
The Phylogenetic Relationships of the Fanniidae within the Muscoid Grade (Diptera: Calyptrata) Based on the Musculature of the Male Terminalia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
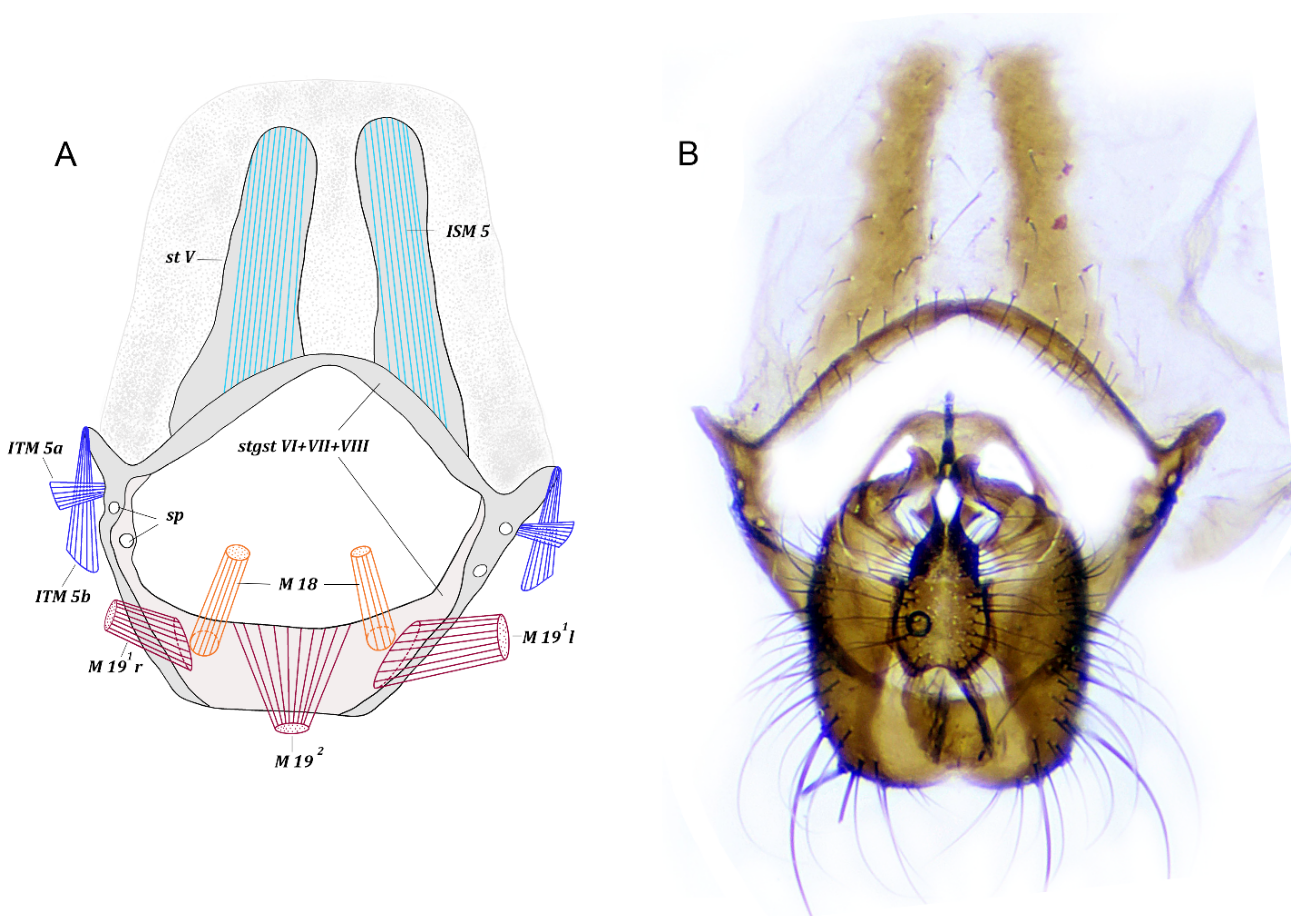
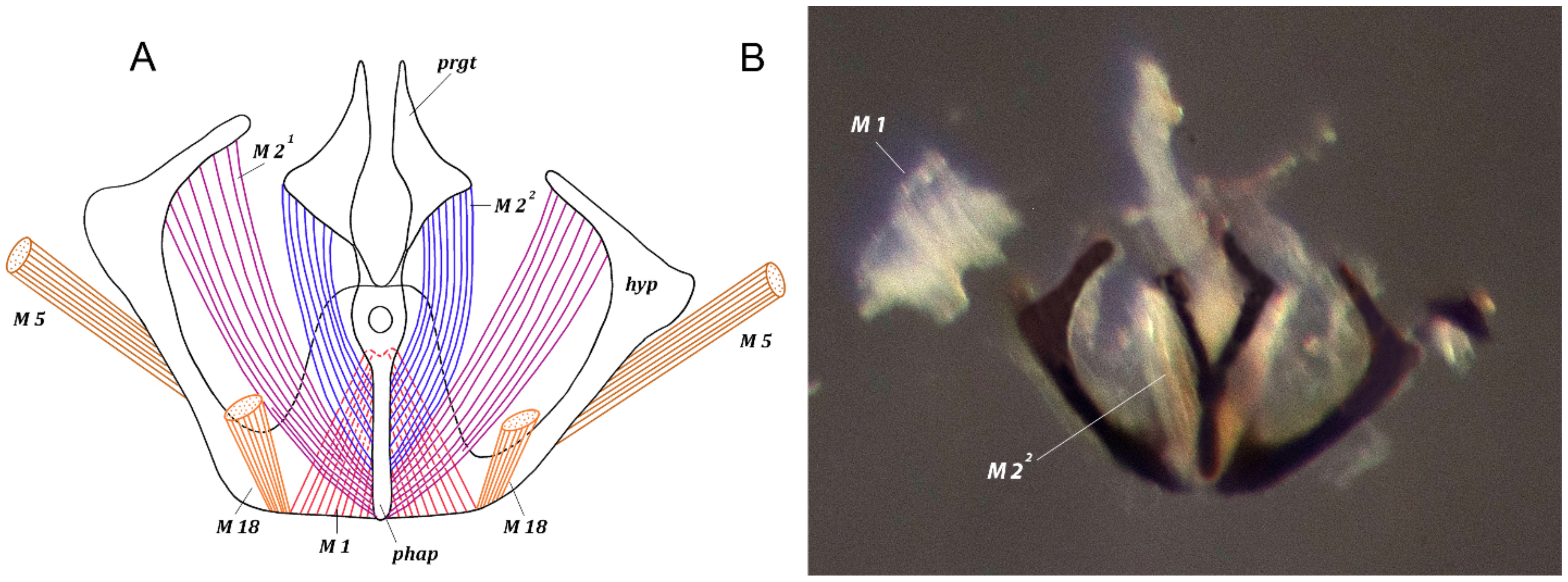
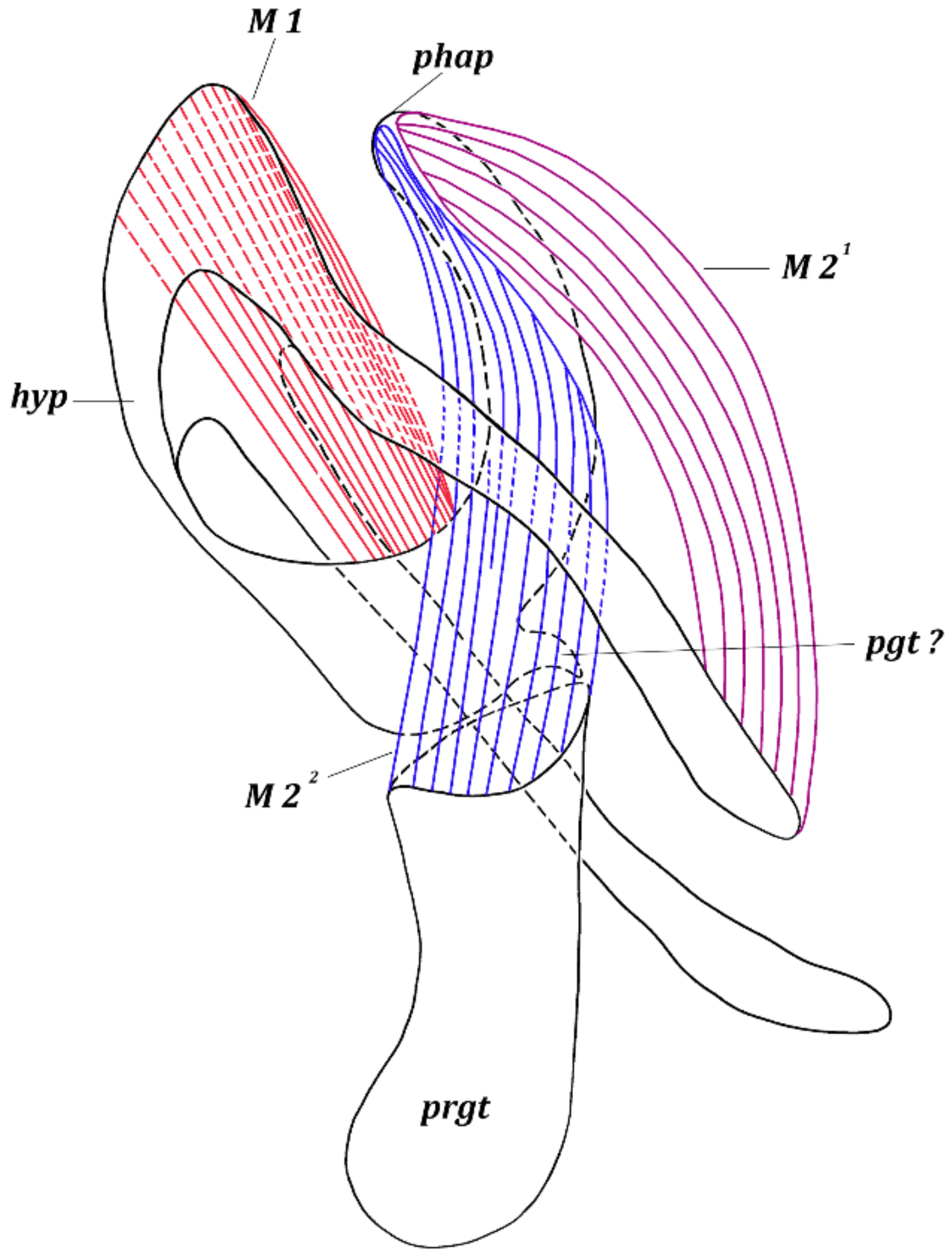
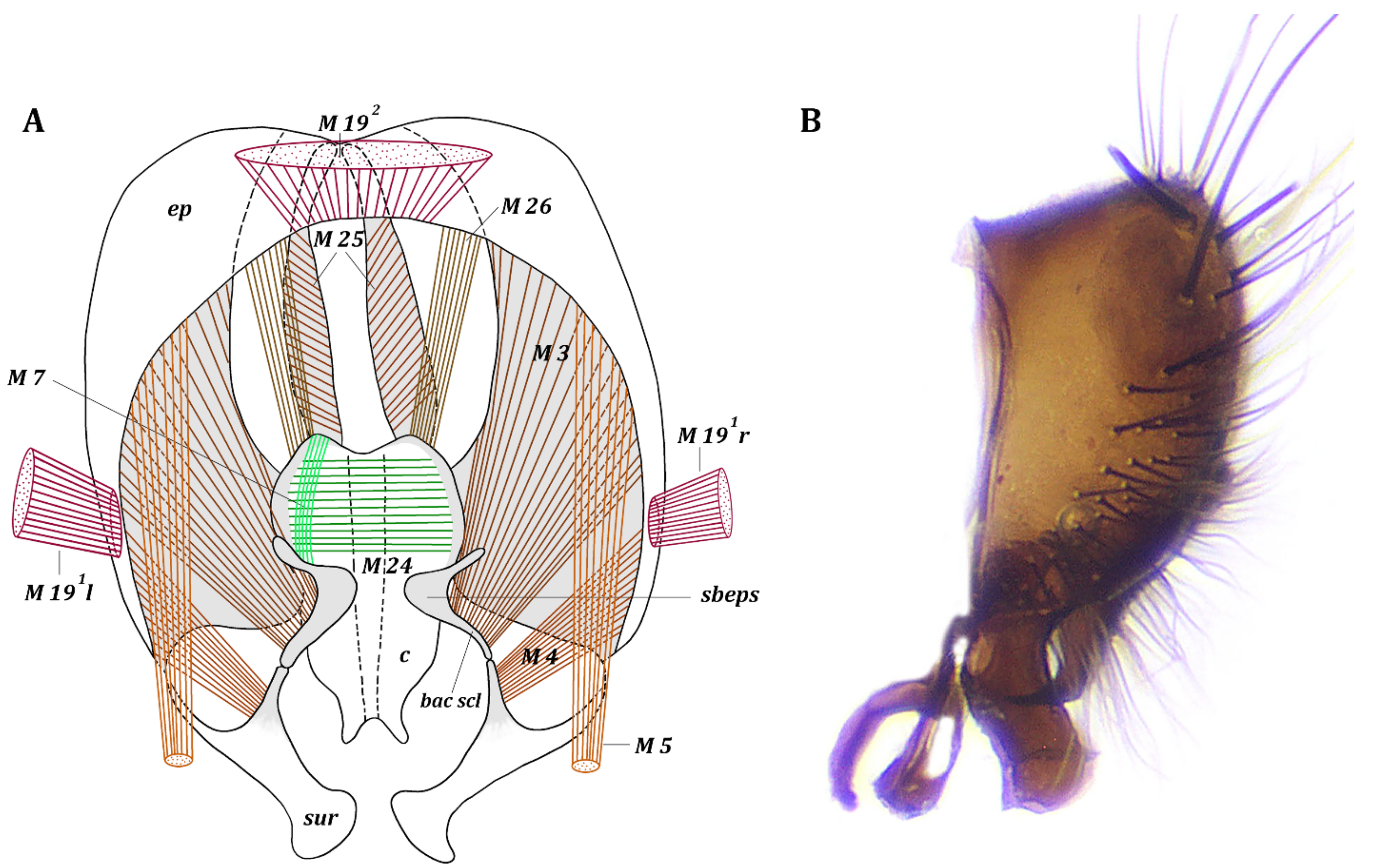
3. Results
3.1. Fannia Subpellucens (Zetterstedt, 1845)
3.1.1. Material Examined
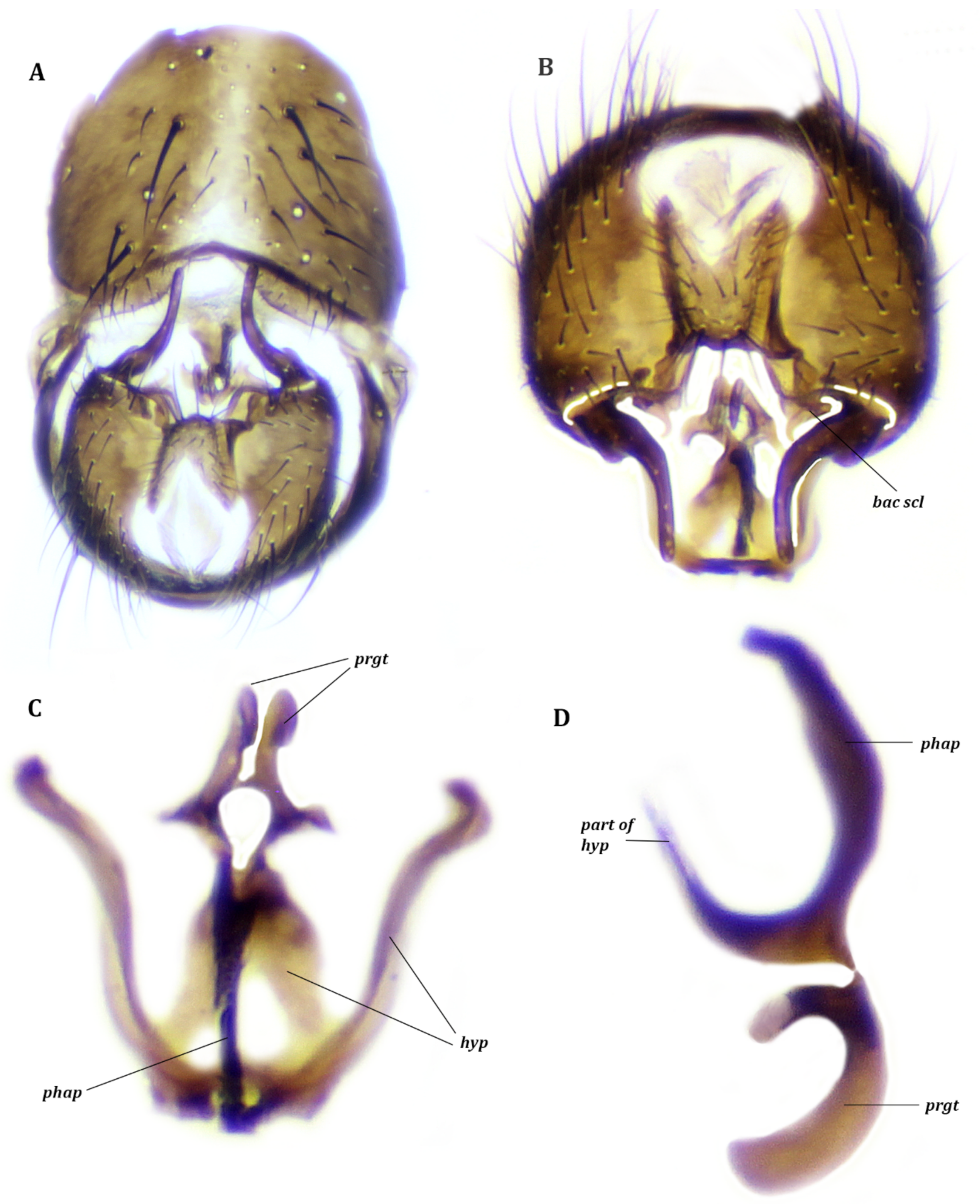
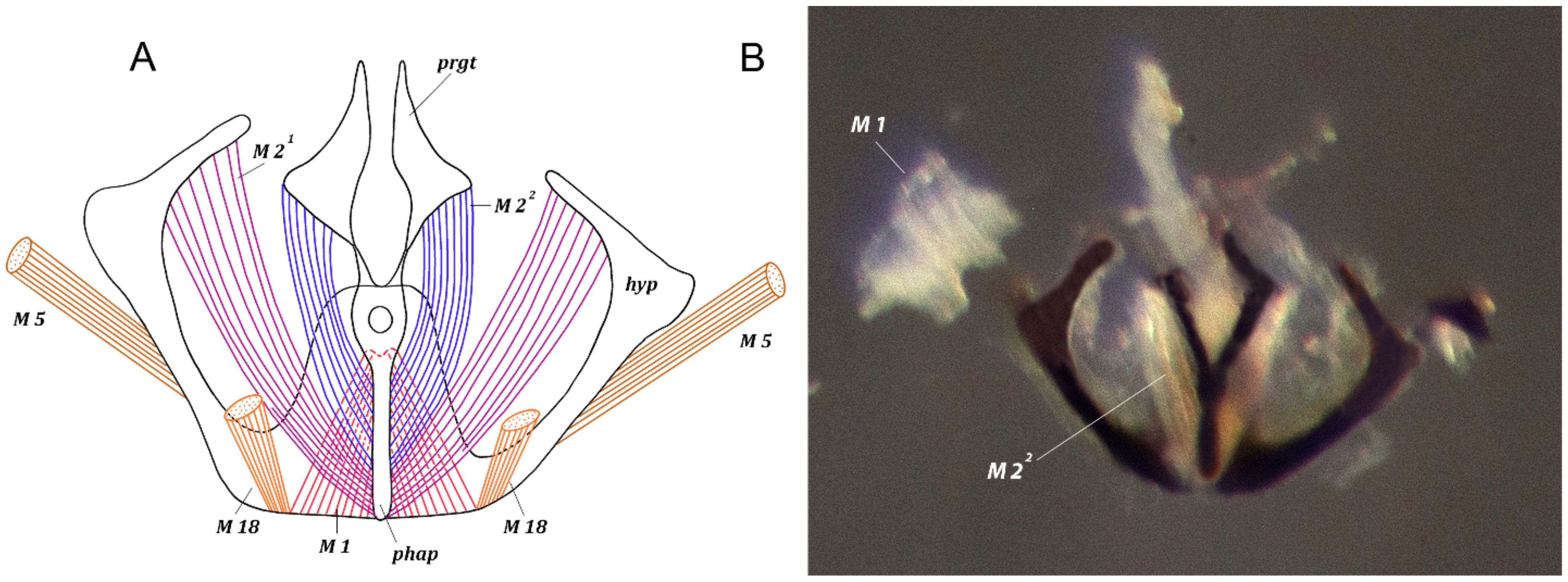
3.1.2. Description
3.2. Fannia Canicularis (Linnaeus, 1761)
3.2.1. Material Examined
3.2.2. Remarks
3.3. Fannia Incisurata (Zetterstedt, 1838)
3.3.1. Material Examined
3.3.2. Remarks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pont, A.C. Family Fanniidae. In Contributions to a Manual of Palaearctic Diptera (with Special Reference to Flies of Economic Importance); Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 2000; pp. 447–454. [Google Scholar]
- Domínguez, M.C.; Roig-Juñent, S.A. A phylogeny of the family Fanniidae Schnabl (Insecta: Diptera: Calyptratae) based on adult morphological characters, with special reference to the Austral species of the genus Fannia. Invertebr. Syst. 2008, 22, 563–587. [Google Scholar] [CrossRef]
- Domínguez, M.C.; Pont, A.C. Fanniidae (Insecta: Diptera). In Fauna of New Zealand; Crosby, T.K., Ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2014; Volume 71, 91p. [Google Scholar]
