
Comparative Analysis of Mitochondrial Genomes among Twelve Sibling Species of the Genus Atkinsoniella Distant, 1908 (Hemiptera: Cicadellidae: Cicadellinae) and Phylogenetic Analysis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Genome Organization
2.2. PCR Amplification, Sequencing, and Assembly
2.3. Mitogenome Annotation and Analysis
2.4. Phylogenetic Analysis
3. Results
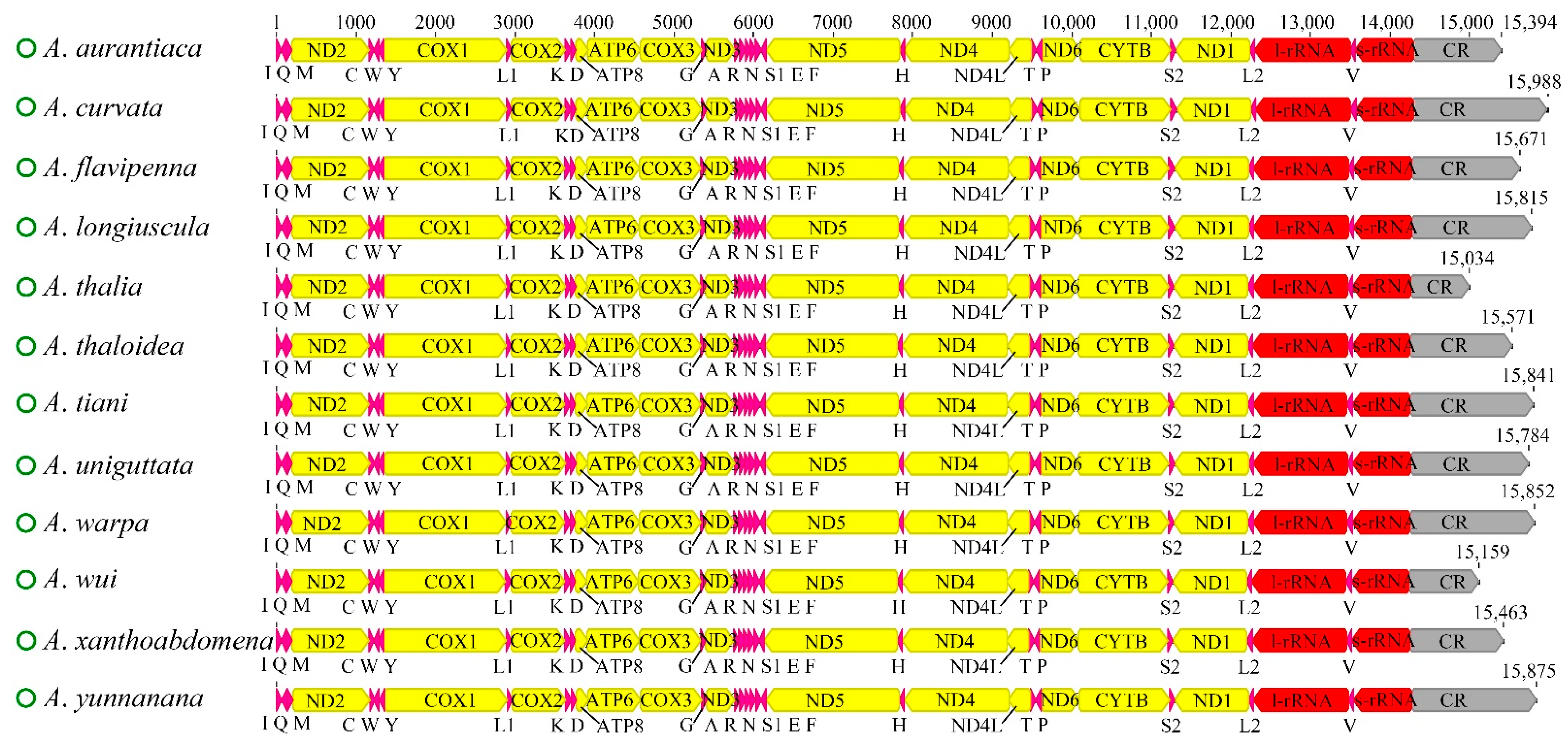
3.1. Genome Organization
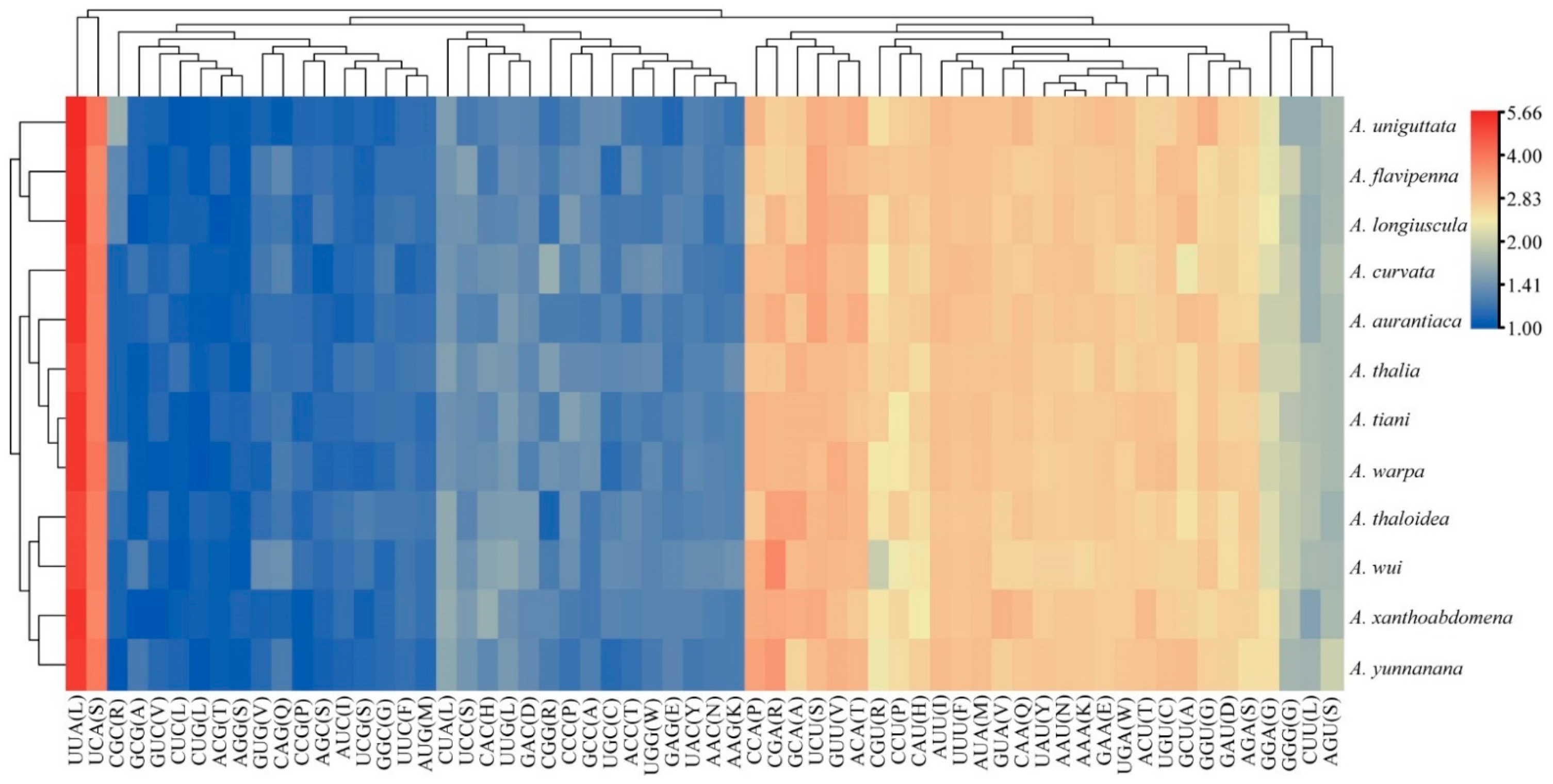
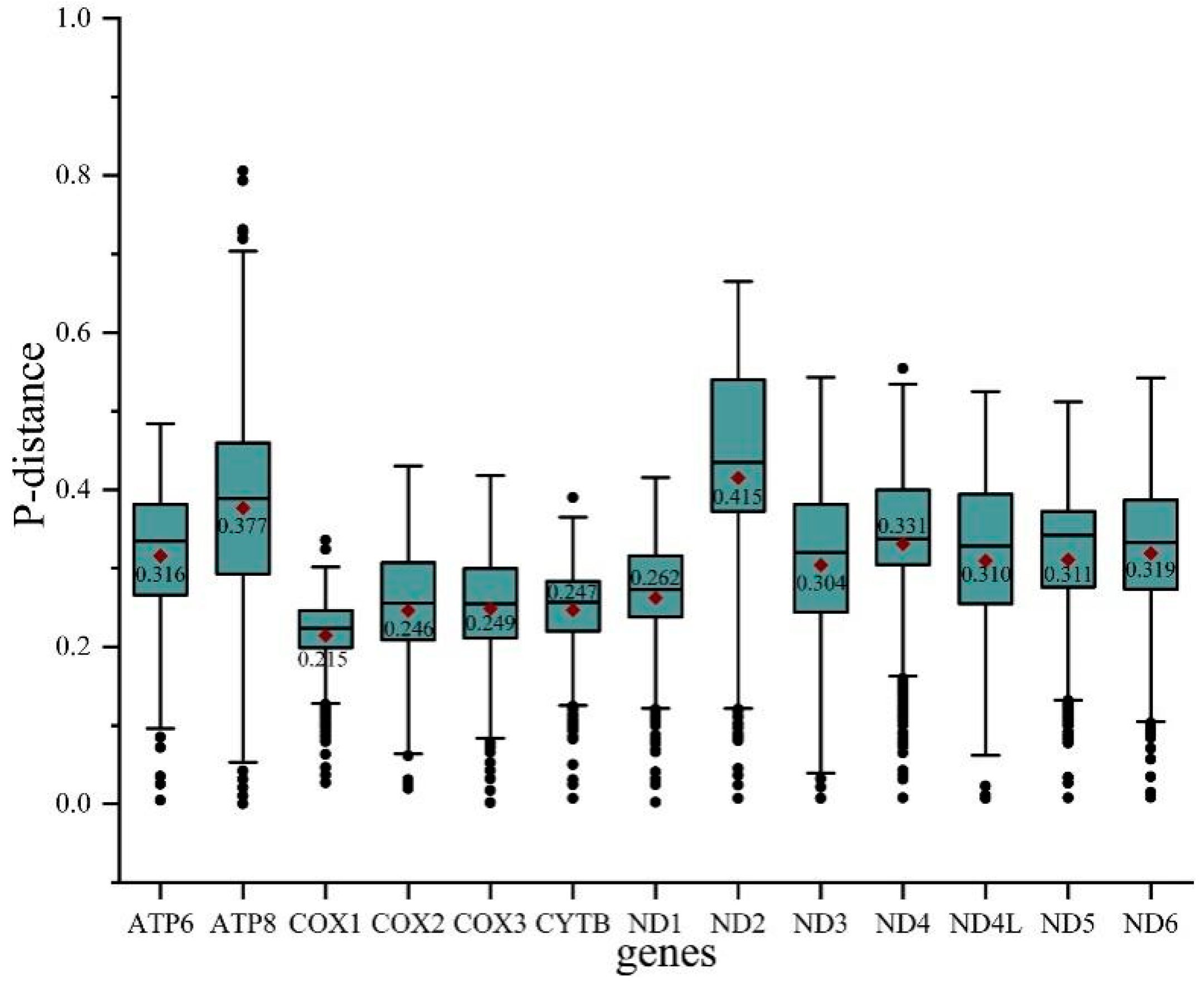
3.2. Protein-Coding Genes and Codon Usage
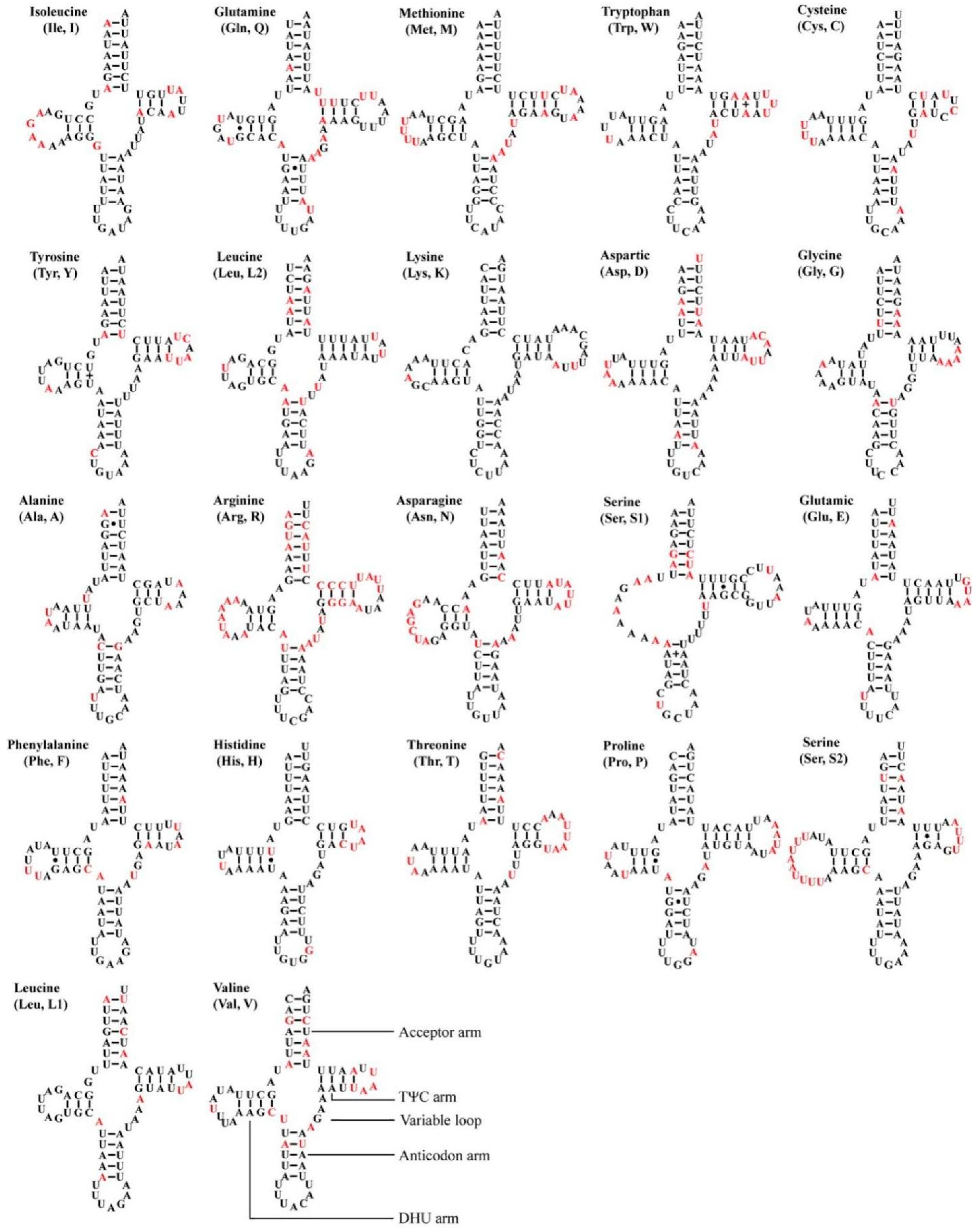
3.3. Transfer and Ribosomal RNA Genes
3.4. Control Region
3.5. Phylogenetic Analyses
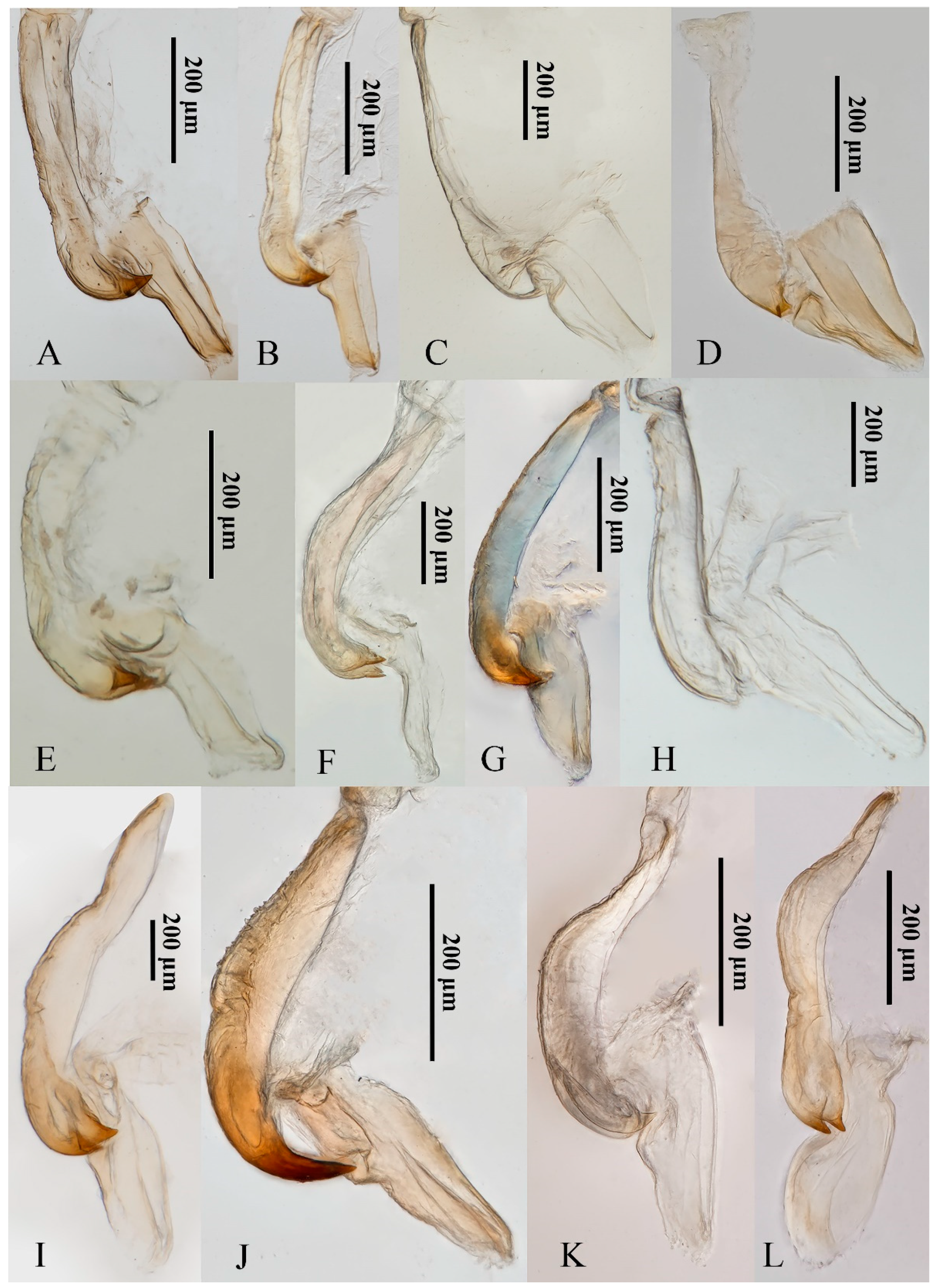
3.6. Morphology
| Key to the 12 Atkinsoniella species | |
| 1. | Basal portion of crown without median black spots ........................................................... 2 |
| Basal portion of crown with a median black spot ................................................................ 3 | |
| 2. | Male pygofer process without branch ............................................................... A. uniguttata |
| Male pygofer process with a small branch at subapex .......................... A. xanthoabdomena | |
| 3. | Apical portion of crown without black spots, the spot of basal portion V-shaped ............ |
| ..................................................................................................................................... A. curvata | |
| Apical portion of crown with a median black spot, the spot of basal portion not V-shap | |
| ed ............................................................................................................................................... 4 | |
| 4. | Crown with large basal black spot, distinctly larger than the anterior median one ......... |
| ............................................................................................................................................ A. wui | |
| Crown with small basal black spot, as big as or smaller than the anterior median one | |
| .................................................................................................................................................... 5 | |
| 5. | Scutellum with a black spot in each basal angle .................................................................. 6 |
| Scutellum without black spots ............................................................................................... 7 | |
| 6 | Male pygofer process with apical 1/3 constricted and contorted; aedeagus broad and |
| short ........................................................................................................................ A. flavipenna | |
