clinical features of Rocky Mountain spotted fever from enhanced surveillance, Sonora, Mexico: 2015–2018. Am. J. Trop. Med. Hyg. 2020, 104, 190–197. [Google Scholar] [CrossRef]
- Helmick, C.G.; Bernard, K.W.; D’Angelo, L.J. Rocky Mountain spotted fever: Clinical, laboratory, and epidemiological features of 262 cases. J. Infect. Dis. 1984, 150, 480–488. [Google Scholar] [CrossRef]
- Chen, L.F.; Sexton, D.J. What’s new in Rocky Mountain spotted fever? Infect. Dis. Clin. N. Am. 2008, 22, 415–432. [Google Scholar] [CrossRef]
- Alvarez-Hernandez, G.; Murillo-Benitez, C.; del Carmen Candia-Plata, M.; Moro, M. Clinical profile and predictors of fatal Rocky Mountain spotted fever in children from Sonora, Mexico. Pediatr. Infect. Dis. J. 2015, 34, 125–130. [Google Scholar] [CrossRef]
- Blanton, L.S. The rickettsioses: A practical update. Infect. Dis. Clin. 2019, 33, 213–229. [Google Scholar] [CrossRef]
- Walker, D.H.; Myers, C.T.E.; Blanton, L.S.; Bloch, K.C.; Fowler, V.G., Jr.; Gaines, D.N.; Paddock, C.D.; Yaglom, H.D. Rickettsiosis subcommittee report to the tick-borne disease working group. Ticks Tick-Borne Dis. 2021, 13, 101855. [Google Scholar] [CrossRef]
- Archibald, L.K.; Sexton, D.J. Long-term sequelae of Rocky Mountain spotted fever. Clin. Infect. Dis. 1995, 20, 1122–1125. [Google Scholar] [CrossRef]
- Sekeyová, Z.; Danchenko, M.; Filipčík, P.; Fournier, P.E. Rickettsial infections of the central nervous system. PLoS Negl. Trop. Dis. 2019, 13, e0007469. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.D.D.L.; Licona-Enríquez, J.D.; Leyva-Gastélum, M.; Mora, D.D.D.L.; Rascón-Alcantar, A.; Álvarez-Hernández, G. Una serie de casos fatales de fiebre manchada de las Montañas Rocosas en Sonora, México. Biomédica 2018, 38, 69–76. [Google Scholar]
- Licona-Enriquez, J.D.; Delgado-De La Mora, J.; Paddock, C.D.; Ramirez-Rodriguez, C.A.; del Carmen Candia-Plata, M.; Hernández, G.Á. Rocky Mountain spotted fever and pregnancy: Four cases from Sonora, Mexico. Am. J. Trop. Med. Hyg. 2017, 97, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zazueta, O.E.; Armstrong, P.A.; Márquez-Elguea, A.; Milán, N.S.H.; Peterson, A.E.; Ovalle-Marroquín, D.F.; Fierro, M.; Arroyo-Machado, R.; Rodriguez-Lomeli, M.; Trejo-Dozal, G.; et al. Rocky Mountain Spotted Fever in a Large Metropolitan Center, Mexico–United States Border, 2009–2019. Emerg. Infect. Dis. 2021, 27, 1567. [Google Scholar] [CrossRef]
- Alvarez, G.; Rosales, C.; Sepulveda, R. Rocky mountain spotted fever, a reemerging disease in Arizona and Sonora–Case study. J. Case Rep. Stud. 2014, 1, 601. [Google Scholar]
- Eremeeva, M.E.; Zambrano, M.L.; Anaya, L.; Beati, L.; Karpathy, S.E.; Santos-Silva, M.M.; Salceda, B.; Macbeth, D.; Olguin, H.; Dasch, G.A.; et al. Rickettsia rickettsii in Rhipicephalus ticks, mexicali, Mexico. J. Med. Entomol. 2011, 48, 418–421. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.; Dantas-Torres, F.; Estrada-Peña, A.; Levin, M. Systematics and ecology of the brown dog tick, Rhipicephalus sanguineus. Ticks -Tick-Borne Dis. 2013, 4, 171–180. [Google Scholar] [CrossRef]
- Koch, H.G.; Tuck, M.D. Molting and survival of the brown dog tick (Acari: Ixodidae) under different temperatures and humidities. Ann. Entomol. Soc. Am. 1986, 79, 11–14. [Google Scholar] [CrossRef]
- López-Pérez, A.M.; Orozco, L.; Zazueta, O.E.; Fierro, M.; Gomez, P.; Foley, J. An exploratory analysis of demography and movement patterns of dogs: New insights in the ecology of endemic Rocky Mountain-Spotted Fever in Mexicali, Mexico. PLoS ONE 2020, 15, e0233567. [Google Scholar] [CrossRef]
