
Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
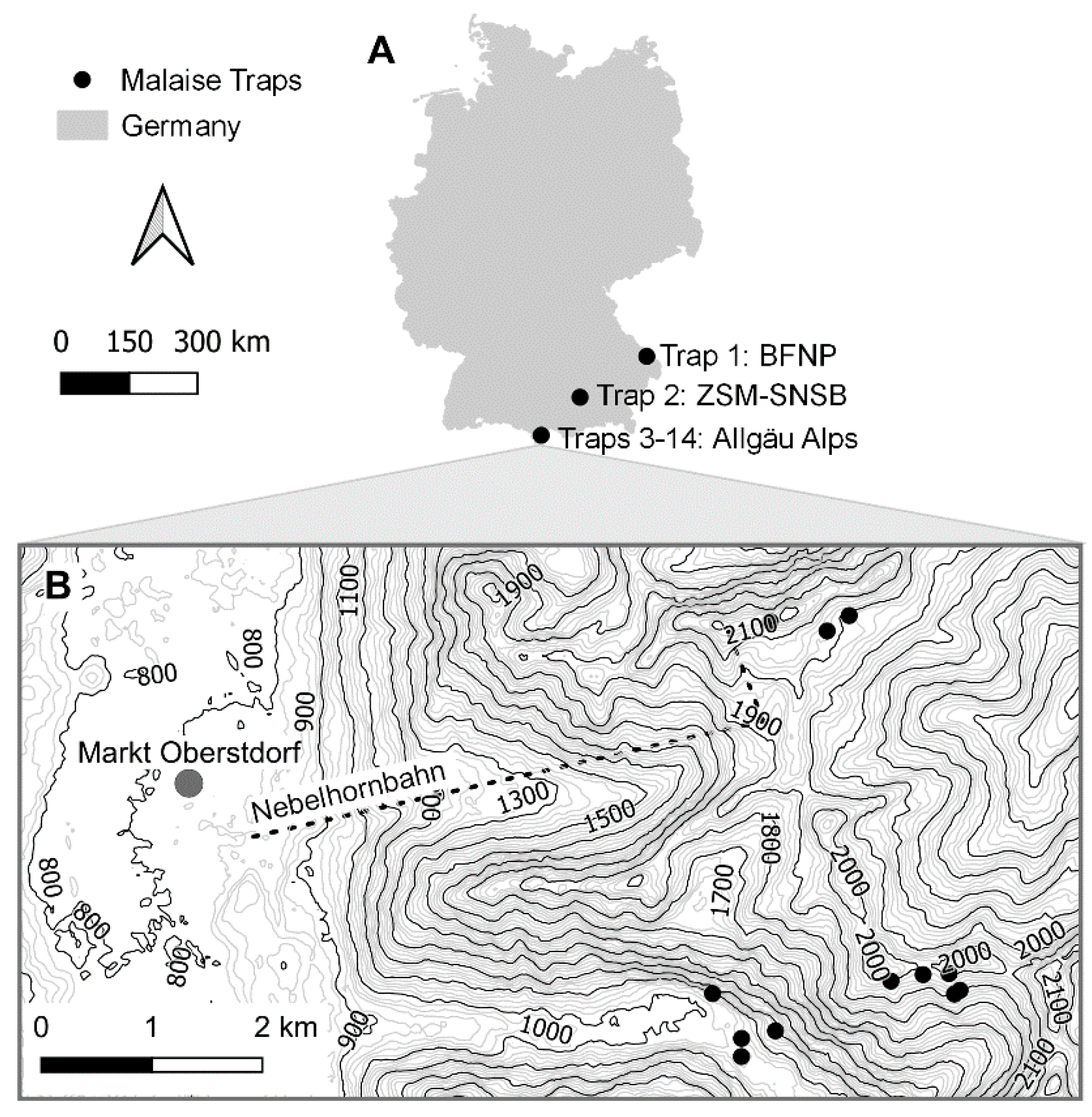
2.1. Malaise Tap Sites
2.2. Processing of Specimens
2.3. Data Analysis
2.4. Extrapolating Species Numbers
- We calculated a Recovery Ratio by dividing the number of BINs detected through sequencing by the species count for each of the 15 families and for all Diptera (BIN/species ratio). This approach generated a ratio for each well-known family, for each DT family, and for all Diptera.