| Male pygofer process normal, with toothed process subapically; aedeagus stout and long, | |
| apex curved dorsad ..................................................................................................... A. thalia | |
| 7. | Male pygofer process with branch or toothed process subapically .................................. 8 |
| Male pygofer process without branch or process ................................................................ 9 | |
| 8. | Male pygofer process with membranous branch; paraphysis curved dorsad from median |
| portion, and with apical 1/3 curved posteroventrally ........................... A. yunnanana | |
| Male pygofer process with toothed process subapically, paraphysis curved not as above | |
| ................................................................................................................................... A. thaloidea | |
| 9. | Male pygofer process foliated and contorted medially .................................................... 10 |
| Male pygofer process banding and not contorted medially ............................................ 11 | |
| 10. | Forewing ivory, base orange-red; male pygofer rounded apically ............ A. longiuscula |
| Forewing orange except apical membrane; male pygofer truncated apically ................... | |
| ................................................................................................................................ A. aurantiaca | |
| 11. | Male pygofer process not extending to apex of pygofer; aedeagus with transverse crack |
| at ventral margin medially .......................................................................................... A. tiani | |
| Male pygofer process extending beyond apex of pygofer; aedeagus without transverse | |
| crack at ventral margin medially .............................................................................. A. warpa | |
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dietrich, C.H. Phylogeny of the leafhopper subfamily Evacanthinae with a review of Neotropical species and notes on related groups (Hemiptera: Membracoidea: Cicadellidae). Syst. Entomol. 2004, 29, 455–487. [Google Scholar] [CrossRef]
- Dietrich, C.H. Keys to the families of Cicadomorpha and subfamilies and tribes of Cicadellidae (Hemiptera: Auchenorrhyncha). Fla. Entomol. 2005, 88, 502–517. [Google Scholar] [CrossRef]
- Yang, M.F.; Meng, Z.H.; Li, Z.Z. Hemiptera: Cicadellidae (II): Cicadellinae. Fauna Sinica: Insecta. Vol. 67; Han, X., Jiao, T., Eds.; Science Press: Beijing, China, 2017; ISBN 978-7-03-052950-3. (In Chinese) [Google Scholar]
- Feng, L.; Zhang, Y.L. Two new species in the genus Kolla Distant (Hemiptera: Cicadellidae: Cicadellinae) from China, with DNA barcoding data. Zootaxa 2017, 4250, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Linnavuori, R.; Delong, D.M. Studies of the Neotropical Mileewaninae (Homoptera: Cicadellidae). J. Kansas Entomol. Soc. 1977, 50, 410–421. [Google Scholar]
