

Comparative Transcriptome Analysis to Reveal Differentially Expressed Cytochrome P450 in Response to Imidacloprid in the Aphid Lion, Chrysoperla zastrowi sillemi (Esben-Petersen)
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
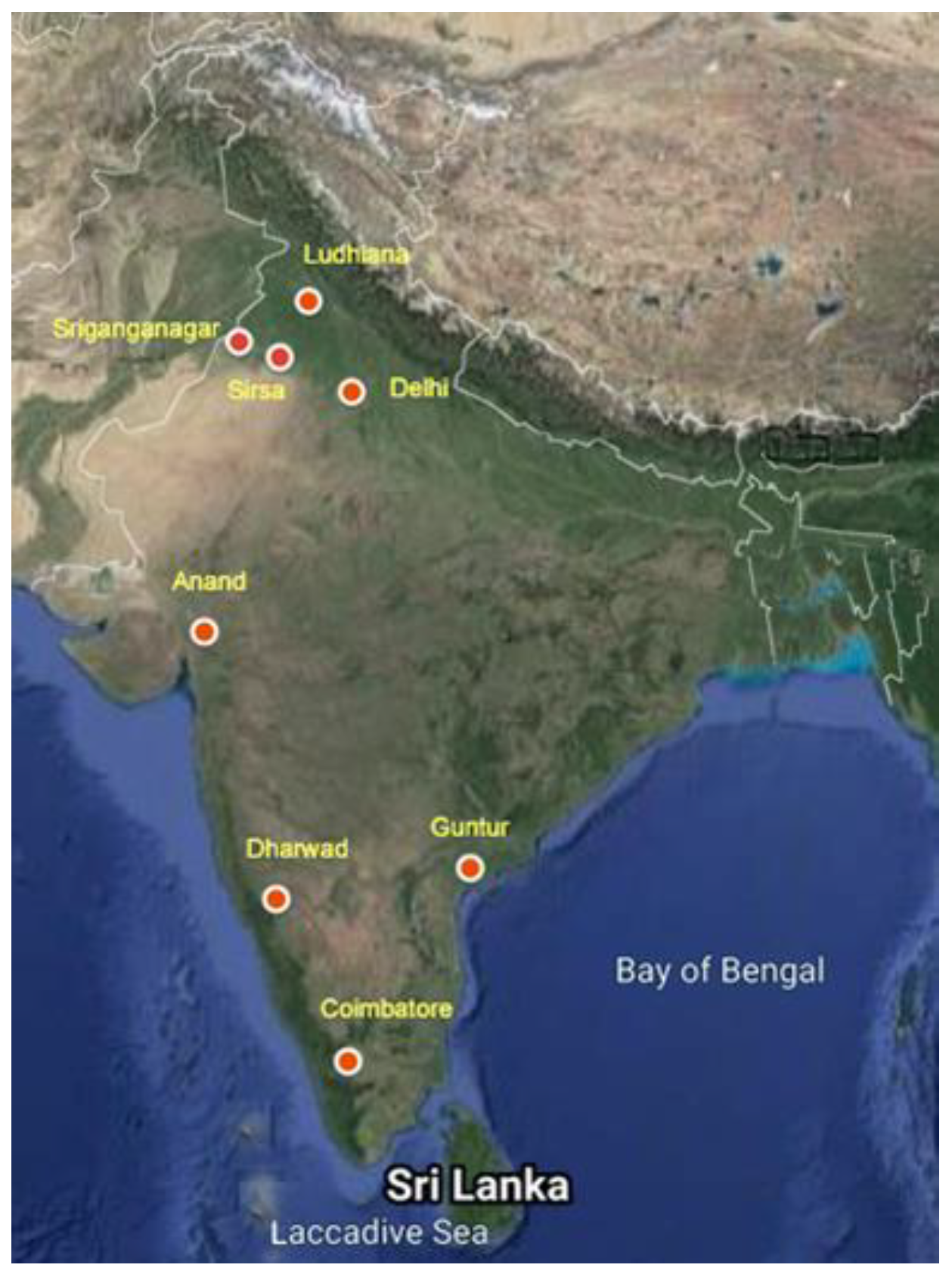
2.1. Insect Collection and Laboratory Maintenance
2.2. Dose Mortality Bioassays
2.3. RNA Isolation, Library Preparation, and Sequencing
2.4. Pre-Processing of Raw Reads, De Novo Assembly, and Quality Assessment
2.5. Transcriptome Annotation
2.6. Transcript Abundance and Differential Gene Expression
2.7. Identification and Phylogenetic Classification of Cytochrome P450 Sequences
2.8. Total RNA Isolation and Reverse Transcription Quantitative Real-Time PCR (qRT-PCR)
3. Results
3.1. Bioassay
3.2. RNA Sequencing and Quality Control
3.3. De novo Transcriptome Assembly and Quality Assessment
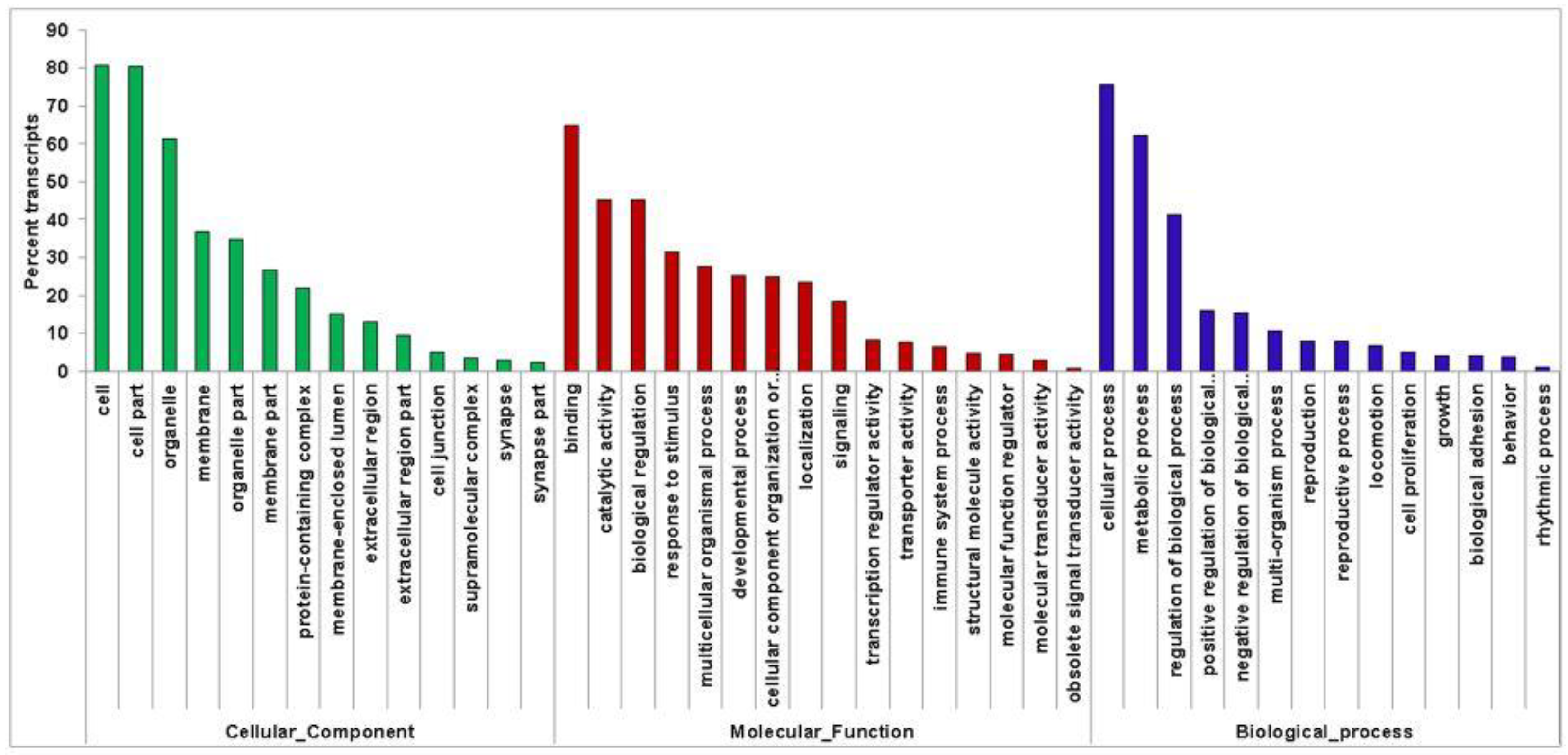
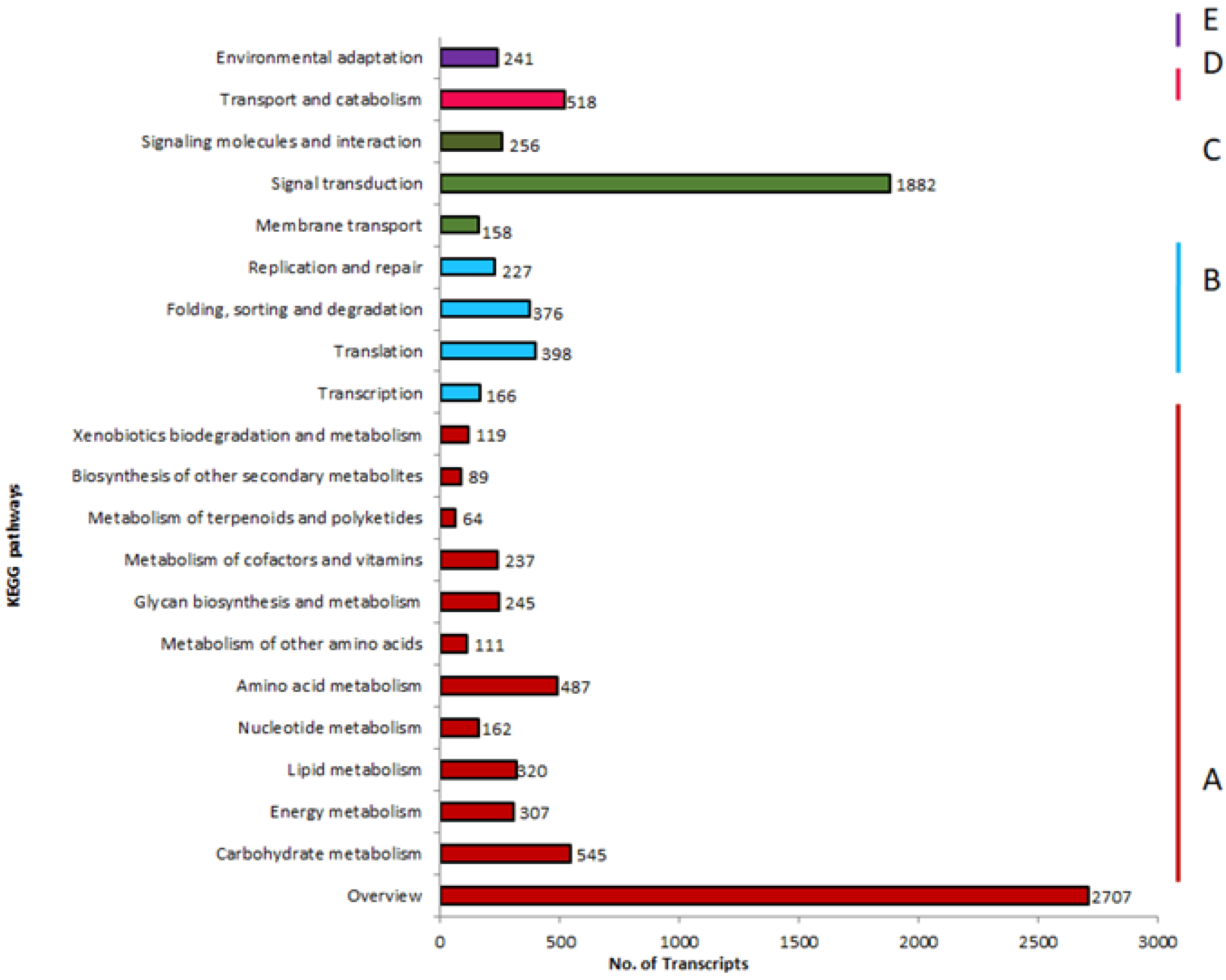
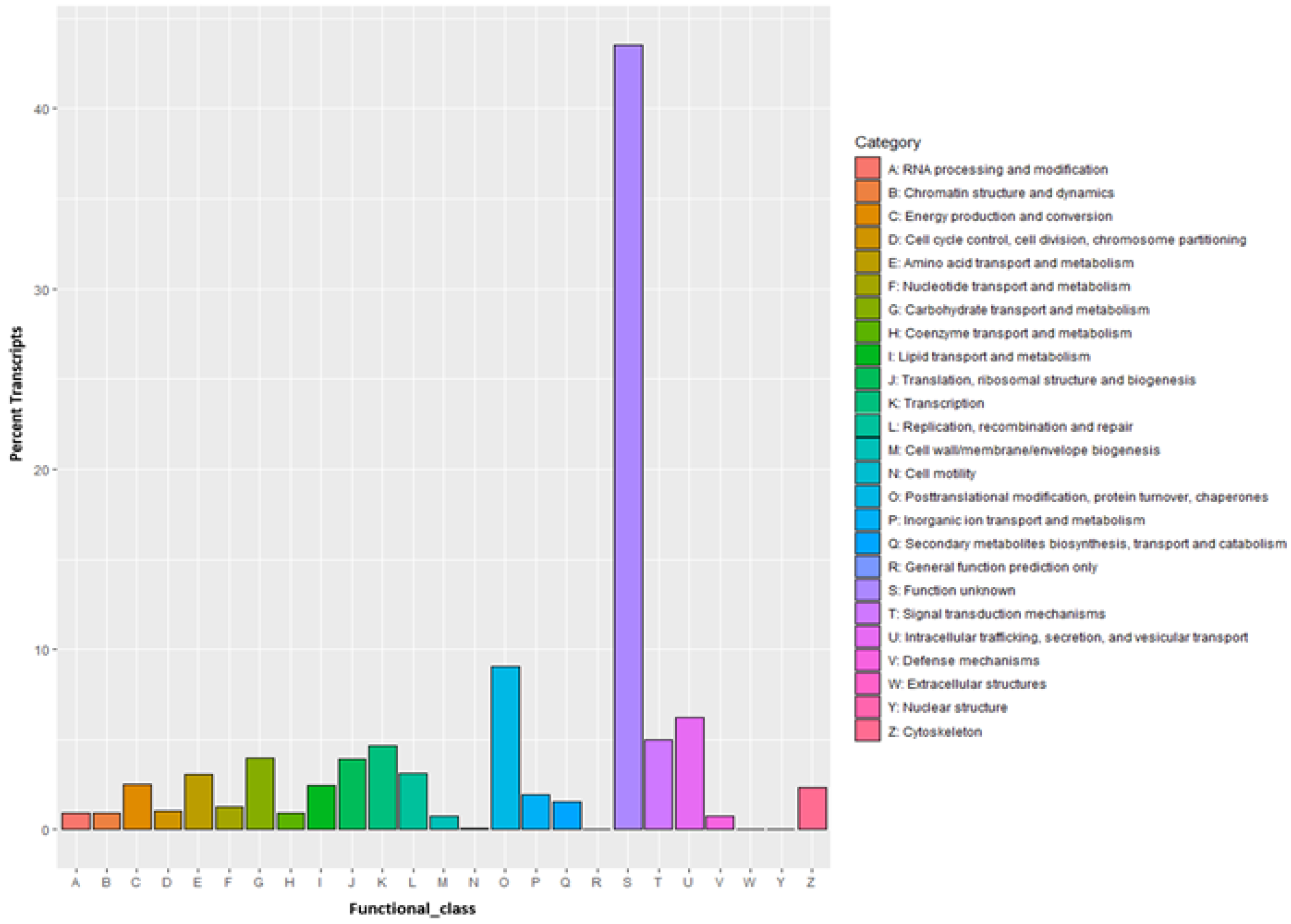
3.4. Transcriptome Annotation
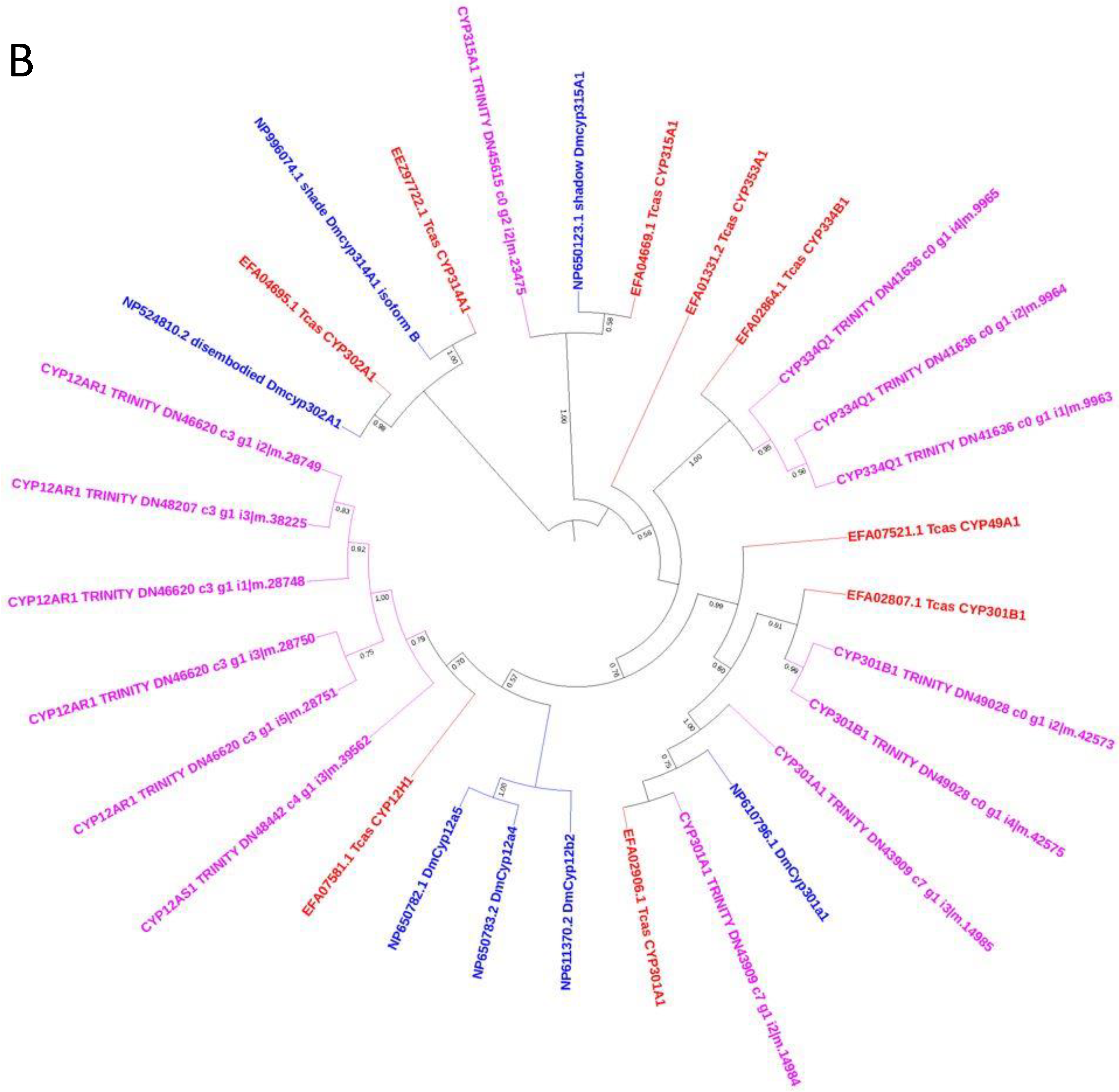
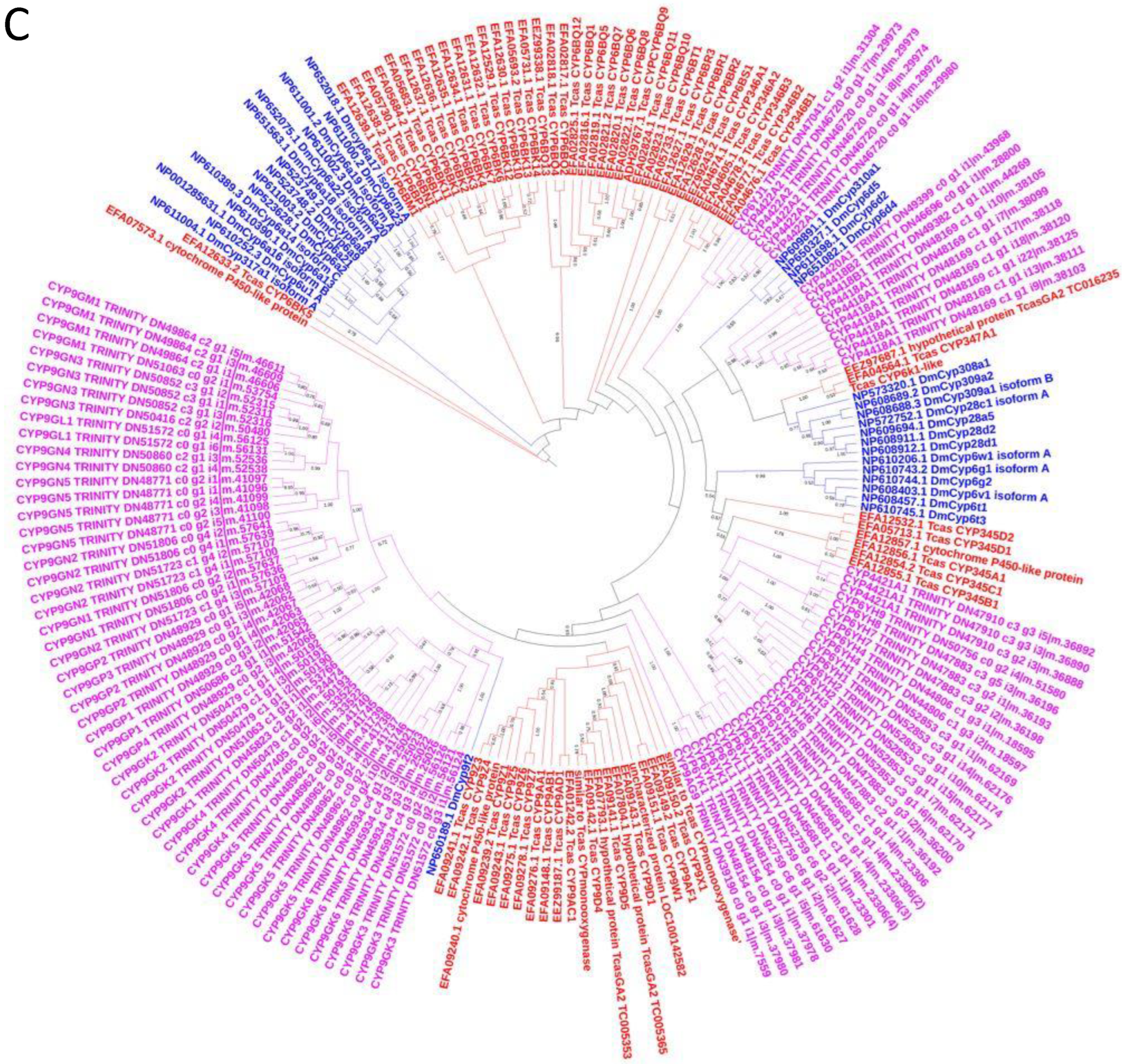
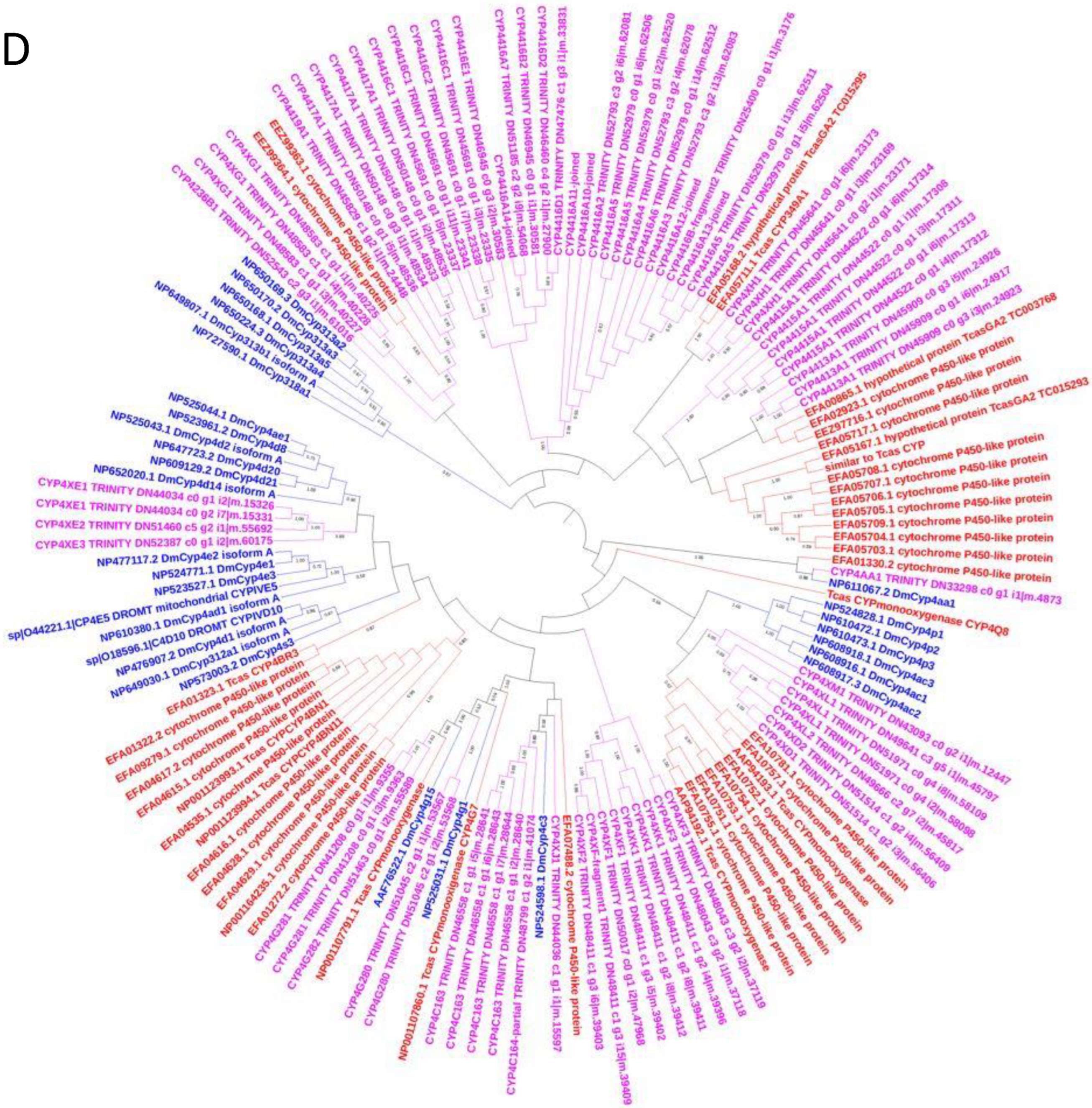
3.5. Phylogenetic Analysis of C. zastrowi sillemi Cytochrome P450 Sequences
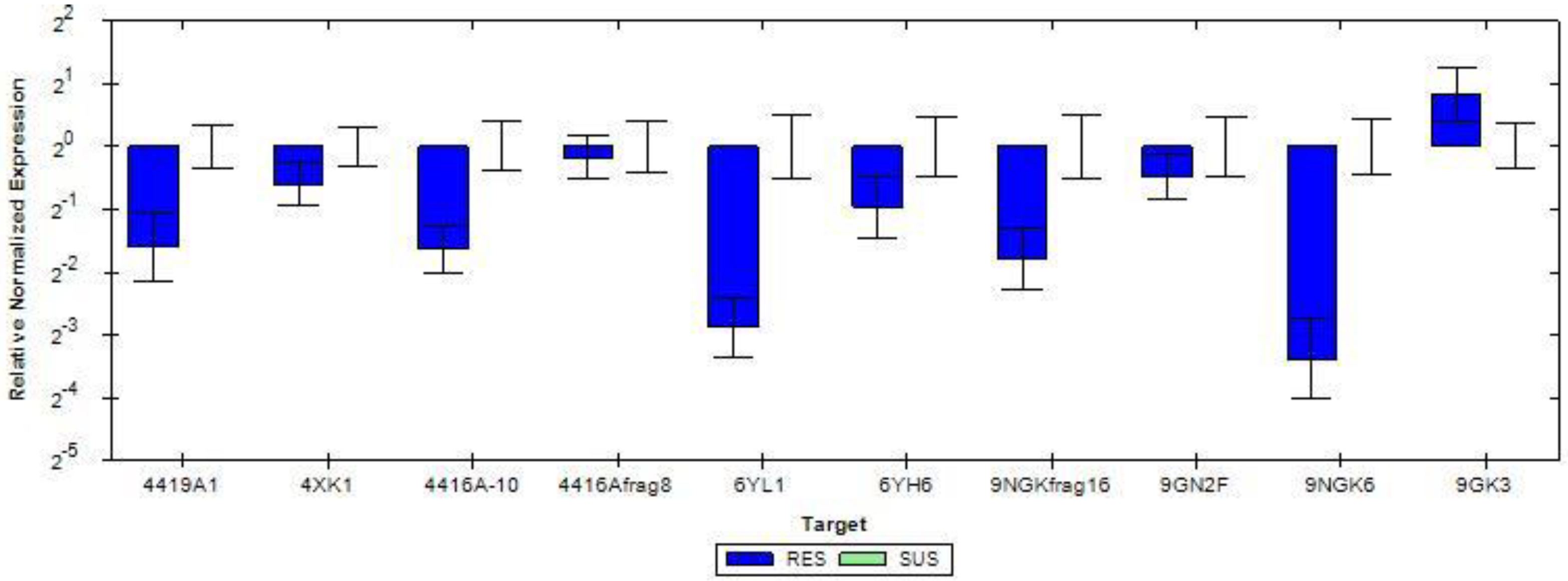
3.6. Differential Gene Expression and Validation by Quantitative Real-Time PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winterton, S.; De Freitas, S. Molecular Phylogeny of the Green Lacewings (Neuroptera: Chrysopidae). Aust. J. Entomol. 2006, 45, 235–243. [Google Scholar] [CrossRef]
- Pappas, M.L.; Broufas, G.D.; Koveos, D.S. Chrysopid Predators and Their Role in Biological Control. J. Entomol. 2011, 8, 301–326. [Google Scholar] [CrossRef]
- Tauber, M.J.; Tauber, C.A.; Daane, K.M.; Hagen, K.S. Commercialization of Predators: Recent Lessons from Green Lacewings (Neuroptera: Chrysopidae: Chrosoperla). Am. Entomol. 2000, 46, 26–38. [Google Scholar] [CrossRef]
- Nordlund, D.A.; Cohen, A.C.; Smith, R.A.; McEwen, P.; New, T.; Whittington, A. Mass-Rearing, Release Techniques, and Augmentation. In Lacewings in the Crop Environment; Cambridge University Press: Cambridge, UK, 2001; pp. 301–319. [Google Scholar] [CrossRef]
- Henry, C.S.; Brooks, S.J.; Johnson, J.B.; Venkatesan, T.; Duelli, P. The Most Important Lacewing Species in Indian Agricultural Crops, Chrysoperla sillemi (Esben-Petersen), is a Subspecies of Chrysoperla zastrowi (Esben-Petersen) (Neuroptera: Chrysopidae). J. Nat. Hist. 2010, 44, 2543–2555. [Google Scholar] [CrossRef]