- Chillcott, J.G. A Revision of the Nearctic Species of Fanniinae (Diptera: Muscidae); Canadian Ent.; Entomological Society of Canada: Ottawa, ON, Canada, 1961; Volume 92, 295p. [Google Scholar]
- Pont, A.C. Observations on anthophilous Muscidae and other Diptera (Insecta) in Abisko National Park. J. Nat. Hist. 1993, 27, 631–643. [Google Scholar] [CrossRef]
- Rozkošný, R.; Gregor, F.; Pont, A.C. The European Fanniidae (Diptera); Acta Scientiarum Naturalium Academiae Scientiarum Bohemicae: Brno, Czech Republic, 1997; Volume 31, 80p. [Google Scholar]
- Pont, A.C. Muscidae (house-flies, stable-flies, etc.). In Insects and Other Arthropods of Medical Importance; Smith, K.G.V., Ed.; British Museum (Natural History): London, UK, 1973; pp. 251–269. [Google Scholar]
- Smith, K.G.V. A Manual of Forensic Entomology; British Museum (Natural History): London, UK, 1986; 205p. [Google Scholar]
- Greenberg, B. Flies and Disease; Princeton University Press: Princeton, NJ, USA, 1971; Volume 1, 856p. [Google Scholar]
- Grzywacz, A.; Wyborska, D.; Piwczyński, M. DNA barcoding allows identification of European Fanniidae (Diptera) of forensic interest. Forensic Sci. Int. 2017, 278, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Verrall, G.H. Note in Séance of 14 October 1891: A reply to Bigot’s earlier (22 July 1891) criticism of his classification of the Anthomyidae. Ann. Soc. Entomol. Fr. 1891, 60, 133–134. [Google Scholar]