- Alvarez-Hernandez, G.; Drexler, N.; Paddock, C.D.; Licona-Enriquez, J.D.; la Mora, J.D.D.; Straily, A.; del Carmen Candia-Plata, M.; Cruz-Loustaunau, D.I.; Arteaga-Cardenas, V.A. Community-based prevention of epidemic Rocky Mountain spotted fever among minority populations in Sonora, Mexico, using a One Health approach. Trans. R. Soc. Trop. Med. Hyg. 2020, 114, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Straily, A.; Drexler, N.; Cruz-Loustaunau, D.; Paddock, C.; Alvarez-Hernandez, G. Notes from the field: Community-based prevention of Rocky Mountain spotted fever–Sonora, Mexico, 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1302–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, D.; Ratti, V.; Kodali, A.; Winter, J.M.; Ayres, M.P.; Chipman, J.W.; Aoki, C.F.; Osterberg, E.C.; Silvanic, C.; Partridge, T.F.; et al. Effect of rising temperature on Lyme disease: Ixodes scapularis population dynamics and Borrelia burgdorferi transmission and prevalence. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 9817930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratti, V.; Winter, J.M.; Wallace, D.I. Dilution and amplification effects in Lyme disease: Modeling the effects of reservoir-incompetent hosts on Borrelia burgdorferi sensu stricto transmission. Ticks Tick-Borne Dis. 2021, 12, 101724. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.M.; Partridge, T.F.; Wallace, D.; Chipman, J.W.; Ayres, M.P.; Osterberg, E.C.; Dekker, E.R. Modeling the sensitivity of blacklegged ticks (Ixodes scapularis) to temperature and land cover in the northeastern United States. J. Med. Entomol. 2021, 58, 416–427. [Google Scholar] [CrossRef]
- Feldman-Muhsam, B. Some observations on the hibernation of Rhipicephalus sanguineus in Jerusalem. In Tick Biology and Control: Proceedings of an International Conference, 27–29 January 1981; Whitehead, G.B., Gibson, J.D., Eds.; Tick Research Unit, Rhodes University: Grahamstown, South Africa, 1981. [Google Scholar]
- Dantas-Torres, F.; Giannelli, A.; Otranto, D. Starvation and overwinter do not affect the reproductive fitness of Rhipicephalus sanguineus. Vet. Parasitol. 2012, 185, 260–264. [Google Scholar] [CrossRef]
- Oyarzún-Ruiz, P.; Espinoza-Carniglia, M.; Reidembach, S.; Muñoz, P.; Moreno, L. Expansion in the latitudinal distribution of Rhipicephalus sanguineus sensu stricto (Acari: Ixodidae) to southern Chile. Exp. Appl. Acarol. 2021, 83, 107–114. [Google Scholar] [CrossRef]
- Sannasi, A.; Subramoniam, T. Hormonal rupture of larval diapause in the tick Rhipicephalus sanguineus (Lat.). Experientia 1972, 28, 666–667. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Capelli, G.; Giannelli, A.; Ramos, R.A.N.; Lia, R.P.; Cantacessi, C.; De Caprariis, D.; De Tommasi, A.S.; Latrofa, M.S.; Lacasella, V.; et al. Efficacy of an imidacloprid/flumethrin collar against fleas, ticks and tick-borne pathogens in dogs. Parasites Vectors 2013, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stanneck, D.; Kruedewagen, E.M.; Fourie, J.J.; Horak, I.G.; Davis, W.; Krieger, K.J. Efficacy of an imidacloprid/flumethrin collar against fleas, ticks, mites and lice on dogs. Parasites Vectors 2012, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- MATLAB R2016a; TheMathWorks, Inc.: Natick, MA, USA, 2016.