- Aggarwal, N. Neetan Predatory Efficiency of Cheilomenes sexmaculata (Fabricius) and Chrysoperla zastrowi sillemi (Esben-Petersen) on Cotton mealy bug, Phenacoccus solenopsis tinsley under Laboratory Conditions. Acta Phytopathol. Entomol. Hungarica 2014, 49, 73–81. [Google Scholar] [CrossRef]
- Manjunatha, D.K.; Sharanabasappa, S.E.; Maruthi, M.S. Feeding Potential of Chrysoperla carnea (Steph.) on Different Host. J. Entomol. Zool. Stud. 2018, 6, 1028–1030. [Google Scholar]
- Venkatesan, T.; Jalali, S.K.; Murthy, K.S.; Rabindra, R.J.; Lalitha, Y. Occurrence of Insecticide Resistance in Field Populations of Chrysoperla zastrowi arabica (Neuroptera: Chrysopidae) in India. Indian J. Agric. Sci. 2009, 79, 910–912. [Google Scholar]
- Khan Pathan, A.; Sayyed, A.H.; Aslam, M.; Liu, T.-X.; Razzaq, M.; Ahmed Gillani, W. Resistance to Pyrethroids and Organophosphates Increased Fitness and Predation Potential of Chrysoperla carnae (Neuroptera: Chrysopidae). J. Econ. Entomol. 2010, 103, 823–834. [Google Scholar] [CrossRef]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides Acting on Insect Nicotinic Acetylcholine Receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Bass, C.; Carvalho, R.A.; Oliphant, L.; Puinean, A.M.; Field, L.M.; Nauen, R.; Williamson, M.S.; Moores, G.; Gorman, K. Overexpression of a Cytochrome P450 Monooxygenase, CYP6ER1, is Associated with Resistance to Imidacloprid in the Brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2011, 20, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, W.; Wang, S.; Wu, Q.; Pan, H.; Li, R.; Yang, N.; Liu, B.; Xu, B.; Zhou, X. Two Cytochrome P450 Genes Are Involved in Imidacloprid Resistance in Field Populations of the Whitefly, Bemisia tabaci, in China. Pestic. Biochem. Physiol. 2013, 107, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.; Schoville, S.; Peterson, N.; Huseth, A.S.; Lan, Q.; Groves, R.L. RNA Interference of Three Up-Regulated Transcripts Associated with Insecticide Resistance in an Imidacloprid Resistant Population of Leptinotarsa decemlineata. Pestic. Biochem. Physiol. 2017, 135, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Elzaki, M.E.A.; Miah, M.A.; Wu, M.; Zhang, H.; Pu, J.; Jiang, L.; Han, Z. Imidacloprid Is Degraded by CYP353D1v2, a Cytochrome P450 Overexpressed in a Resistant Strain of Laodelphax striatellus. Pest Manag. Sci. 2017, 73, 1358–1363. [Google Scholar] [CrossRef]
