
Standard Methods for Dissection of Varroa destructor Females


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Material, Methods and Results of the Dissection Protocols
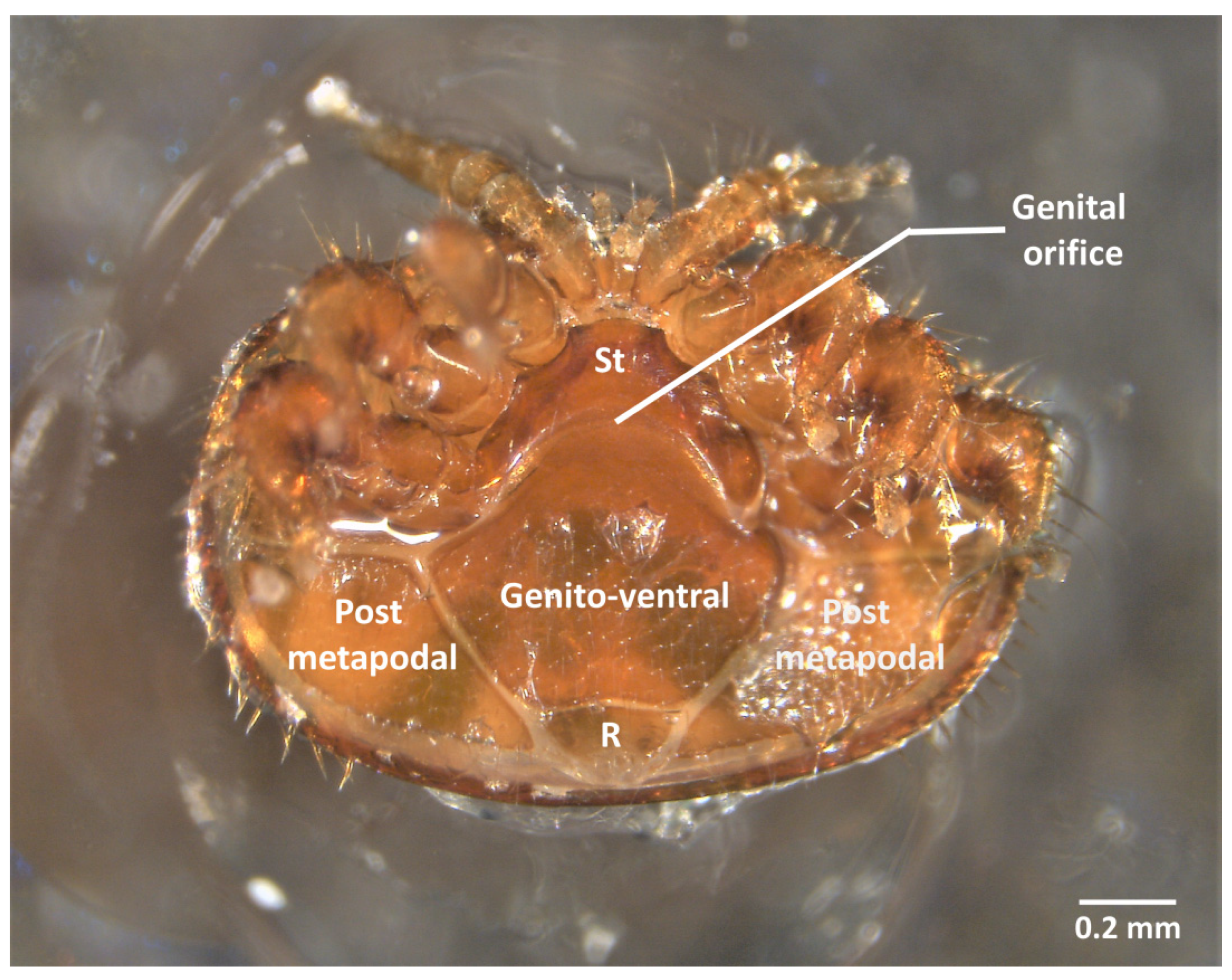