- We estimated the maximum number of species for each “dark taxon” for both Germany and Bavaria by dividing its BIN count by the average BIN/species ratio of all 11 well-known families.
- We estimated the minimum species number for each “dark taxon” by dividing all Diptera BINs by all Diptera species (i.e., 9544). Because this calculation includes numerous families with cryptic diversity, the resultant values underestimate the diversity of the DT families.
3. Results
3.1. Sequencing Results
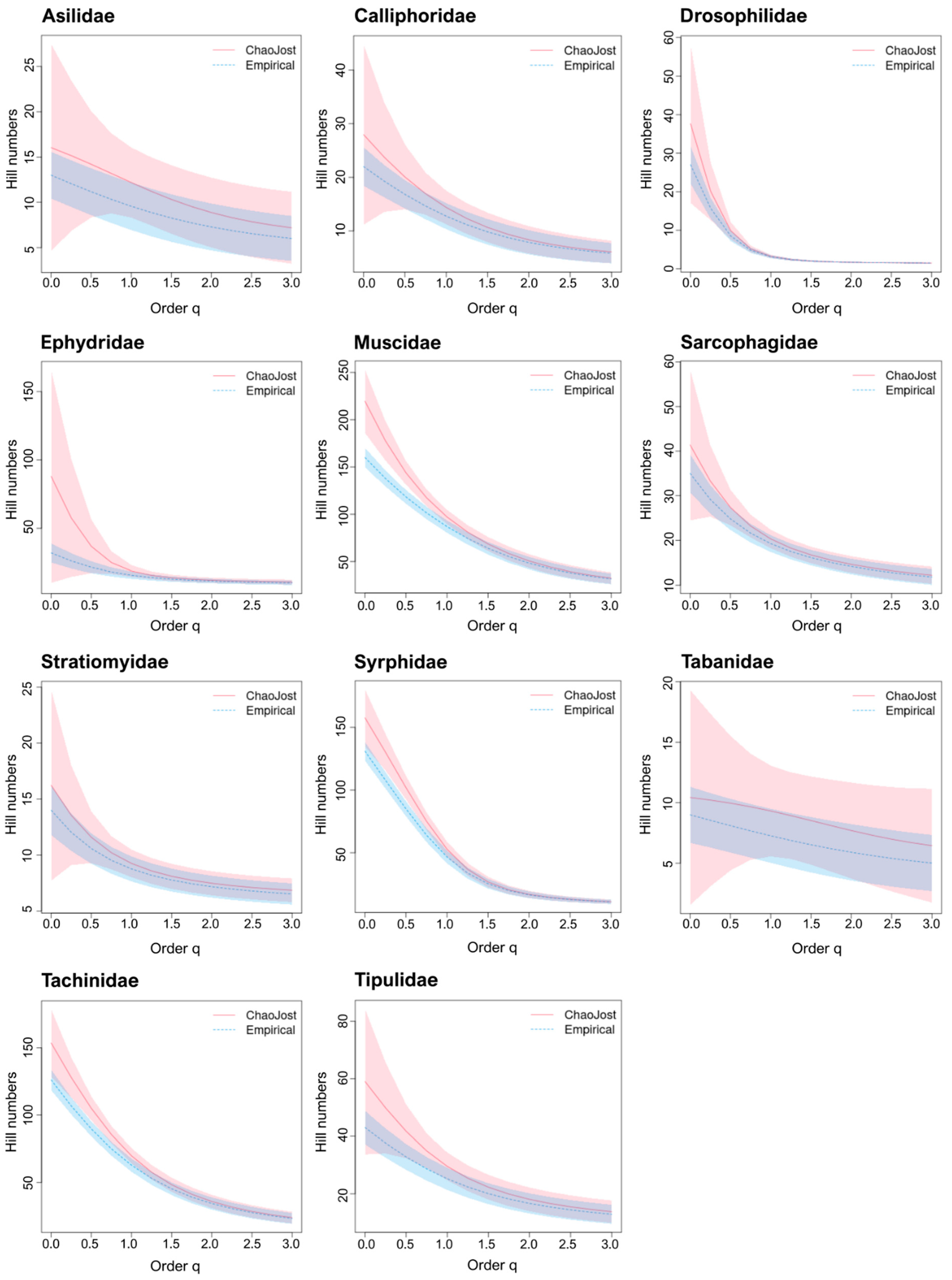