- Nelson, D.R.; Nebert, D.W. Cytochrome P450 (CYP) Gene Superfamily. eLS 2018, 1–19. [Google Scholar] [CrossRef]
- Ronis, M.J.J.; Hodgson, E. Cytochrome P-450 Monooxygenases in Insects. Xenobiotica 1989, 19, 1077–1092. [Google Scholar] [CrossRef]
- Daborn, P.J.; Le Goff, G. The Genetics and Genomics of Insecticide Resistance. TRENDS Genet. 2004, 20, 163–170. [Google Scholar]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular Mechanisms of Metabolic Resistance to Synthetic and Natural Xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Nauen, R.; Denholm, I. Resistance of Insect Pests to Neonicotinoid Insecticides: Current Status and Future Prospects. Arch. insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2005, 58, 200–215. [Google Scholar] [CrossRef]
- Hsu, J.-C.; Lin, Y.-Y.; Chang, C.-C.; Hua, K.-H.; Chen, M.-J.M.; Huang, L.-H.; Chen, C.-Y. Discovery of Organophosphate Resistance-Related Genes Associated with Well-Known Resistance Mechanisms of Plutella xylostella (L.) (Lepidoptera: Plutellidae) by RNA-Seq. J. Econ. Entomol. 2016, 109, 1378–1386. [Google Scholar] [CrossRef]
- Graveley, B.R.; Brooks, A.N.; Carlson, J.W.; Duff, M.O.; Landolin, J.M.; Yang, L.; Artieri, C.G.; van Baren, M.J.; Boley, N.; Booth, B.W. The Developmental Transcriptome of Drosophila melanogaster. Nature 2011, 471, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, S.J.; Baker, R.H.; Simon, S.; DeSalle, R. We Can’t All Be Supermodels: The Value of Comparative Transcriptomics to the Study of Non-model Insects. Insect Mol. Biol. 2015, 24, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP Genes and P450 Enzymes. Insect Mol. Biol. Biochem. 2012, 236–316. [Google Scholar] [CrossRef]
- Venkatesan, T.; Mahiba, S.H.; Jalali, S.K.; Ramya, S.L.; Prathibha, M. Detection of Insecticide Resistance and Mechanisms of Resistance in Field Populations of Chrysoperla zastrowi sillemi (Neuroptera: Chrysopidae) Collected from Different Geographical Locations in India. J. Biol. Control 2017, 31, 159–167. [Google Scholar] [CrossRef]