2.1. Morphology and Material
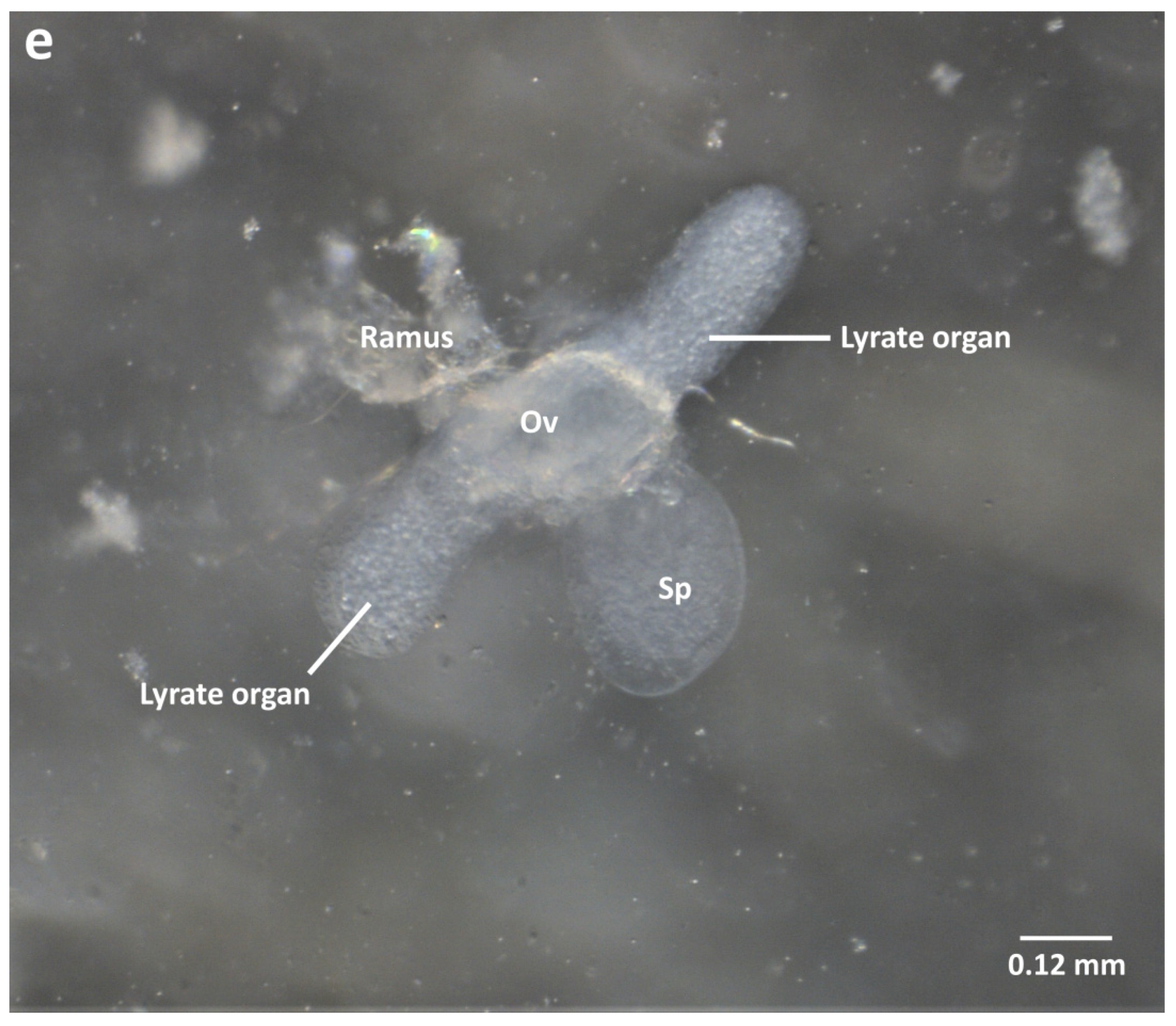
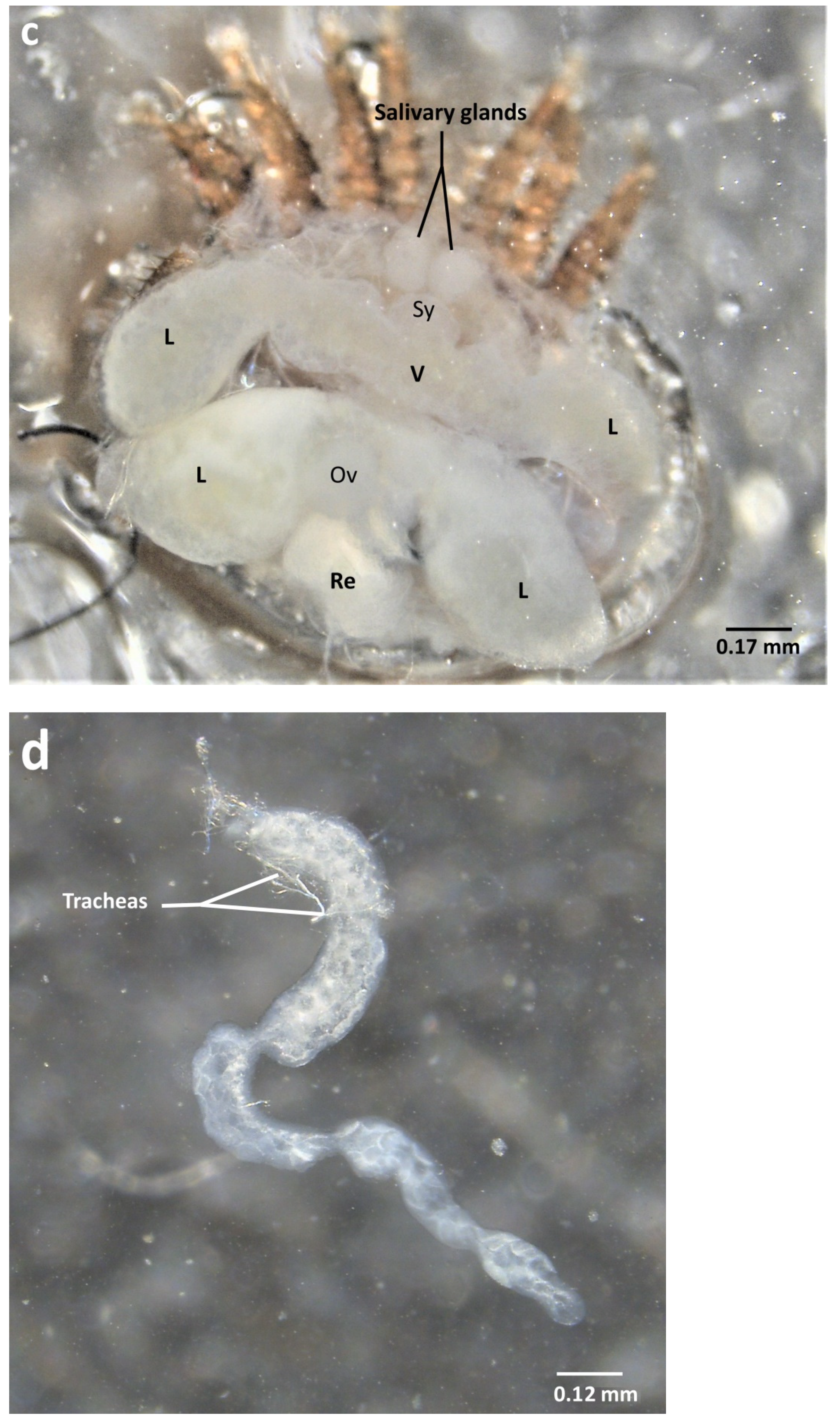