- Young, D.A. Taxonomic Study of the Cicadellinae (Homoptera: Cicadellidae) Pt. 1: Proconiini; United States National Museum Bulletin 261; Smithsonian Institution Press: Washington, DC, USA, 1968; pp. 1–287. [Google Scholar] [CrossRef]
- Naveed, H.; Zhang, Y.L. Newly recorded leafhoppers of the subfamily Cicadellinae (Hemiptera: Cicadellidae) with description of a new species from Pakistan. Zootaxa 2018, 4504, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Redak, R.A.; Purcell, A.H.; Lopes, R.S.; Blua, M.J.; Iii, R.F.M.; Andersen, P.C. The Biology of Xylem Fluid-Feeding Insect Vectors of Xylella Fastidiosa and Their Relation to Disease Epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef]
- Kleina, H.T.; Kudlawiec, K.; Esteves, M.B.; Dalbó, M.A. Settling and feeding behavior of sharpshooter vectors of Xylella fastidiosa on plum genotypes resistant to leaf scald disease. Eur. J. Plant Pathol. 2020, 158, 633–644. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Purcell, A.H. Xylella fastidiosa: Cause of Pierce’s Disease of Grapevine and Other Emergent Diseases. Plant Dis. 2002, 86, 1056–1066. [Google Scholar] [CrossRef] [Green Version]
- Krugner, R.; Sisterson, M.S.; Backus, E.A.; Burbank, L.P.; Redak, R.A. Sharpshooters: A review of what moves Xylella fastidiosa. Austral Entomol. 2019, 58, 248–267. [Google Scholar] [CrossRef] [Green Version]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, D.R. Animal Mitochondrial DNA: Structure and Evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar] [CrossRef] [PubMed]
- Yeo, D.; Puniamoorthy, J.; Ngiam, R.W.J.; Meier, R. Towards holomorphology in entomology: Rapid and cost-effective adult–larva matching using NGS barcodes. Syst. Entomol. 2018, 43, 678–691. [Google Scholar] [CrossRef]
- Sun, C.H.; Liu, H.Y.; Lu, C.H. Five new mitogenomes of Phylloscopus (Passeriformes, Phylloscopidae): Sequence, structure, and phylogenetic analyses. Int. J. Biol. Macromol. 2020, 146, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, V.; Gabrieli, P.; Brandini, S.; Capodiferro, M.R.; Javier, P.A.; Chen, X.G.; Achilli, A.; Semino, O.; Gomulski, L.M.; Malacrida, A.R.; et al. The worldwide spread of the tiger mosquito as revealed by mitogenome haplogroup diversity. Front. Genet. 2016, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.Y.; Hasegawa, H.; Cooley, J.R.; Simon, C.; Yoshimura, J.; Cai, W.; Sota, T.; Li, H. Mitochondrial genomics reveals shared phylogeographic patterns and demographic history among three periodical cicada species groups. Mol. Biol. Evol. 2019, 36, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yang, P.C.; Jiang, F.; Chapuis, M.P.; Shali, Y.; Sword, G.A.; Kang, L. Mitochondrial genomes reveal the global phylogeography and dispersal routes of the migratory locust. Mol. Ecol. 2012, 21, 4344–4358. [Google Scholar] [CrossRef]