- Khan Pathan, A.; Sayyed, A.H.; Aslam, M.; Razaq, M.; Jilani, G.; Saleem, M.A. Evidence of Field-Evolved Resistance to Organophosphates and Pyrethroids in Chrysoperla carnea (Neuroptera: Chrysopidae). J. Econ. Entomol. 2008, 101, 1676–1684. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). In Cold Spring Harbor Protocols; CSHL Press: Cold Spring Harbor, NY, USA, 2010. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; ScienceOpen, Inc.: Boston, MA, USA, 2010. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a Full-Length Transcriptome without a Genome from RNA-Seq Data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinformatics 2011, 12, 1–16. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 Nomenclature, 2004. In Cytochrome P450 Protocols; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–10. [Google Scholar]
- Tijet, N.; Helvig, C.; Feyereisen, R. The Cytochrome P450 Gene Superfamily in Drosophila melanogaster: Annotation, Intron-Exon Organization and Phylogeny. Gene 2001, 262, 189–198. [Google Scholar] [CrossRef]
- Zhu, F.; Moural, T.W.; Shah, K.; Palli, S.R. Integrated Analysis of Cytochrome P450 Gene Superfamily in the Red flour beetle, Tribolium castaneum. BMC Genom. 2013, 14, 1–12. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bustin, S.A.; Beaulieu, J.-F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.B.; Olsvik, P.A.; Penning, L.C.; Toegel, S. MIQE Precis: Practical Implementation of Minimum Standard Guidelines for Fluorescence-Based Quantitative Real-Time PCR Experiments. BMC Mol. Biol. 2010, 11, 1–5. [Google Scholar] [CrossRef]
- Rewitz, K.F.; O’Connor, M.B.; Gilbert, L.I. Molecular Evolution of the Insect Halloween Family of Cytochrome P450s: Phylogeny, Gene Organization and Functional Conservation. Insect Biochem. Mol. Biol. 2007, 37, 741–753. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T.; Feyereisen, R. Diversity and Evolution of the P450 Family in Arthropods. Insect Biochem. Mol. Biol. 2020, 127, 103490. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Xiao, D.; He, Y.; Yao, J.; Zhu, G.; Zhu, K.Y. Insecticide-Mediated up-Regulation of Cytochrome P450 Genes in the Red flour beetle (Tribolium castaneum). Int. J. Mol. Sci. 2015, 16, 2078–2098. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Kang, Z.-W.; Liu, F.-H.; Pang, R.-P.; Tian, H.-G.; Liu, T.-X. Effect of Sublethal Doses of Imidacloprid on the Biological Performance of Aphid Endoparasitoid Aphidius gifuensis (Hymenoptera: Aphidiidae) and Influence on Its Related Gene Expression. Front. Physiol. 2018, 9, 1729. [Google Scholar] [CrossRef]