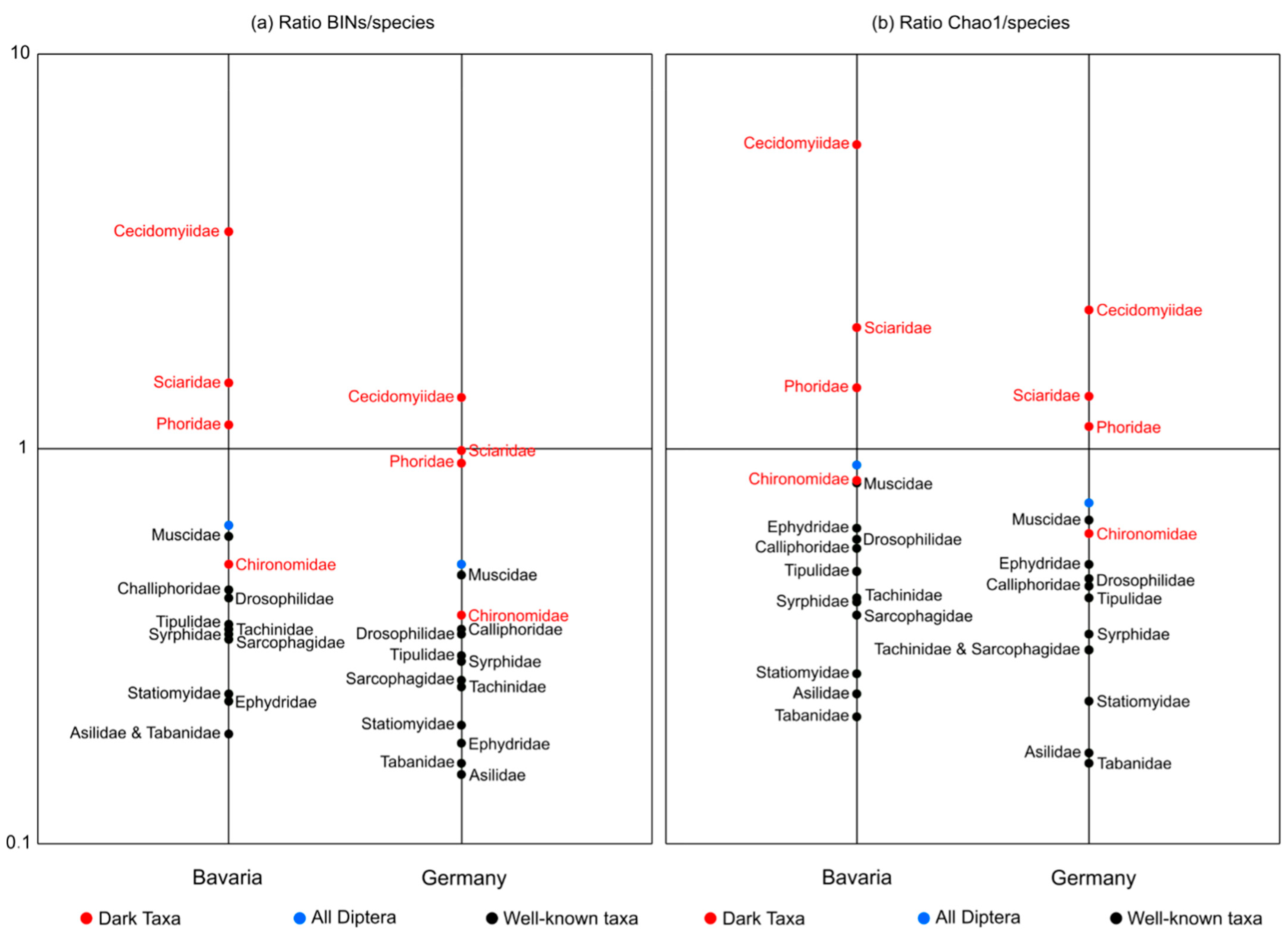
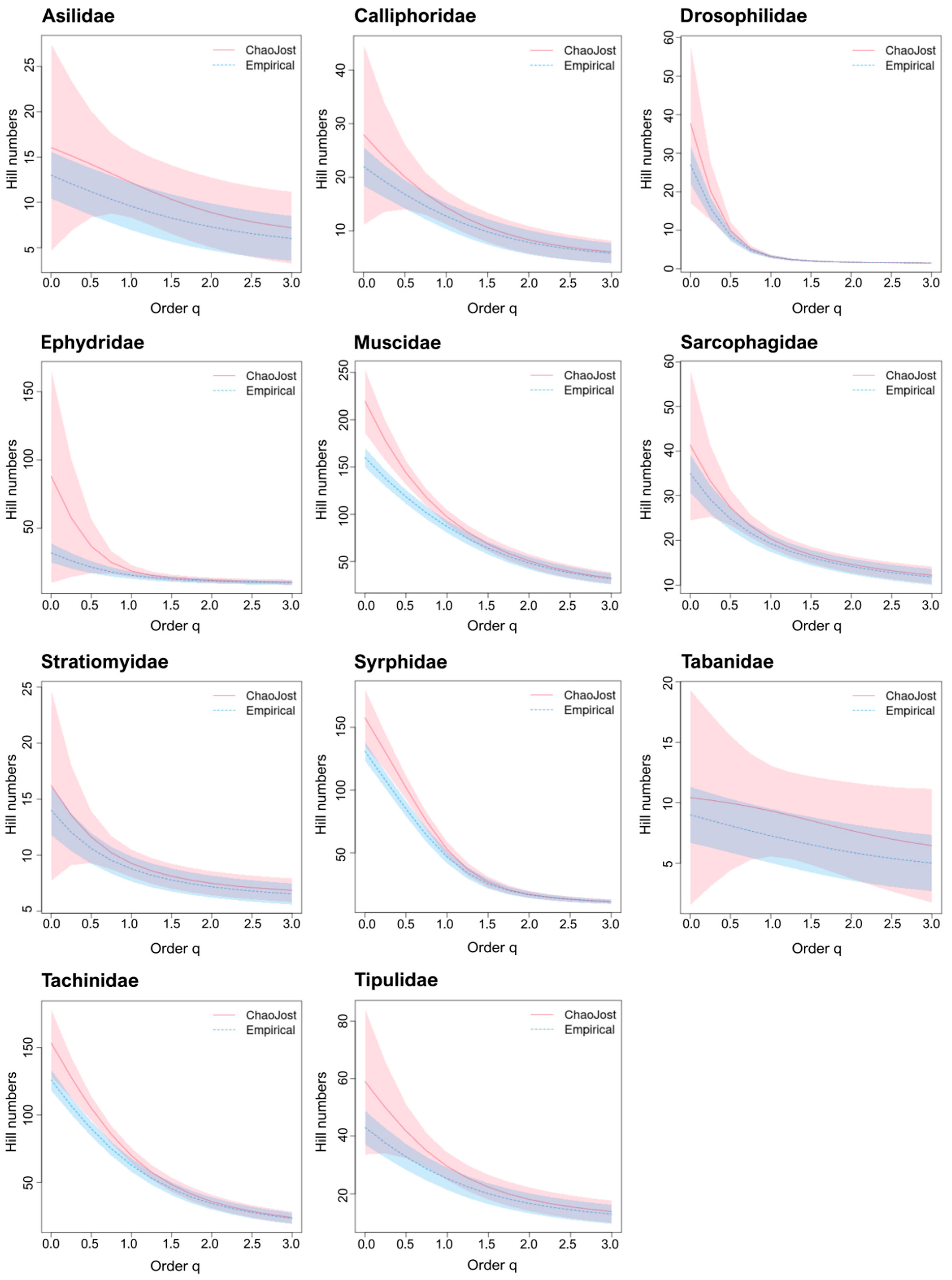
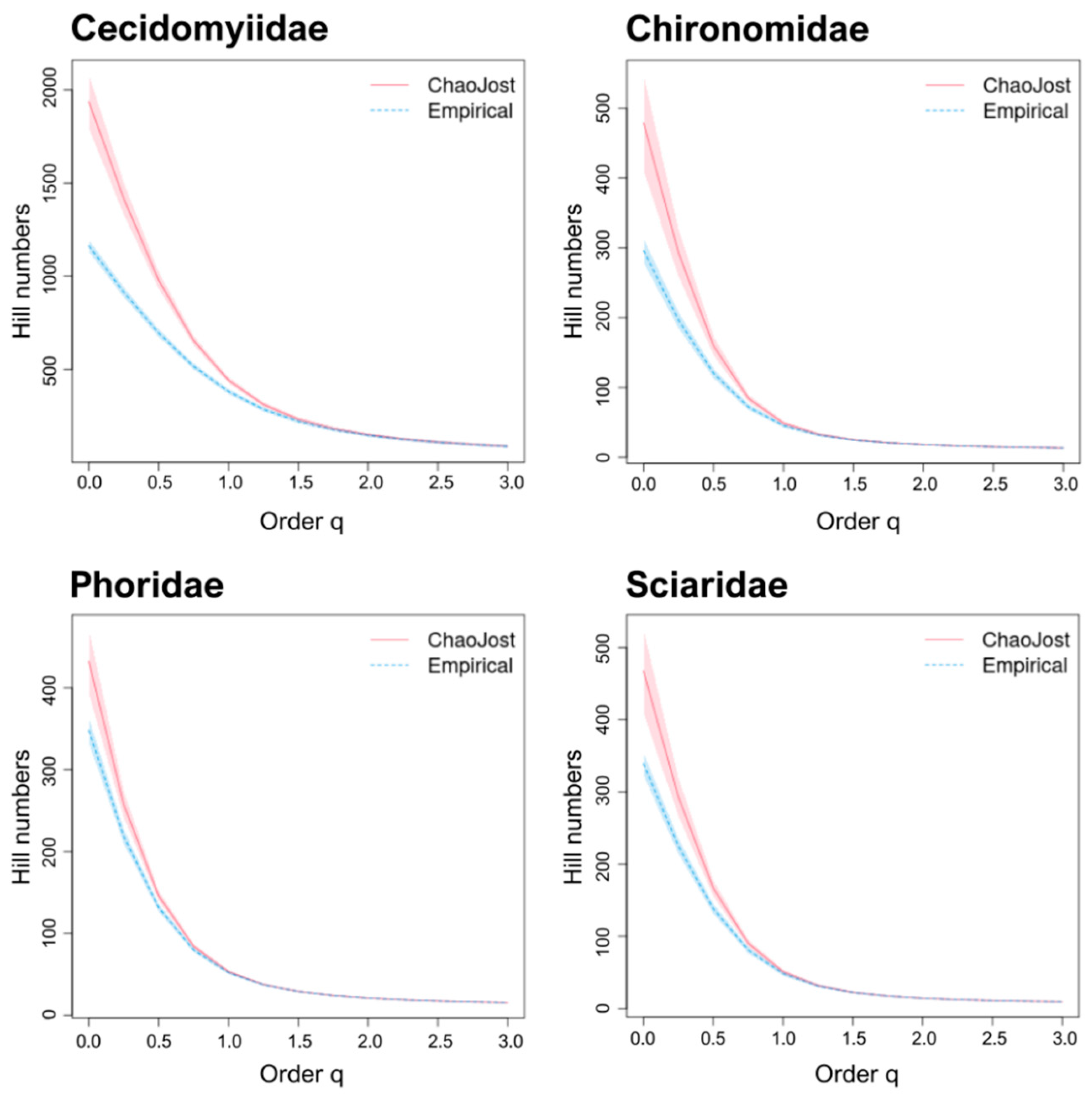
3.2. Estimation of Taxon Diversity Using BIN/Species Ratios
3.3. Estimation of Taxon Diversity Using Chao1/Species Ratios
3.4. Extrapolating Species Numbers
4. Discussion
4.1. DNA Barcoding: Using BIN Numbers as Proxies for Species Numbers
- Limited geographic sampling as our data originates from few sites in Bavaria only, covering a tiny fraction of habitat types otherwise present.