- Explorer, W.C. ERA5 Reanalysis Data. Available online: http://https://climexp.knmi.nl/selectdailyfield2.cgi (accessed on 31 March 2021).
- National Weather Service, U. Relative Humidity and Dewpoint Temperature from Temperature and Wet Bulb Temperature. Available online: http://https://www.weather.gov/epz/wxcalc (accessed on 31 March 2021).
- Ioffe-Uspensky, I.; Mumcuoglu, K.Y.; Uspensky, I.; Galun, R. Rhipicephalus sanguineus and R. turanicus (Acari: Ixodidae): Closely related species with different biological characteristics. J. Med. Entomol. 1997, 34, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Varma, M. The culture of the tick Rhipicephalus sanguineus (Latreille)(Ixodidae) in the laboratory. J. Med. Entomol. 1964, 1, 154–157. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Lotufo Brant de Carvalho, J.; Andrea de Santis Bastos, P.; Katagiri, S.; Yamada Batalha, E.; Okano, W.; Maia da Costa, V. Prevalence of Rickettsia rickettsii in Ticks: Systematic Review and Meta-analysis. Vector-Borne Zoonotic Dis. 2021, 21, 557–565. [Google Scholar] [PubMed]
- Foley, J.; Tinoco-Gracia, L.; Rodriguez-Lomelí, M.; Estrada-Guzmán, J.; Fierro, M.; Mattar-Lopez, E.; Peterson, A.; Pascoe, E.; Gonzalez, Y.; Hori-Oshima, S.; et al. Unbiased assessment of abundance of Rhipicephalus sanguineus sensu lato ticks, canine exposure to spotted fever group Rickettsia, and risk factors in Mexicali, México. Am. J. Trop. Med. Hyg. 2019, 101, 22. [Google Scholar] [CrossRef]
- Diniz, P.P.V.; Beall, M.J.; Omark, K.; Chandrashekar, R.; Daniluk, D.A.; Cyr, K.E.; Koterski, J.F.; Robbins, R.G.; Lalo, P.G.; Hegarty, B.C.; et al. High prevalence of tick-borne pathogens in dogs from an Indian reservation in northeastern Arizona. Vector-Borne Zoonotic Dis. 2010, 10, 117–123. [Google Scholar] [CrossRef]
- Piranda, E.M.; Faccini, J.L.H.; Pinter, A.; Pacheco, R.C.; Cançado, P.H.; Labruna, M.B. Experimental infection of Rhipicephalus sanguineus ticks with the bacterium Rickettsia rickettsii, using experimentally infected dogs. Vector-Borne Zoonotic Dis. 2011, 11, 29–36. [Google Scholar] [CrossRef]
- Dantas-Torres, F. The brown dog tick, Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae): From taxonomy to control. Vet. Parasitol. 2008, 152, 173–185. [Google Scholar] [CrossRef]
- Silveira, J.A.; Passos, L.M.; Ribeiro, M.F. Population dynamics of Rhipicephalus sanguineus (Latrielle, 1806) in Belo Horizonte, Minas Gerais state, Brazil. Vet. Parasitol. 2009, 161, 270–275. [Google Scholar] [CrossRef]
- Needham, G.R.; Teel, P.D. Off-host physiological ecology of ixodid ticks. Annu. Rev. Entomol. 1991, 36, 659–681. [Google Scholar] [CrossRef]
- Hadani, A.; Rechav, Y. Tick-host relationships 1. The existence of a circadian rhythm of “drop-off” of engorged ticks from their hosts. Acta Trop. 1969, 26, 173–179. [Google Scholar]