- Malaquias, J.B.; Ramalho, F.S.; Omoto, C.; Godoy, W.A.C.; Silveira, R.F. Imidacloprid Affects the Functional Response of Predator Podisus nigrispinus (Dallas) (Heteroptera: Pentatomidae) to Strains of Spodoptera frugiperda (JE Smith) on Bt Cotton. Ecotoxicology 2014, 23, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Krischik, V.A.; Martin, L.A. Effect of Soil Application of Imidacloprid on Survival of Adult Green Lacewing, Chrysoperla carnea (Neuroptera: Chrysopidae), Used for Biological Control in Greenhouse. Biol. Control. 2007, 42, 172–177. [Google Scholar] [CrossRef]
- Roush, R.T.; Daly, J.C. The Role of Population Genetics in Resistance Research and Management. In Pesticide Resistance in Arthropods; Springer: Berlin/Heidelberg, Germany, 1990; pp. 97–152. [Google Scholar]
- Ashwini, M.; Mohan, M.; Sivakumar, G.; Venkatesan, T. Enhanced Insecticide-Resistance Spectrum in Green Lacewing Predator, Chrysoperla zastrowi sillemi (Strain PTS-8) and Its Potential Role in the Management of Sucking Pests of Cotton. Curr. Sci. 2021, 120, 423–428. [Google Scholar] [CrossRef]
- Li, Z.-Q.; Zhang, S.; Ma, Y.; Luo, J.-Y.; Wang, C.-Y.; Lv, L.-M.; Dong, S.-L.; Cui, J.-J. First Transcriptome and Digital Gene Expression Analysis in Neuroptera with an Emphasis on Chemoreception Genes in Chrysopa pallens (Rambur). PLoS ONE 2013, 8, e67151. [Google Scholar] [CrossRef]
- Zhu, F.; Moural, T.W.; Nelson, D.R.; Palli, S.R. A Specialist Herbivore Pest Adaptation to Xenobiotics through Up-Regulation of Multiple Cytochrome P450s. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Antony, B.; Johny, J.; Abdelazim, M.M.; Jakše, J.; Al-Saleh, M.A.; Pain, A. Global Transcriptome Profiling and Functional Analysis Reveal That Tissue-Specific Constitutive Overexpression of Cytochrome P450s Confers Tolerance to Imidacloprid in Palm weevils in Date Palm Fields. BMC Genom. 2019, 20, 1–23. [Google Scholar] [CrossRef]
- Guittard, E.; Blais, C.; Maria, A.; Parvy, J.-P.; Pasricha, S.; Lumb, C.; Lafont, R.; Daborn, P.J.; Dauphin-Villemant, C. CYP18A1, a Key Enzyme of Drosophila Steroid Hormone Inactivation, Is Essential for Metamorphosis. Dev. Biol. 2011, 349, 35–45. [Google Scholar] [CrossRef]
- Warren, J.T.; Petryk, A.; Marqués, G.; Jarcho, M.; Parvy, J.-P.; Dauphin-Villemant, C.; O’Connor, M.B.; Gilbert, L.I. Molecular and Biochemical Characterization of Two P450 Enzymes in the Ecdysteroidogenic Pathway of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2002, 99, 11043–11048. [Google Scholar] [CrossRef] [PubMed]