- Limited funding constrained analysis to just 5% of the 1.2 million specimens that were collected.
4.2. BIN & Chao1/Species Ratios: Well-Known Families versus DT Families
4.3. Discrepancies in Taxa Coverage in Our Malaise Traps
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Trap | Processed Collection Events |
|---|---|---|
| BFNP 2012 | 1 | 8 May; 22 May; 8 June; 20 June; 4 July; 25 July; 12 August; 3 September; 22 September 2012. |
| ZSM-SNSB | 2 | 10 April; 8 May; 5 June; 3 July; 31 July; 28 August; 25 September; 23 October; 20 November; 29 December 2017. |
| Allgäu Alps: Oytal | 3 | 4 May; 17 May; 1 June; 16 June; 5 July: 20 July; 7 August; 29 August; 2 October; 27 October 2014. |
| Allgäu Alps: Oytal | 4 | 4 May; 17 May; 1 June; 16 June; 5 July: 20 July; 7 August; 29 August; 2 October; 27 October 2014. |
| Allgäu Alps: Oytal | 5 | 4 May; 17 May; 1 June; 16 June; 5 July: 20 July; 7 August; 29 August; 2 October; 27 October 2014. |
| Allgäu Alps: Oytal | 6 | 4 May; 17 May; 1 June; 16 June; 5 July: 20 July; 7 August; 29 August; 2 October; 27 October 2014. |
| Allgäu Alps: Schochen | 7 | 21 June; 4 July; 17 July; 6 August; 4 September; 29 September; 19 October 2014. |
| Allgäu Alps: Schochen | 8 | 21 June; 4 July; 17 July; 6 August; 4 September; 29 September; 19 October 2014. |
| Allgäu Alps: Schochen | 9 | 21 June; 4 July; 17 July; 6 August; 4 September; 29 September; 19 October 2014. |
| Allgäu Alps: Schochen | 10 | 21 June; 4 July; 17 July; 6 August; 4 September; 29 September; 19 October 2014. |
| Allgäu Alps: Schochen | 11 | 21 June; 4 July; 17 July; 6 August; 4 September; 29 September; 19 October 2014. |
| Allgäu Alps: Koblat | 12 | 23 June, 4 July, 17 July; 8 August; 8 September; 5 September, 27 September; 20 October 2014. |
| Allgäu Alps: Koblat | 13 | 23 June, 4 July, 17 July; 8 August; 8 September; 5 September, 27 September; 20 October 2014. |
| Allgäu Alps: Koblat | 14 | 23 June, 4 July, 17 July; 8 August; 8 September; 5 September, 27 September; 20 October 2014. |
References
- Hausmann, A.; Krogmann, L.; Peters, R.; Rduch, V.; Schmidt, S. GBOL III: DARK TAXA. iBOL Barcode Bull. 2020, 10, 4. [Google Scholar] [CrossRef]
- Dathe, H.H.; Taeger, A.; Blank, S.M. Entomofauna Germanica Band 4 (Hymenoptera). Verzeichnis der Hautflügler Deutschlands. Entomol. Nachr. Ber. 2001, Beiheft 7, 1–180. [Google Scholar]
- Dathe, H.H.; Blank, S.M. Nachträge zum Verzeichnis der Hautflügler Deutschlands, Entomofauna Germanica Band 4 (Hymenoptera) (1). Entomol. Nachr. Ber. 2004, 48, 179–183. [Google Scholar]
- Völkl, W.; Blick, T.; Kornacher, P.M.; Martens, H. Quantitativer Überblick über die rezente Fauna von Deutschland. Nat. Landsch. 2004, 79, 293–295. [Google Scholar]
- Schumann, H. Checkliste der Dipteren Deutschlands. In Entomofauna Germanica 2. Studia Dipterologica Supplement; Ampyx-Verlag: Halle/Saale, Germany, 1999; Volume 2, p. 354. [Google Scholar]
- Schumann, H. Erster Nachtrag zur “Checkliste der Dipteren Deutschlands”. Studia Dipterol. 2002, 9, 437–445. [Google Scholar]
- Schumann, H. Zweiter Nachtrag zur “Checkliste der Dipteren Deutschlands”. Studia Dipterol. 2004, 11, 619–630. [Google Scholar]
- Schumann, H. Dritter Nachtrag zur “Checkliste der Dipteren Deutschlands”. Studia Dipterol. 2010, 16, 17–27. [Google Scholar]
- de Carvalho, M.R. Revisiting the taxonomic impediment. Science 2005, 307, 353b. [Google Scholar] [CrossRef]
- Mandelik, Y.; Roll, U.; Fleischer, A. Cost-efficiency of biodiversity indicators for Mediterranean ecosystems and the effects of socio-economic factors. J. Appl. Ecol. 2010, 47, 1179–1188. [Google Scholar] [CrossRef]
- Chan, A.; Chiang, L.-P.; Hapuarachchi, H.C.; Tan, C.-H.; Pang, S.-C.; Lee, R.; Lee, K.-S.; Ng, L.-C.; Lam-Phua, S.-G. DNA Barcoding: Complementing morphological identification of mosquito species in Singapore. Parasites Vectors 2014, 7, 569. [Google Scholar] [CrossRef]
- Porter, T.M.; Gibson, J.F.; Shokralla, S.; Baird, D.J.; Golding, G.B.; Hajibabaei, M. Rapid and accurate taxonomic classification of insect (class Insecta) cytochrome c oxidase subunit 1 (COI) DNA barcode sequences using a naïve Bayesian classifier. Mol. Ecol. Resour. 2014, 14, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Morinière, J.; Balke, M.; Doczkal, D.; Geiger, M.F.; Hardulak, L.A.; Haszprunar, G.; Hausmann, A.; Hendrich, L.; Regalado, L.; Rulik, B.; et al. A DNA barcode library for 5,200 German flies and midges (Insecta: Diptera) and its implications for metabarcoding-based biomonitoring. Mol. Ecol. Resour. 2019, 19, 900–928. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.V. Sampling methods for adult flies (Diptera). In Measuring Arthropod Biodiversity: A Handbook of Sampling Methods; Santos, J.C., Fernandes, G.W., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 187–204. [Google Scholar] [CrossRef]
- Giangrande, A. Biodiversity, conservation, and the ‘taxonomic impediment’. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 451–459. [Google Scholar] [CrossRef]
- de Carvalho, M.R.; Bockmann, F.A.; Amorim, D.S.; Brandão, C.R.F.; de Vivo, M.; de Figueiredo, J.L.; Britski, H.A.; de Pinna, M.C.C.; Menezes, N.A.; Marques, F.P.L.; et al. Taxonomic impediment or impediment to taxonomy? A commentary on systematics and the cybertaxonomic-automation paradigm. Evol. Biol. 2007, 34, 140–143. [Google Scholar] [CrossRef]
- Wenzel, M.; Schmitt, T.; Weitzel, M.; Seitz, A. The severe decline of butterflies on western German calcareous grasslands during the last 30 years: A conservation problem. Biol. Conserv. 2006, 4, 542–552. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Brook, B.W.; Bradshaw, C.J.A. Causes and consequences of species extinctions. In The Princeton Guide to Ecology; Levin, S.A., Ed.; Princton University Press: Princeton, NJ, USA, 2009; pp. 514–520. [Google Scholar]
- Coleman, C.O. Taxonomy in times of the taxonomic impediment—Examples from the community of experts on amphipod crustaceans. J. Crustacean Biol. 2015, 35, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D. The insect apocalypse, and why it matters. Curr. Biol. 2019, 29, R967–R971. [Google Scholar] [CrossRef]
- Page, R.D.M. DNA barcoding and taxonomy: Dark taxa and dark texts. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150334. [Google Scholar] [CrossRef] [Green Version]
- GBOL III: Dark Taxa—Dark Taxa. Available online: https://bolgermany.de/home/gbol3/ (accessed on 16 December 2021).