- Li, H.; Leavengood, J.M.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.P.; Zhou, X.G.; Cai, W.Z. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171223. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, H.; Liu, G.H.; Wang, W.; James, P.; Colwell, D.D.; Tran, A.; Gong, S.Y.; Cai, W.Z.; Shao, R.F. Mitochondrial genome fragmentation unites the parasitic lice of Eutherian mammals. Syst. Biol. 2019, 68, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Shao, R.F.; Song, N.; Song, F.; Jiang, P.; Li, Z.H.; Cai, W.Z. Higher-level phylogeny of paraneopteran insects inferred from mitochondrial genome sequences. Sci. Rep. 2014, 5, 8527. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Q.; Li, H.; Song, F.; Zhao, Y.S.; Wilson, J.J.; Cai, W.Z. Higher-level phylogeny and evolutionary history of Pentatomomorpha (Hemiptera: Heteroptera) inferred from mitochondrial genome sequences. Syst. Entomol. 2019, 44, 810–819. [Google Scholar] [CrossRef]
- Chen, L.; Wahlberg, N.; Liao, C.Q.; Wang, C.B.; Ma, F.Z.; Huang, G.H. Fourteen complete mitochondrial genomes of butterflies from the genus Lethe (Lepidoptera, Nymphalidae, Satyrinae) with mitogenome-based phylogenetic analysis. Genomics 2020, 112, 4435–4441. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.P.; Zhang, X.Y.; Li, Y.; Shang, Y.J.; Chen, S.; Wang, S.W.; Qu, Y.H.; Cai, J.F.; Guo, Y.D. Comparative analysis of mitochondrial genomes among the subfamily Sarcophaginae (Diptera: Sarcophagidae) and phylogenetic implications. Int. J. Biol. Macromol. 2020, 161, 214–222. [Google Scholar] [CrossRef]
- Curole, J.P.; Kocher, T.D. Mitogenomics: Digging deeper with complete mitochondrial genomes. Trends Ecol. Evol. 1999, 14, 394–398. [Google Scholar] [CrossRef]
- Qin, J.; Zhang, Y.Z.; Zhou, X.; Kong, X.B.; Wei, S.J.; Ward, R.D.; Zhang, A.B. Mitochondrial phylogenomics and genetic relationships of closely related pine moth (Lasiocampidae: Dendrolimus) species in China, using whole mitochondrial genomes. BMC Genom. 2015, 16, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, L.; Gao, J.F.; Liu, G.H.; Fu, X.; Su, X.; Yue, D.M.; Gao, Y.; Zhang, Y.; Wang, C.R. The complete mitochondrial genome of Metorchis orientalis (Trematoda: Opisthorchiidae): Comparison with other closely related species and phylogenetic implications. Infect. Genet. Evol. 2016, 39, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [Green Version]