- Sztal, T.; Chung, H.; Berger, S.; Currie, P.D.; Batterham, P.; Daborn, P.J. A Cytochrome P450 Conserved in Insects is Involved in Cuticle Formation. PLoS ONE 2012, 7, e36544. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, X.; Liu, Z.; Wang, F.; Fan, L.; Wu, C.; Yao, Y. Knockdown of CYP301B1 and CYP6AX1v2 Increases the Susceptibility of the Brown planthopper to Beta-Asarone, a Potential Plant-Derived Insecticide. Int. J. Biol. Macromol. 2021, 171, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Guzov, V.M.; Unnithan, G.C.; Chernogolov, A.A.; Feyereisen, R. CYP12A1, a Mitochondrial Cytochrome P450 from the House fly. Arch. Biochem. Biophys. 1998, 359, 231–240. [Google Scholar] [CrossRef]
- Brandt, A.; Scharf, M.; Pedra, J.H.F.; Holmes, G.; Dean, A.; Kreitman, M.; Pittendrigh, B.R. Differential Expression and Induction of Two Drosophila Cytochrome P450 Genes near the Rst (2) DDT Locus. Insect Mol. Biol. 2002, 11, 337–341. [Google Scholar] [CrossRef]
- Li, F.; Ma, K.; Liu, Y.; Zhou, J.-J.; Gao, X. Characterization of the Cytochrome P450 Gene CYP305A1 of the Cotton aphid (Hemiptera: Aphididae) and Its Responsive Cis-Elements to Plant Allelochemicals. J. Econ. Entomol. 2019, 112, 1365–1371. [Google Scholar] [CrossRef]
- Feyereisen, R. Arthropod CYPomes Illustrate the Tempo and Mode in P450 Evolution. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 19–28. [Google Scholar] [CrossRef]
- Feyereisen, R. Evolution of Insect P450. Biochem. Soc. Trans. 2006, 34, 1252–1255. [Google Scholar] [CrossRef]
- Puinean, A.M.; Foster, S.P.; Oliphant, L.; Denholm, I.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Bass, C. Amplification of a Cytochrome P450 Gene is Associated with Resistance to Neonicotinoid Insecticides in the Aphid Myzus persicae. PLoS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef]
- Wu, S.-F.; Zeng, B.; Zheng, C.; Mu, X.-C.; Zhang, Y.; Hu, J.; Zhang, S.; Gao, C.-F.; Shen, J.-L. The Evolution of Insecticide Resistance in the Brown planthopper (Nilaparvata lugens Stål) of China in the Period 2012–2016. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Joußen, N.; Heckel, D.G.; Haas, M.; Schuphan, I.; Schmidt, B. Metabolism of Imidacloprid and DDT by P450 CYP6G1 Expressed in Cell Cultures of Nicotiana tabacum Suggests Detoxification of These Insecticides in Cyp6g1-overexpressing Strains of Drosophila melanogaster, Leading to Resistance. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.; Boundy, S.; Yen, J.; Pittendrigh, B. DDT Resistance in Drosophila Correlates with Cyp6g1 Over-Expression and Confers Cross-Resistance to the Neonicotinoid Imidacloprid. Mol. Genet. Genomics 2001, 266, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-L.; Liu, S.-W.; Baerson, S.R.; Qin, Z.; Ma, Z.-H.; Su, Y.-J.; Zhang, J.-E. Identification and Functional Analysis of a Novel Cytochrome P450 Gene CYP9A105 Associated with Pyrethroid Detoxification in Spodoptera exigua Hübner. Int. J. Mol. Sci. 2018, 19, 737. [Google Scholar] [CrossRef] [PubMed]
- David, J.P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J.I. Role of Cytochrome P450s in Insecticide Resistance: Impact on the Control of Mosquito-Borne Diseases and Use of Insecticides on Earth. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef] [PubMed]
- Dulbecco, A.B.; Moriconi, D.E.; Calderón-Fernández, G.M.; Lynn, S.; McCarthy, A.; Roca-Acevedo, G.; Salamanca-Moreno, J.A.; Juárez, M.P.; Pedrini, N. Integument CYP Genes of the Largest Genome-Wide Cytochrome P450 Expansions in Triatomines Participate in Detoxification in Deltamethrin-Resistant Triatoma infestans. Sci. Reports 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Feyereisen, R. Origin and Evolution of the CYP4G Subfamily in Insects, Cytochrome P450 Enzymes Involved in Cuticular Hydrocarbon Synthesis. Mol. Phylogenet. Evol. 2020, 143, 106695. [Google Scholar] [CrossRef]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized Hepatocyte-like Cells Regulate Drosophila Lipid Metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef]
- Maıbeche-Coisne, M.; Monti-Dedieu, L.; Aragon, S.; Dauphin-Villemant, C. A New Cytochrome P450 from Drosophila melanogaster, CYP4G15, Expressed in the Nervous System. Biochem. Biophys. Res. Commun. 2000, 273, 1132–1137. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Population | LC50 (mL/Liter) | Slope ± SE | Fiducial Limits | LC90 (mL/Liter) | Fiducial Limits | X2 | Df | RR | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | ||||||||
| 1. | Coimbatore | 0.33 | 1.08 ± 0.28 | 0.116 | 0.618 | 5.99 | 2.197 | 131.74 | 8.8 | 7 | 3.60 |
| 2. | Ludhiana | 0.17 | 0.98 ± 0.30 | 0.027 | 0.347 | 3.47 | 1.384 | 80.072 | 7.52 | 7 | 1.90 |
| 3. | Sirsa | 0.45 | 1.18 ± 0.29 | 0.232 | 0.815 | 5.53 | 2.310 | 51.35 | 7.34 | 7 | 5.0 |
| 4. | Delhi | 0.47 | 1.45 ± 0.31 | 0.275 | 0.758 | 3.53 | 1.766 | 16.399 | 0.225 | 7 | 5.20 |
| 5. | Darwad | 0.41 | 1.39 ± 0.31 | 0.230 | 0.678 | 3.46 | 1.690 | 17.823 | 6.93 | 7 | 4.50 |