- German Barcode of Life. Available online: https://bolgermany.de (accessed on 8 October 2021).
- Hausmann, A.; Haszprunar, G.; Hebert, P.D.N. DNA barcoding the geometrid fauna of Bavaria (Lepidoptera): Successes, surprises, and questions. PLoS ONE 2011, 6, e17134. [Google Scholar] [CrossRef]
- Hendrich, L.; Morinière, J.; Haszprunar, G.; Hebert, P.D.N.; Hausmann, A.; Köhler, F.; Balke, M. A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3500 identified species to BOLD. Mol. Ecol. Resour. 2015, 15, 795–818. [Google Scholar] [CrossRef] [PubMed]
- Kotrba, M. The DNA barcoding project on German Diptera: An appreciative and critical analysis with four suggestions for improving the development and reliability of DNA-based identification. Eur. J. Entomol. 2020, 117, 315–327. [Google Scholar] [CrossRef]
- Schacht, W. Katalog der Zweiflügler (Mücken und Fliegen) Bayerns Familienliste—Artenliste—Literaturverzeichnis—Bestimmungsliteratur (Insecta: Diptera). Available online: http://www.zsm.mwn.de/docs_zsm/htdocs/dip/DiptBayKat.pdf (accessed on 8 October 2021).
- Wolff, D.; Gebel, M.; Geller-Grimm, F. Die Raubfliegen Deutschlands; Quelle & Meyer: Wiebelsheim, Germany, 2018. [Google Scholar]
- Jentzsch, M. Erster Entwurf einer Bibliographie zur Waffenfliegen-Fauna Deutschlands (Diptera: Stratiomyidae). Studia Dipterol. 2015, 22, 187–197. [Google Scholar]
- Doczkal, D.; Claußen, C.; Ssymank, A. Erster Nachtrag und Korrekturen zur Checkliste der Schwebfliegen Deutschlands (Diptera, Syrphidae). Volucella 2002, 6, 167–173. [Google Scholar]
- Heiss, R.; Fahldieck, M.; Stuke, J.-H. Kommentierte Checkliste der Schnaken der Bundesländer Niedersachsen und Bremen (Diptera, Tipulidae). Entomol. Nachr. Ber. 2019, 63, 245–259. [Google Scholar]
- Heiss, R.; Merkel-Wallner, G. Ein Beitrag zur Schnaken-Fauna Bayerns—Neue und wenig bekannte Arten aus Malaisefallen-Fängen 2007 bis 2009. Beiträge Bayer. Entomofaunist. 2013, 12, 17–30. [Google Scholar]
- Menzel, F. Die Trauermücken—Fauna der Bundesrepublik Deutschland (Diptera: Sciaridae). Beiträge Entomol. 2000, 50, 317–355. [Google Scholar] [CrossRef]
- Drees, M. Die Fleischfliegen des Hagener Raumes (Diptera: Sarcophagidae). Entomol. Z. 2014, 124, 3. [Google Scholar]
- Stuke, J.-H. Eine Kritische Liste der aus Deutschland nachgewiesenen Ephydridae mit der Beschreibung einer neuen Art (Diptera). Entomol. Z. 2011, 121, 115–126. [Google Scholar]
- Stuke, J.-H. Die Fruchtfliegen Niedersachsens und Bremens (Diptera, Drosophilidae). Linz. Biol. Beiträge 2020, 52, 475–499. [Google Scholar]
- Skuhravá, M.; Skuhravý, V.; Meyer, H. Gall midges (Diptera: Cecidomyiidae: Cecidomyiinae) of Germany—Faunistics, ecology and zoogeography. Faun.-Ökol. Mitt. 2014, Suppl. 38, 1–200. [Google Scholar]
- Global Malaise Program. Available online: https://biodiversitygenomics.net/projects/gmp/ (accessed on 8 October 2021).
- Chan-Canché, R.; Ballina-Gómez, H.; Leirana-Alcocer, J.; Bordera, S.; González-Moreno, A. Sampling of parasitoid Hymenoptera: Influence of the height on the ground. J. Hymenopt. Res. 2020, 78, 19–31. [Google Scholar] [CrossRef]
- Resources of CCDB. Available online: https://ccdb.ca/resources/ (accessed on 8 October 2021).
- Hebert, P.D.N.; Braukmann, T.W.A.; Prosser, S.W.J.; Ratnasingham, S.; deWaard, J.R.; Ivanova, N.V.; Janzen, D.H.; Hallwachs, W.; Naik, S.; Sones, J.E.; et al. A sequel to Sanger: Amplicon sequencing that scales. BMC Genom. 2018, 19, 219. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chiu, C.-H. Estimation of species richness and shared species richness. In Methods and Applications of Statistics in the Atmospheric and Earth Sciences; Balakrishnan, N., Ed.; Wiley: New York, NY, USA, 2012; pp. 76–111. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Ma, K.H.; Chiu, T.C.H. Spade R: Species-richness prediction and diversity estimation with R. Available online: https://cran.r-project.org/web/packages/SpadeR/SpadeR.pdf (accessed on 16 December 2021).