- Schnabl, J.; Dziedzicki, H. Die Anthomyiden. In Nova Acta Academiae Caesarea Leopoldina-Carolinae Germanicae Naturae Curiosorum; E. Blochmanni et Filii: Dresden, Germany, 1911; Volume 95, pp. 53–358. [Google Scholar]
- Hennig, W. Muscidae [Part, Lieferung 182]. In Die Fliegen der Palaearktischen Region; Lindner, E., Ed.; Schweizerbart: Stuttgart, Germany, 1955; pp. 1–48. [Google Scholar]
- Huckett, H.C. The Muscidae of Northern Canada, Alaska and Greenland (Diptera); Memoirs of the Entomological Society of Canada: Ottawa, ON, Canada, 1965; Volume 42, 370p. [Google Scholar] [CrossRef]
- Pont, A.C. Family Muscidae. In A Catalogue of the Diptera of the Americas South of the United States; Papavero, N., Ed.; Museum of Zoology of the University of Sao Paulo: São Paulo, Brazil, 1972; pp. 1–111. [Google Scholar]
- Roback, S.S. A classification of the muscoid calyptrate Diptera. Ann. Entomol. Soc. Am. 1951, 44, 327–361. [Google Scholar] [CrossRef]
- Griffiths, G.C.D. The phylogenetic classification of Diptera Cyclorrhapha, with special reference to the structures of the male postabdomen. Ser. Entomol. 1972, 8, 1–340. [Google Scholar]
- Pont, A.C. A revision of Australian Fanniidae (Diptera: Calyptrata). Aust. J. Zool. Suppl. Ser. 1977, 51, 1–60. [Google Scholar] [CrossRef]
- Pont, A.C. Family Fanniidae. In Catalogue of the Diptera of the Afrotropical Region; Crosskey, R.W., Ed.; British Museum (Natural History): London, UK, 1980; pp. 719–720. [Google Scholar]