| 6. | Guntur | 0.36 | 1.29 ± 0.31 | 0.180 | 0.603 | 3.53 | 1.655 | 22.23 | 12.49 | 7 | 4.0 |
| 7. | Sriganganagar | 1.08 | 1.67 ± 0.32 | 0.709 | 1.852 | 6.26 | 3.154 | 24.797 | 15.29 | 7 | 12.0 |
| 8. | Anand | 0.33 | 1.24± 0.31 | 0.156 | 0.567 | 3.55 | 1.633 | 25.12 | 9.56 | 7 | 3.60 |
| 9. | Susceptible | 0.09 | 1.09 ± 0.35 | 0.004 | 0.197 | 1.34 | 0.643 | 16.510 | 6.54 | 7 | |
| Sl. No. | Strain | LC50 (mL/Liter) | Slope ± SE | Fiducial Limits | LC90 (mL/Liter) | Fiducial Limits | X2 | Df | RR | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | ||||||||
| 1. | Resistant (Sriganganagar) | 2.24 | 1.10± 0.15 | 1.473 | 3.921 | 32.58 | 14.101 | 132.713 | 8.8 | 7 | 56 |
| 2. | Susceptible | 0.04 | 1.0 ± 0.15 | 0.016 | 0.065 | 0.713 | 0.419 | 1.622 | 1.62 | 7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pathak, J.; Ramasamy, G.G.; Agrawal, A.; Srivastava, S.; Basavaarya, B.R.; Muthugounder, M.; Muniyappa, V.K.; Maria, P.; Rai, A.; Venkatesan, T. Comparative Transcriptome Analysis to Reveal Differentially Expressed Cytochrome P450 in Response to Imidacloprid in the Aphid Lion, Chrysoperla zastrowi sillemi (Esben-Petersen). Insects 2022, 13, 900. https://doi.org/10.3390/insects13100900
Pathak J, Ramasamy GG, Agrawal A, Srivastava S, Basavaarya BR, Muthugounder M, Muniyappa VK, Maria P, Rai A, Venkatesan T. Comparative Transcriptome Analysis to Reveal Differentially Expressed Cytochrome P450 in Response to Imidacloprid in the Aphid Lion, Chrysoperla zastrowi sillemi (Esben-Petersen). Insects. 2022; 13(10):900. https://doi.org/10.3390/insects13100900
Chicago/Turabian StylePathak, Jyoti, Gandhi Gracy Ramasamy, Aditi Agrawal, Subhi Srivastava, Bhusangar Raghavendra Basavaarya, Mohan Muthugounder, Venugopal Kundalagurki Muniyappa, Pratheepa Maria, Anil Rai, and Thiruvengadam Venkatesan. 2022. "Comparative Transcriptome Analysis to Reveal Differentially Expressed Cytochrome P450 in Response to Imidacloprid in the Aphid Lion, Chrysoperla zastrowi sillemi (Esben-Petersen)" Insects 13, no. 10: 900. https://doi.org/10.3390/insects13100900
APA StylePathak, J., Ramasamy, G. G., Agrawal, A., Srivastava, S., Basavaarya, B. R., Muthugounder, M., Muniyappa, V. K., Maria, P., Rai, A., & Venkatesan, T. (2022). Comparative Transcriptome Analysis to Reveal Differentially Expressed Cytochrome P450 in Response to Imidacloprid in the Aphid Lion, Chrysoperla zastrowi sillemi (Esben-Petersen). Insects, 13(10), 900. https://doi.org/10.3390/insects13100900