- Meng, G.L.; Li, Y.Y.; Yang, C.T.; Liu, S.L. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rijk, P.; Wuyts, J.; De Wachter, R. Rna Viz 2: An improved representation of RNA secondary structure. Bioinformatics 2003, 19, 299–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6 : Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Le, S.; Li, Y.; Hu, F.Q. SeqKit: A cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kück, P.; Longo, G.C. FASconCAT-G: Extensive functions for multiple sequence alignment preparations concerning phylogenetic studies. Front. Zool. 2014, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partitionfinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, H.X.; Yu, X.F.; Yang, M.F. Characterization of Two Complete Mitochondrial Genomes of Atkinsoniella (Hemiptera: Cicadellidae: Cicadellinae) and the Phylogenetic Implications. Insects 2021, 12, 338. [Google Scholar] [CrossRef]
- Yu, P.F.; Li, Q.; Wang, M.X.; Cheng, P.; Lin, C.; Han, B.Y. Analysis of Complete Mitochondrial Genome and Phylogenetic Relationship of Bothrogonia ferruginea. J. Agric. Biotechnol. 2019, 27, 1246–1258. (In Chinese) [Google Scholar] [CrossRef]
- Xu, X.L.; Yan, B.; Yu, X.F.; Yang, M.F. The complete mitochondrial genome of Bothrogonia qiongana (Hemiptera: Cicadellidae) with phylogenetic analyses. Mitochondrial DNA Part B Resour. 2020, 5, 2750–2751. [Google Scholar] [CrossRef]
- Xu, X.L.; Yan, B.; Yu, X.F.; Yang, M.F. The complete mitochondrial genomes of Bothrogonia tongmaiana and Bothrogonia yunana (Hemiptera: Cicadellidae) from China. Mitochondrial DNA Part B Resour. 2021, 6, 613–614. [Google Scholar] [CrossRef]
- Song, N.; Zhang, H.; Zhao, T. Insights into the phylogeny of Hemiptera from increased mitogenomic taxon sampling. Mol. Phylogenet. Evol. 2019, 137, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.K.; Li, H.X.; Yu, X.F.; Yang, M.F. Complete mitochondrial genome sequence of Cicadella viridis (Hemiptera: Cicadellidae: Cicadellinae). Mitochondrial DNA Part B Resour. 2019, 4, 1287–1288. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.K.; Yang, M.F.; Yu, X.F. The mitochondrial genome of Cofana yasumatsui (Hemiptera: Cicadellidae: Cicadellinae). Mitochondrial DNA Part B Resour. 2020, 5, 1075–1076. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.F.; Wu, G.H.; Han, B.Y. Sequencing and analysis of the mitochondrial genome of Kolla paulula (Walker) (Hemiptera: Cicadellidae). J. Anhui Agric. Univ. 2017, 44, 874–881. [Google Scholar]
- Wang, J.J.; Li, H.; Dai, R.H. Complete mitochondrial genome of Taharana fasciana (Insecta, Hemiptera: Cicadellidae) and comparison with other Cicadellidae insects. Genetica 2017, 145, 593–602. [Google Scholar] [CrossRef]
- Wang, J.J.; Wu, Y.F.; Dai, R.H.; Yang, M.F. Comparative mitogenomes of six species in the subfamily Iassinae (Hemiptera: Cicadellidae) and phylogenetic analysis. Int. J. Biol. Macromol. 2020, 149, 1294–1303. [Google Scholar] [CrossRef]