- Pont, A.C. Family Muscidae. In Catalogue of Palaearctic Diptera. Scathophagidae—Hypodermatidae; Soós, Á., Papp, L., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1986; Volume 11, pp. 57–215. [Google Scholar]
- Pont, A.C. Family Fanniidae. In Catalog of the Diptera of the Australasian and Oceanian Regions; Evenhuis, N.L., Ed.; Bishop Museum Press: Honolulu, HI, USA, 1989; pp. 700–701. [Google Scholar]
- Hennig, W. Vorarbeiten zu einem phylogenetischen System der Muscidae (Diptera: Cyclorrhapha). Stuttg. Beiträge Nat. 1965, 141, 1–100. [Google Scholar]
- Hennig, W. Diptera (Zweiflügler). In Handbuch der Zoologie. Arthropoda—2. Hälfte: Insecra, 2. Teil: Spezielles, 2nd ed.; Helmcke, J.-G., Stark, D., Wermuth, H., Eds.; Walter de Gruyter: Berlin, Germany, 1973; Volume 4, pp. 1–337. [Google Scholar]
- McAlpine, J.F. Phylogeny and classification of the Muscomorpha. In Manual of Nearctic Diptera, Monograph 28; McAlpine, J.F., Ed.; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1989; Volume 3, pp. 1397–1502. [Google Scholar]
- Kutty, S.N.; Meusemann, K.; Bayless, K.M.; Marinho, M.A.T.; Pont, A.C.; Zhou, X.; Misof, B.; Wiegmann, B.M.; Yeates, D.; Cerretti, P.; et al. Phylogenomic analysis of Calyptratae: Resolving a major radiation of Diptera. Cladistics 2019, 35, 605–622. [Google Scholar] [CrossRef]
- Kutty, S.N.; Pape, T.; Pont, A.C.; Wiegmann, B.M.; Meier, R. The Muscoidea (Diptera: Calyptratae) are paraphyletic: Evidence from four mitochondrial and four nuclear genes. Mol. Phylogenet. Evol. 2008, 49, 639–652. [Google Scholar] [CrossRef]