- Haszprunar, G. Barcoding Fauna Bavarica—Eine Chance für die Entomologie. Nachr. Bayer. Entomol. 2009, 58, 4. [Google Scholar]
- Büro für Freilandökologie Dr. Jürgen Esser. Available online: https://freilandoekologie-esser.de/ (accessed on 16 December 2021).
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; deWaard, J.R. Counting animal species with DNA barcodes: Canadian insects. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150333. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. Available online: http://www.barcodinglife.org (accessed on 16 December 2021). [CrossRef] [Green Version]
- Hebert, P.D.N.; deWaard, J.R.; Landry, J.-F. DNA barcodes for 1/1000 of the animal kingdom. Biol. Lett. 2010, 6, 359–362. [Google Scholar] [CrossRef]
- Reid, B.N.; Le, M.; McCord, W.P.; Iverson, J.B.; Georges, A.; Bergmann, T.; Amato, G.; Desalle, R.; Naro-Maciel, E. Comparing and combining distance-based and character-based approaches for barcoding turtles. Mol. Ecol. Resour. 2011, 11, 956–967. [Google Scholar] [CrossRef] [Green Version]
- Chimeno, C.; Morinière, J.; Podhorna, J.; Hardulak, L.; Hausmann, A.; Reckel, F.; Grunwald, J.E.; Penning, R.; Haszprunar, G. DNA barcoding in forensic entomology—Establishing a DNA reference library of potentially forensic relevant arthropod species. J. Forensic Sci. 2019, 64, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Astrin, J.J.; Hannig, K.; Peters, M.K.; Stoeckle, M.Y.; Wägele, J.-W. Molecular species identification of Central European ground beetles (Coleoptera: Carabidae) using nuclear rDNA expansion segments and DNA barcodes. Front. Zool. 2010, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamilo, P.; Viljakainen, L.; Vihavainen, A. Exceptionally high density of NUMTs in the honeybee genome. Mol. Biol. Evol. 2007, 24, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many Species in One: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [Green Version]
- Buhay, J.E. “COI-like” sequences are becoming problematic in molecular systematic and DNA barcoding studies. J. Crustacean Biol. 2009, 29, 96–110. [Google Scholar] [CrossRef]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W. Molecular poltergeists: Mitochondrial DNA copies (Numts) in sequenced nuclear genomes. PLoS Genet. 2010, 6, e1000834. [Google Scholar] [CrossRef]
- Nesi, N.; Nakouné, E.; Cruaud, C.; Hassanin, A. DNA barcoding of African fruit bats (Mammalia, Pteropodidae). the mitochondrial genome does not provide a reliable discrimination between Epomophorus gambianus and Micropteropus pusillus. Comptes Rendus Biol. 2011, 334, 544–554. [Google Scholar] [CrossRef]
- Ermakov, O.A.; Simonov, E.; Surin, V.L.; Titov, S.V.; Brandler, O.V.; Ivanova, N.V.; Borisenko, A.V. Implications of hybridization, Numts, and overlooked diversity for DNA barcoding of Eurasian ground squirrels. PLoS ONE 2015, 10, e0117201. [Google Scholar] [CrossRef]
- Dufresnes, C.; Berroneau, M.; Dubey, S.; Litvinchuk, S.N.; Perrin, N. The effect of phylogeographic history on species boundaries: A comparative framework in Hyla tree frogs. Sci. Rep. 2020, 10, 5502. [Google Scholar] [CrossRef] [Green Version]
- Kmiec, B.; Woloszynska, M.; Janska, H. Heteroplasmy as a Common State of Mitochondrial Genetic Information in Plants and Animals. Curr. Genet. 2006, 50, 149–159. [Google Scholar] [CrossRef]
- Dobson, S.L. Evolution of Wolbachia cytoplasmic incompatibility. Evolution 2007, 58, 2156–2166. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. the diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, A.; Godfray, H.C.J.; Huemer, P.; Mutanen, M.; Rougerie, R.; van Nieukerken, E.J.; Ratnasingham, S.; Hebert, P.D.N. Genetic patterns in European geometrid moths revealed by the Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e84518. [Google Scholar] [CrossRef]
- The Chironomid Home Page. Available online: www.chironomidae.net (accessed on 16 December 2021).