- Du, Y.; Dietrich, C.H.; Dai, W. Complete mitochondrial genome of Macrosteles quadrimaculatus (Matsumura) (Hemiptera: Cicadellidae: Deltocephalinae) with a shared tRNA rearrangement and its phylogenetic implications. Int. J. Biol. Macromol. 2019, 122, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Dong, Y.L.; Qiao, P.P.; Yang, Z.F. Complete Mitogenomic Structure and Phylogenetic Implications of the Genus Ostrinia (Lepidoptera: Crambidae). Insects 2020, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.L.; Yu, T.H.; Zhang, Y.L. Characterization of the Complete Mitochondrial Genome of Drabescus ineffectus and Roxasellana stellata (Hemiptera: Cicadellidae: Deltocephalinae: Drabescini) and Their Phylogenetic Implications. Insects 2020, 11, 534. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, Y. Characterization of two complete mitochondrial genomes of Ledrinae (Hemiptera: Cicadellidae) and phylogenetic analysis. Insects 2020, 11, 609. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Dietrich, C.H.; Huang, M. Characterization of the complete mitochondrial genomes of two species with preliminary investigation on phylogenetic status of zyginellini (Hemiptera: Cicadellidae: Typhlocybinae). Insects 2020, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Godeiro, N.N.; Pacheco, G.; Liu, S.; Gioia Cipola, N.; Berbel-Filho, W.M.; Zhang, F.; Gilbert, M.T.P.; Bellini, B.C. Phylogeny of Neotropical Seirinae (Collembola, Entomobryidae) based on mitochondrial genomes. Zool. Scr. 2020, 49, 329–339. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, H.; Ge, X.; Yang, G.; Xie, G.; Yang, Y. A Mitochondrial Genome Phylogeny of Cleridae (Coleoptera, Cleroidea). Insects 2022, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome coxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270 (Suppl. 1), S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Taleb, M.; Tail, G.; Açıkgöz, H.N. DNA barcoding of Stearibia nigriceps (Meigen) and Piophila casei (Linnaeus) (Diptera: Piophilidae) from Algeria and the first African report of Stearibia nigriceps. Int. J. Legal Med. 2020, 134, 895–902. [Google Scholar] [CrossRef]
- Ma, J.; Liu, J.; Shen, Y.; Fan, Z.; Yue, B.; Zhang, X. Population genetic structure and intraspecific genetic distance of Periplaneta americana (Blattodea: Blattidae) based on mitochondrial and nuclear DNA markers. Ecol. Evol. 2019, 9, 12928–12939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.H.; Cui, B.Y.; Li, Z.H.; Jiang, F.; Yang, Q.Q.; Kucerova, Z.; Stejskal, V.; Opit, G.; Cao, Y.; Li, F.J. The establishment of species-specific primers for the molecular identification of ten stored-product psocids based on ITS2 rDNA. Sci. Rep. 2016, 6, 21022. [Google Scholar] [CrossRef]