- Kutty, S.N.; Pape, T.; Wiegmann, B.M.; Meier, R. Molecular phylogeny of the Calyptratae (Diptera: Cyclorrhapha) with an emphasis on the superfamily Oestroidea and the position of Mystacinobiidae and McAlpine’s fly. Syst. Entomol. 2010, 35, 614–635. [Google Scholar] [CrossRef]
- Domínguez, M.C.; Roig-Juñent, S.A. Phylogeny and taxonomic revision of the genus Euryomma Stein (Diptera: Calyptratae: Fanniidae). Arthropod Syst. Evol. 2017, 75, 303–326. [Google Scholar]
- Woodley, N.E.; Borkent, A.; Wheeler, T.A. Phylogeny of the Diptera. In Manual of Central American Diptera; Brown, B.V., Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E., Zumbado, M.A., Eds.; National Research Council of Canada: Ottawa, ON, Canada, 2009; Chapter 5; Volume 1, pp. 79–94. [Google Scholar]
- Bernasconi, M.V.; Valsangiacomo, C.; Piffaretti, J.C.; Ward, P.I. Phylogenetic relationships among Muscoidea (Diptera: Calyptratae) based on mitochondrial DNA sequences. Insect Mol. Biol. 2000, 9, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.W.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Peterson, K.J.; Bayless, K.M.; et al. Episodic radiations in the fly tree of life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [Green Version]
- Ovtshinnikova, O.G. Muscles of the male genitalia of Brachycera-Orthorrhapha (Diptera). Tr. Zool. Inst. Akad. Nauk. SSSR 1989, 190, 1–166. (In Russian) [Google Scholar]