- Jacobs, P.; Fourie, L.; Horak, I. A laboratory comparison of the life cycles of the dog ticks Haemaphysalis leachi and Rhipicephalus sanguineus. Onderstepoort J. Vet. Res. 2004, 71, 15–28. [Google Scholar] [PubMed]
- Ortega-Morales, A.I.; Nava-Reyna, E.; Ávila-Rodríguez, V.; González-Álvarez, V.H.; Castillo-Martínez, A.; Siller-Rodríguez, Q.K.; Cabezas-Cruz, A.; Dantas-Torres, F.; Almazán, C. Detection of Rickettsia spp. in Rhipicephalus sanguineus (sensu lato) collected from free-roaming dogs in Coahuila state, northern Mexico. Parasites Vectors 2019, 12, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Treatment | # Houses Inspected | # Houses with Tick Infestations | % Houses Inspected with Dogs Present (n) | R. sanguineus Larvae | R. sanguineus Nymphs | R. sanguineus Adults | R. sanguineus Total |
|---|---|---|---|---|---|---|---|
| Pre Intervention | |||||||
| Day 1 | |||||||
| Control | 19 | 2 | 80 (n = 12) | 0 | 1 | 19 | 20 |
| WIP1m | 15 | 0 | 47 (n = 7) | 0 | 0 | 0 | 0 |
| WIP | 15 | 4 | 80 (n = 12) | 0 | 2 | 33 | 35 |
| IRS-PPX | 12 | 0 | 100 (n = 12) | 0 | 0 | 0 | 0 |
| Total | 61 | 6 | 0 | 3 | 52 | 55 | |
| Post-Intervention | |||||||
| Days 2–9 | |||||||
| Control | 60 | 15 | 75 (n = 45) | 45 | 28 | 119 | 192 |
| WIP1m | 53 | 9 | 58 (n = 31) | 9 | 21 | 12 | 42 |
| WIP | 53 | 6 | 92 (n = 49) | 0 | 6 | 5 | 11 |
| IRS-PPX | 55 | 11 | 75 (n = 41) | 45 | 17 | 42 | 104 |
| Total | 221 | 41 | 99 | 72 | 178 | 349 |
| Treatment Group | Mean Adults Per Dog (SDpop) | Mean Adults Per House (SDpop) | Mean Adults Per House With Dog (SDpop) |
|---|---|---|---|
| Control | 10.56 (7.17) | 1.50 (1.36) | 1.89 (1.54) |
| WIP1m | 21.31 (14.91) | 0.18 (0.29) | 0.278 (0.416) |
| WIP | 5.71 (3.74) | 0.32 (0.67) | 0.384 (0.806) |
| IRS-PPX | 3.16 (3.52) | 0.65 (1.20) | 0.708 (1.195) |
| Comparison | WIP1m | WIP | IRS-PPX |
|---|---|---|---|
| Control | Adults Per House With Dog: Control > WIP1m () | Adults Per House: Control > WIP () Adults Per House With Dog: Control > WIP () | |
| WIP1m | Adults Per dog: WIP1m > WIP () | Adults Per dog: WIP1m > IRS-PPX () |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Hernandez, G.; Trejo, A.V.; Ratti, V.; Teglas, M.; Wallace, D.I. Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico. Insects 2022, 13, 263. https://doi.org/10.3390/insects13030263
Alvarez-Hernandez G, Trejo AV, Ratti V, Teglas M, Wallace DI. Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico. Insects. 2022; 13(3):263. https://doi.org/10.3390/insects13030263
Chicago/Turabian StyleAlvarez-Hernandez, Gerardo, Alejandro Villegas Trejo, Vardayani Ratti, Michael Teglas, and Dorothy I. Wallace. 2022. "Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico" Insects 13, no. 3: 263. https://doi.org/10.3390/insects13030263
APA StyleAlvarez-Hernandez, G., Trejo, A. V., Ratti, V., Teglas, M., & Wallace, D. I. (2022). Modeling of Control Efforts against Rhipicephalus sanguineus, the Vector of Rocky Mountain Spotted Fever in Sonora Mexico. Insects, 13(3), 263. https://doi.org/10.3390/insects13030263