- Haiyan, Z.; Guanghui, C.; Abduklimu, G.; Xiuying, Z.; Yutao, W. Cicadellidae in Some Areas of Xinjiang: Molecular Identification Based on mtDNA COI Gene. Chin. Agric. Sci. Bull. 2021, 37, 119–129. (In Chinese) [Google Scholar]
- Wang, X.Y.; Wang, J.J.; Fan, Z.H.; Dai, R.H. Complete mitogenome of Olidiana ritcheriina (Hemiptera: Cicadellidae) and phylogeny of Cicadellidae. PeerJ 2019, 7, e8072. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Yang, M.F.; Dai, R.H.; Li, H.; Wang, X.Y. Characterization and phylogenetic implications of the complete mitochondrial genome of Idiocerinae (Hemiptera: Cicadellidae). Int. J. Biol. Macromol. 2018, 120, 2366–2372. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Li, D.F.; Li, H.; Yang, M.F.; Dai, R.H. Structural and phylogenetic implications of the complete mitochondrial genome of Ledra auditura. Sci. Rep. 2019, 9, 15746. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Huang, M.; Zhang, Y.L. Structural features and phylogenetic implications of 11 new mitogenomes of Typhlocybinae (Hemiptera: Cicadellidae). Insects 2021, 12, 678. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.H.; Zhang, Y.L. Two complete mitochondrial genomes of Mileewinae (Hemiptera: Cicadellidae) and a phylogenetic analysis. Insects 2021, 12, 668. [Google Scholar] [CrossRef]
- He, H.; Yang, M. Characterization of the leafhopper mitogenome of Mileewa alara (Hemiptera: Cicadellidae: Mileewinae) and its phylogenetic analysis. Mitochondrial DNA Part B Resour. 2021, 6, 1265–1266. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yang, M. Characterization and phylogenetic analysis of the mitochondrial genome of Mileewa ponta (Hemiptera: Cicadellidae: Mileewinae). Mitochondrial DNA Part B Resour. 2020, 5, 2994–2995. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Species | Size (bp) | A + T (%) | Accession Number | Reference |
|---|---|---|---|---|---|
| Cicadellinae | Atkinsoniella aurantiaca | 15,394 | 79.2 | OL677863 | This Study |
| Cicadellinae | Atkinsoniella curvata | 15,988 | 79.3 | OL677864 | This Study |
| Cicadellinae | Atkinsoniella flavipenna | 15,671 | 79.1 | OL677864 | This Study |
| Cicadellinae | Atkinsoniella grahami | 15,621 | 78.6 | MW533712 | [49] |
| Cicadellinae | Atkinsoniella longiuscula | 15,815 | 79.2 | OL677864 | This Study |
| Cicadellinae | Atkinsoniella thalia | 15,034 | 77.8 | OL677864 | This Study |
| Cicadellinae | Atkinsoniella thaloidea | 15,571 | 78.2 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella tiani | 15,841 | 79.2 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella uniguttata | 15,784 | 79.8 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella warpa | 15,852 | 79.0 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella wui | 15,159 | 77.7 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella xanthoabdomena | 15,463 | 78.9 | OL677868 | This Study |