- Ovtshinnikova, O.G. On the homology of male genital sclerites of Brachycera Orthorrhapha and Cyclorrhapha (Diptera) based on musculature. Dipterol. Res. 1994, 5, 263–269. [Google Scholar]
- Sinclair, B.J.; Cumming, J.M.; Brooks, S.E. Male terminalia of Diptera (Insecta): A review of evolutionary trends, homology and phylogenetic implications. Insect Syst. Evol. 2013, 44, 373–415. [Google Scholar] [CrossRef]
- Hennig, W. Anthomyiidae [Part, Lieferung 315]. In Die Fliegen der Palaearktischen Region; Lindner, E., Ed.; Schweizerbart: Stuttgart, Germany, 1976; pp. 1–78. [Google Scholar]
- Ovtshinnikova, O.G.; Galinskaya, T.V.; Sorokina, V.S. Musculature of the male abdominal segments and terminalia in Musca autumnalis De Geer, 1776 and Pyrellia rapax (Harris, 1780) (Diptera, Muscidae: Muscini). Entomol. Rev. 2018, 98, 678–689. [Google Scholar] [CrossRef]
- Ovtshinnikova, O.G.; Sorokina, V.S.; Galinskaya, T.V. Musculature of the male abdominal segments and terminalia of Mydaea urbana (Meigen, 1826) and Graphomya maculata (Scopoli, 1763) (Diptera, Muscidae: Mydaeinae). Entomol. Rev. 2019, 99, 628–638. [Google Scholar] [CrossRef]
- Ovtshinnikova, O.G.; Sorokina, V.S. Musculature of the male abdominal segments and terminalia of Spilogona Schnabl, 1911 (Diptera, Muscidae: Coenosiinae). Entomol. Rev. 2020, 100, 637–646. [Google Scholar] [CrossRef]
- Sorokina, V.S.; Ovtshinnikova, O.G. The position of the Azeliinae in the Muscidae (Diptera) based on musculature of the male terminalia. Zookeys 2020, 975, 87–110. [Google Scholar] [CrossRef]