- Karlsson, D.; Forshage, M.; Holston, K.; Ronquist, F. The data of the Swedish Malaise Trap Project, a countrywide inventory of Sweden’s insect fauna. Biodivers. Data J. 2020, 8, e56286. [Google Scholar] [CrossRef] [PubMed]
- Moeed, A.; Meads, M.J. Seasonality of arthropods caught in a malaise trap in mixed lowland forest of the Orongorongo Valley, New Zealand. N. Z. J. Zool. 1987, 14, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Matthews, R.; Matthews, J. The Malaise trap: Its utility and potential for sampling insect populations. Great Lakes Entomol. 2017, 4, 4. [Google Scholar]

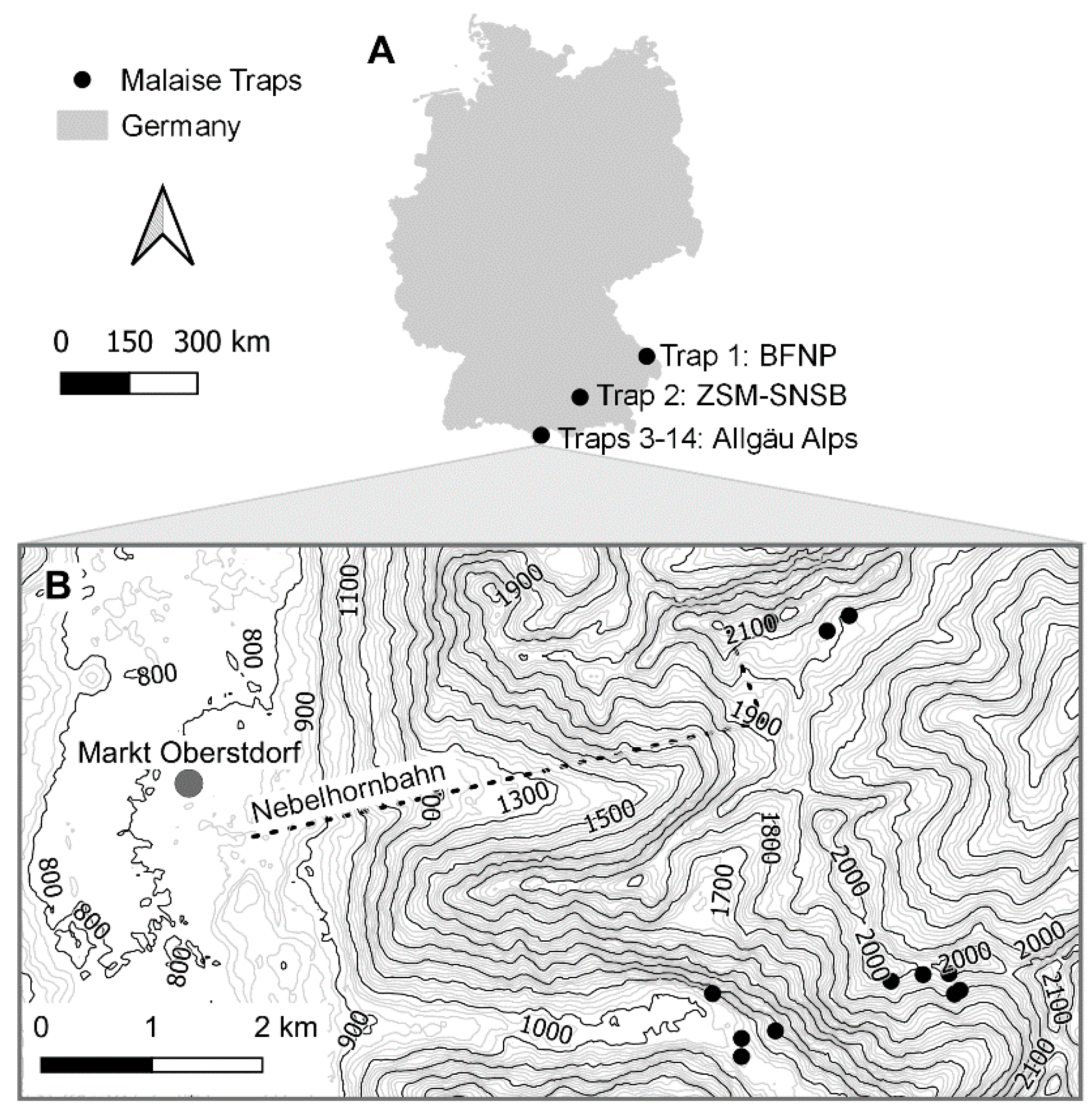
| Site | Trap | Coordinates | Elevation | Habitat |
|---|---|---|---|---|
| BFNP | Trap 1 | 48.9509° N 13.422° E | 842 m | Natural forest |
| ZSM | Trap 2 | 48.1648° N 11.4849° E | 519 m | Urban, pre-alpine meadow |
| Allgäu Alps: Oytal | Trap 3 | 47.39205° N 10.34093° E | 1122 m | Lake rock face |
| Allgäu Alps: Oytal | Trap 4 | 47.38903° N 10.34846° E | 1200 m | Cone of scree |
| Allgäu Alps: Oytal | Trap 5 | 47.38842° N 10.34440° E | 1056 m | Rough pasture |
| Allgäu Alps: Oytal | Trap 6 | 47.38695° N 10.34438° E | 1036 m | River |
| Allgäu Alps: Schochen | Trap 7 | 47.39202° N 10.36991° E | 1930 m | Alpine grassland |
| Allgäu Alps: Schochen | Trap 8 | 47.39232° N 10.37057° E | 1908 m | Spring |
| Allgäu Alps: Schochen | Trap 9 | 47.39368° N 10.36926° E | 2032 m | South-exposed ridge with Blaugras-Horstseggenrasen |
| Allgäu Alps: Schochen | Trap 10 | 47.39307° N 10.36229° E | 2010 m | South-exposed rock |
| Allgäu Alps: Schochen | Trap 11 | 47.39360° N 10.36615° E | 1980 m | Snow bed |
| Allgäu Alps: Koblat | Trap 12 | 47.42223° N 10.34783° E | 2160 m | South-exposed rock face |
| Allgäu Alps: Koblat | Trap 13 | 47.42147° N 10.35465° E | 2033 m | Snow bed |
| Allgäu Alps: Koblat | Trap 14 | 47.42272° N 10.35730° E | 2005 m | Mountain pine bush |
| Taxon | Bavarian Species Count | German Species Count | Species Count Bavaria/Germany |
|---|---|---|---|
| Asilidae | 68 [28] | 85 [29] | 0.80 |
| Calliphoridae | 50 * | 62 [35] | 0.80 * |
| Drosophilidae | 64 [28] | 81 [37] | 0.79 |
| Ephydridae | 140 * | 174 [38] | 0.80 * |
| Muscidae | 267 * | 334 [48] | 0.80 * |
| Sarcophagidae | 107 * | 134 [35] | 0.80 * |
| Stratiomyidae | 59 [28] | 71 [30,48] | 0.83 |
| Syrphidae | 389 [28] | 458 [31] | 0.85 |
| Tabanidae | 47 [28] | 58 [8,48] | 0.81 |
| Tachinidae | 361 [28] | 501 [48] | 0.72 |
| Tipulidae | 120 [33] | 142 [32] | 0.85 |
| Cecidomyiidae | 328 [38] | 859 [5,6,7,8] | 0.38 |
| Chironomidae | 576 [28] | 781 [5,6,7,8] | 0.74 |
| Phoridae | 302 * | 378 [5,6,7,8] | 0.80 * |
| Sciaridae | 231 [28] | 343 [43] | 0.67 |
| All Diptera | 7635 * | 9544 [8] | 0.80 * |
| BFNP | ZSM | Allgäu Alps | Total | |
|---|---|---|---|---|
| Samples (trap × collection events) | 1 × 9 = 9 | 1 × 10 = 10 | 8 × 7 + 4 × 10 = 96 | 100 |
| All | ||||
| Specimens | 29,481 | 32,592 | 20,250 | 82,323 |
| COI sequences (% success) | 25,217 (85.6%) | 28,923 (88.7%) | 16,152 (79.8%) | 70,293 (85.4%) |
| BINs (% new to BOLD) | 2565 (19.4%) | 3870 (15.8%) | 4043 (23.0%) | 8790 (23.8%) |
| Diptera | ||||
| Specimens (% of all specimens) | 23,114 (78%) | 15,448 (47%) | 14,238 (70%) | 52,800 (64%) |
| COI sequences (% success) | 20,909 (91%) | 14,983 (97%) | 12,338 (87%) | 48,230 (91%) |
| BINs (in % of all BINs) | 1571 (61%) | 1676 (43%) | 2632 (65%) | 4863 (55%) |
| Diptera BINs new to BOLD | 375 | 260 | 736 | 1413 |
| DT BINs new to BOLD (% of all new Diptera BINs) | 337 (90%) | 215 (83%) | 215 (29%) | 780 (55%) |
| Taxa | BINs | Bavarian Species | German Species | Bavarian/German Species | BINs/Bavarian Species | BINs/German Species |
|---|---|---|---|---|---|---|
| Asilidae | 13 | 68 | 85 | 0.80 | 0.19 | 0.15 |
| Calliphoridae | 22 | 50 | 62 | 0.80 | 0.44 | 0.35 |
| Drosophilidae | 27 | 64 | 81 | 0.79 | 0.42 | 0.34 |
| Ephydridae | 32 | 140 | 174 | 0.80 | 0.23 | 0.18 |
| Muscidae | 160 | 267 | 334 | 0.80 | 0.60 | 0.48 |