| Cicadellinae | Atkinsoniella xanthonota | 15,895 | 78.4 | MW533713 | [49] |
| Cicadellinae | Atkinsoniella yunnanana | 15,875 | 79.0 | OL677874 | This Study |
| Cicadellinae | Bothrogonia ferruginea | 15,262 | 76.5 | KU167550 | [50] |
| Cicadellinae | Bothrogonia qiongana | 15,788 | 76.9 | NC_049894 | [51] |
| Cicadellinae | Bothrogonia tongmaiana | 15,539 | 76.5 | MT500857 | [52] |
| Cicadellinae | Bothrogonia yunana | 15,585 | 76.4 | NC_049896 | [52] |
| Cicadellinae | Cicadella sp. | 15,131 | 78.2 | MK251092 | [53] |
| Cicadellinae | Cicadella viridis | 13,461 | 78.1 | KY752061 | Unpublished |
| Cicadellinae | Cicadella viridis | 15,891 | 78.8 | MK335936 | [54] |
| Cicadellinae | Cofana unimaculata | 14,865 | 77.6 | MK251095 | [53] |
| Cicadellinae | Cofana yasumatsui | 15,019 | 77.2 | NC_049087 | [55] |
| Cicadellinae | Cuerna septentrionalis | 12,420 | 77.1 | MK251107 | [53] |
| Cicadellinae | Draeculacephala crassicornis | 15,214 | 76.9 | MK251091 | [53] |
| Cicadellinae | Draeculacephala sp. | 12,400 | 75.5 | MK251110 | [53] |
| Cicadellinae | Homalodisca sp. | 14,921 | 78.7 | MK251090 | [53] |
| Cicadellinae | Homalodisca vitripennis | 15,304 | 78.4 | NC_006899 | Unpublished |
| Cicadellinae | Kolla paulula | 13,579 | 73.3 | KY680277 | [56] |
| Cicadellinae | Paracrocampsa sp. | 14,749 | 75.6 | MK251085 | [53] |
| Coelidiinae | Taharana fasciana | 15,161 | 77.9 | NC_036015 | [57] |
| Iassinae | Iassus dorsalis | 15,176 | 80.1 | NC_046066 | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Li, H.-X.; Yu, X.-F.; Yang, M.-F. Comparative Analysis of Mitochondrial Genomes among Twelve Sibling Species of the Genus Atkinsoniella Distant, 1908 (Hemiptera: Cicadellidae: Cicadellinae) and Phylogenetic Analysis. Insects 2022, 13, 254. https://doi.org/10.3390/insects13030254
Jiang Y, Li H-X, Yu X-F, Yang M-F. Comparative Analysis of Mitochondrial Genomes among Twelve Sibling Species of the Genus Atkinsoniella Distant, 1908 (Hemiptera: Cicadellidae: Cicadellinae) and Phylogenetic Analysis. Insects. 2022; 13(3):254. https://doi.org/10.3390/insects13030254
Chicago/Turabian StyleJiang, Yan, Hao-Xi Li, Xiao-Fei Yu, and Mao-Fa Yang. 2022. "Comparative Analysis of Mitochondrial Genomes among Twelve Sibling Species of the Genus Atkinsoniella Distant, 1908 (Hemiptera: Cicadellidae: Cicadellinae) and Phylogenetic Analysis" Insects 13, no. 3: 254. https://doi.org/10.3390/insects13030254
APA StyleJiang, Y., Li, H.-X., Yu, X.-F., & Yang, M.-F. (2022). Comparative Analysis of Mitochondrial Genomes among Twelve Sibling Species of the Genus Atkinsoniella Distant, 1908 (Hemiptera: Cicadellidae: Cicadellinae) and Phylogenetic Analysis. Insects, 13(3), 254. https://doi.org/10.3390/insects13030254