- Sinclair, B.J. Morphology and terminology of Diptera male terminalia. In Contributions to a Manual of Palaearctic Diptera (with Special Reference to Flies of Economic Importance); Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 2000; Volume 1, pp. 53–74. [Google Scholar]
- Cumming, J.M.; Wood, D.M. Adult morphology and terminology. In Manual of Afrotropical Diptera. Introductory Chapters and Keys to Diptera Families; Suricata 4; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, Namibia, 2017; Volume 1, pp. 89–133. [Google Scholar]
- Ovtshinnikova, O.G. Muscles of the male genitalia of Syrphidae (Diptera). In N.A. Kholodkovsky Memorial Lectures. A Report to the 52nd Annual Meeting, 1st April 1999; Sinev, S.Y., Ed.; Russian Entomological Society: St. Petersburg, Russia, 2000; pp. 3–70. (In Russian) [Google Scholar]
- Salzer, R. Konstruktionsanatomische Untersuchung des mannlichen Postabdomens von Calliphora erythrocephala Meigen (Insecta, Diptera). Z. Morphol. Tiere 1968, 63, 155–238. [Google Scholar] [CrossRef]
- Ovtshinnikova, O.G.; Sorokina, V.S. Musculature of the male abdominal segments and terminalia of Tachina nupta Rondani (Diptera: Tachinidae). Acta Zool. 2021, 1–9. [Google Scholar] [CrossRef]
- Schlein, Y.; Theodor, O. On the genitalia of the Pupipara and their homologies with those of Calliphora and Glossina. Parasitology 1971, 63, 331–342. [Google Scholar] [CrossRef]