| Sarcophagidae | 35 | 107 | 134 | 0.80 | 0.33 | 0.26 |
| Stratiomyidae | 14 | 59 | 71 | 0.83 | 0.24 | 0.20 |
| Syrphidae | 131 | 389 | 458 | 0.85 | 0.34 | 0.29 |
| Tabanidae | 9 | 47 | 58 | 0.81 | 0.19 | 0.16 |
| Tachinidae | 126 | 361 | 501 | 0.72 | 0.35 | 0.25 |
| Tipulidae | 43 | 120 | 142 | 0.85 | 0.36 | 0.30 |
| Average values | 0.33 ± 0.9 | 0.27 ± 0.7 | ||||
| Cecidomyiidae | 1163 | 328 | 859 | 0.38 | 3.55 | 1.35 |
| Chironomidae | 296 | 576 | 781 | 0.74 | 0.51 | 0.38 |
| Phoridae | 348 | 302 | 378 | 0.80 | 1.15 | 0.92 |
| Sciaridae | 339 | 231 | 343 | 0.72 | 1.47 | 0.99 |
| Average values | 1.67 ± 0.9 | 0.91 ± 0.3 | ||||
| All Diptera | 4863 | 7635 | 9544 | 0.80 | 0.64 | 0.51 |
| Taxon | BINs | Chao1 | BIN/Chao1 | Bavarian Species | German Species | Chao1/ Bavarian Species | Chao1/ German Species |
|---|---|---|---|---|---|---|---|
| Asilidae | 13 | 16 | 0.81 | 68 | 85 | 0.24 | 0.16 |
| Calliphoridae | 22 | 28 | 0.79 | 50 | 62 | 0.56 | 0.45 |
| Drosophilidae | 27 | 38 | 0.71 | 64 | 81 | 0.59 | 0.47 |
| Ephydridae | 32 | 88 | 0.36 | 140 | 174 | 0.63 | 0.51 |
| Muscidae | 160 | 220 | 0.73 | 267 | 334 | 0.82 | 0.66 |
| Sarcophagidae | 35 | 41 | 0.85 | 107 | 134 | 0.38 | 0.31 |
| Stratiomyidae | 14 | 16 | 0.88 | 59 | 71 | 0.27 | 0.23 |
| Syrphidae | 131 | 158 | 0.83 | 389 | 458 | 0.41 | 0.34 |
| Tabanidae | 9 | 10 | 0.90 | 47 | 58 | 0.21 | 0.17 |
| Tachinidae | 126 | 153 | 0.82 | 361 | 501 | 0.42 | 0.31 |
| Tipulidae | 43 | 59 | 0.73 | 120 | 142 | 0.49 | 0.42 |
| Average values | 0.46 ± 0.2 | 0.37 ± 0.2 | |||||
| Cecidomyiidae | 1163 | 1937 | 0.60 | 328 | 859 | 5.91 | 2.25 |
| Chironomidae | 296 | 479 | 0.62 | 576 | 781 | 0.83 | 0.61 |
| Phoridae | 348 | 432 | 0.81 | 302 | 378 | 1.43 | 1.14 |
| Sciaridae | 339 | 468 | 0.72 | 231 | 343 | 2.03 | 1.36 |
| Average values | 2.55 ± 1.7 | 1.34 ± 0.5 | |||||
| All Diptera | 4863 | 6927 | 0.70 | 7635 | 9544 | 0.91 | 0.73 |
| Dark Taxa | BINs | Estimates Bavaria | Bavarian Species | New Records Bavaria | Estimates Germany | German Species | New Records Germany |
|---|---|---|---|---|---|---|---|
| BIN/species ratio | |||||||
| Cecidomyiidae | 1163 | 1817–3524 | 328 | 1489–3196 | 2280–4307 | 859 | 1421–3448 |
| Chironomidae | 296 | 463–897 | 576 | 0–321 | 580–1096 | 781 | 0–315 |
| Phoridae | 348 | 544–1055 | 302 | 242–753 | 682–1289 | 378 | 304–911 |
| Sciaridae | 339 | 530–1027 | 231 | 299–796 | 665–1256 | 343 | 322–913 |
| Chao1/species ratio | |||||||
| Cecidomyiidae | 1937 | 2129–4211 | 328 | 1801–3883 | 2653–5235 | 859 | 1794–4376 |
| Chironomidae | 479 | 526–1041 | 576 | 0–465 | 656–1295 | 781 | 0–514 |
| Phoridae | 432 | 475–939 | 302 | 173–637 | 592–1168 | 378 | 214–790 |
| Sciaridae | 468 | 514–1017 | 231 | 283–786 | 641–1265 | 343 | 298–922 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimeno, C.; Hausmann, A.; Schmidt, S.; Raupach, M.J.; Doczkal, D.; Baranov, V.; Hübner, J.; Höcherl, A.; Albrecht, R.; Jaschhof, M.; et al. Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany. Insects 2022, 13, 82. https://doi.org/10.3390/insects13010082
Chimeno C, Hausmann A, Schmidt S, Raupach MJ, Doczkal D, Baranov V, Hübner J, Höcherl A, Albrecht R, Jaschhof M, et al. Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany. Insects. 2022; 13(1):82. https://doi.org/10.3390/insects13010082
Chicago/Turabian StyleChimeno, Caroline, Axel Hausmann, Stefan Schmidt, Michael J. Raupach, Dieter Doczkal, Viktor Baranov, Jeremy Hübner, Amelie Höcherl, Rosa Albrecht, Mathias Jaschhof, and et al. 2022. "Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany" Insects 13, no. 1: 82. https://doi.org/10.3390/insects13010082
APA StyleChimeno, C., Hausmann, A., Schmidt, S., Raupach, M. J., Doczkal, D., Baranov, V., Hübner, J., Höcherl, A., Albrecht, R., Jaschhof, M., Haszprunar, G., & Hebert, P. D. N. (2022). Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany. Insects, 13(1), 82. https://doi.org/10.3390/insects13010082