- Cumming, J.M.; Sinclair, B.J.; Wood, D.M. Homology and phylogenetic implications of male genitalia in Diptera—Eremoneura. Entomol. Scand. 1995, 26, 120–151. [Google Scholar]
- Zaka-ur-Rab, M. Morphology of the male terminalia of Basilia (Paracyclopodia) burmensis (Theodor) (Diptera: Nycteribiidae). Zool. Anz. 1979, 203, 177–181. [Google Scholar]
- Zaka-ur-Rab, M. Morphology of the male genitalia of Hippobosca maculata L. (Diptera: Hippoboscidae). Zool. Anz. 1979, 203, 182–188. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorokina, V.S.; Ovtshinnikova, O.G. The Phylogenetic Relationships of the Fanniidae within the Muscoid Grade (Diptera: Calyptrata) Based on the Musculature of the Male Terminalia. Insects 2022, 13, 210. https://doi.org/10.3390/insects13020210
Sorokina VS, Ovtshinnikova OG. The Phylogenetic Relationships of the Fanniidae within the Muscoid Grade (Diptera: Calyptrata) Based on the Musculature of the Male Terminalia. Insects. 2022; 13(2):210. https://doi.org/10.3390/insects13020210
Chicago/Turabian StyleSorokina, Vera S., and Olga G. Ovtshinnikova. 2022. "The Phylogenetic Relationships of the Fanniidae within the Muscoid Grade (Diptera: Calyptrata) Based on the Musculature of the Male Terminalia" Insects 13, no. 2: 210. https://doi.org/10.3390/insects13020210
APA StyleSorokina, V. S., & Ovtshinnikova, O. G. (2022). The Phylogenetic Relationships of the Fanniidae within the Muscoid Grade (Diptera: Calyptrata) Based on the Musculature of the Male Terminalia. Insects, 13(2), 210. https://doi.org/10.3390/insects13020210