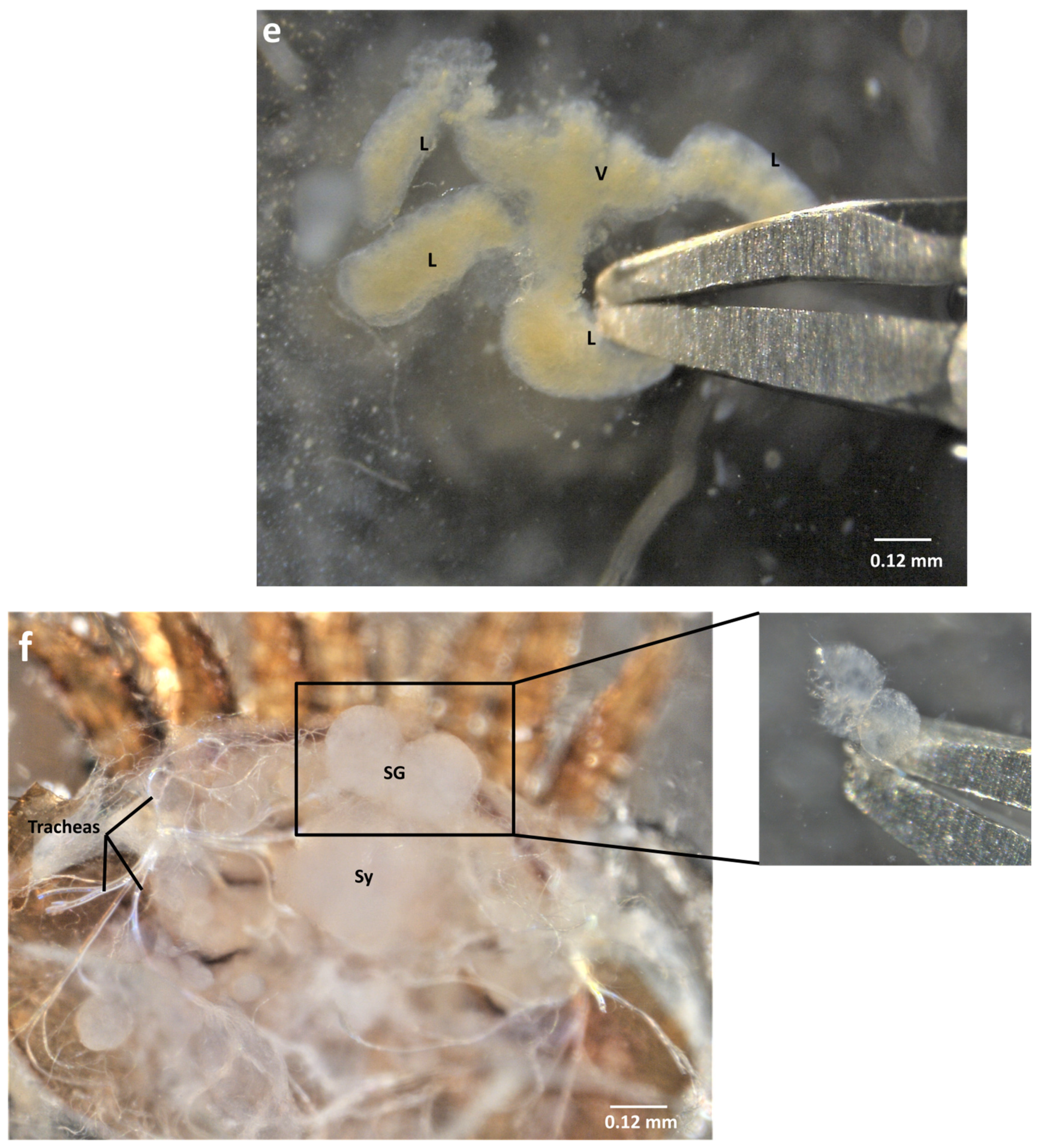
2.2. Ventral Dissection for the Isolation of Reproductive Organs
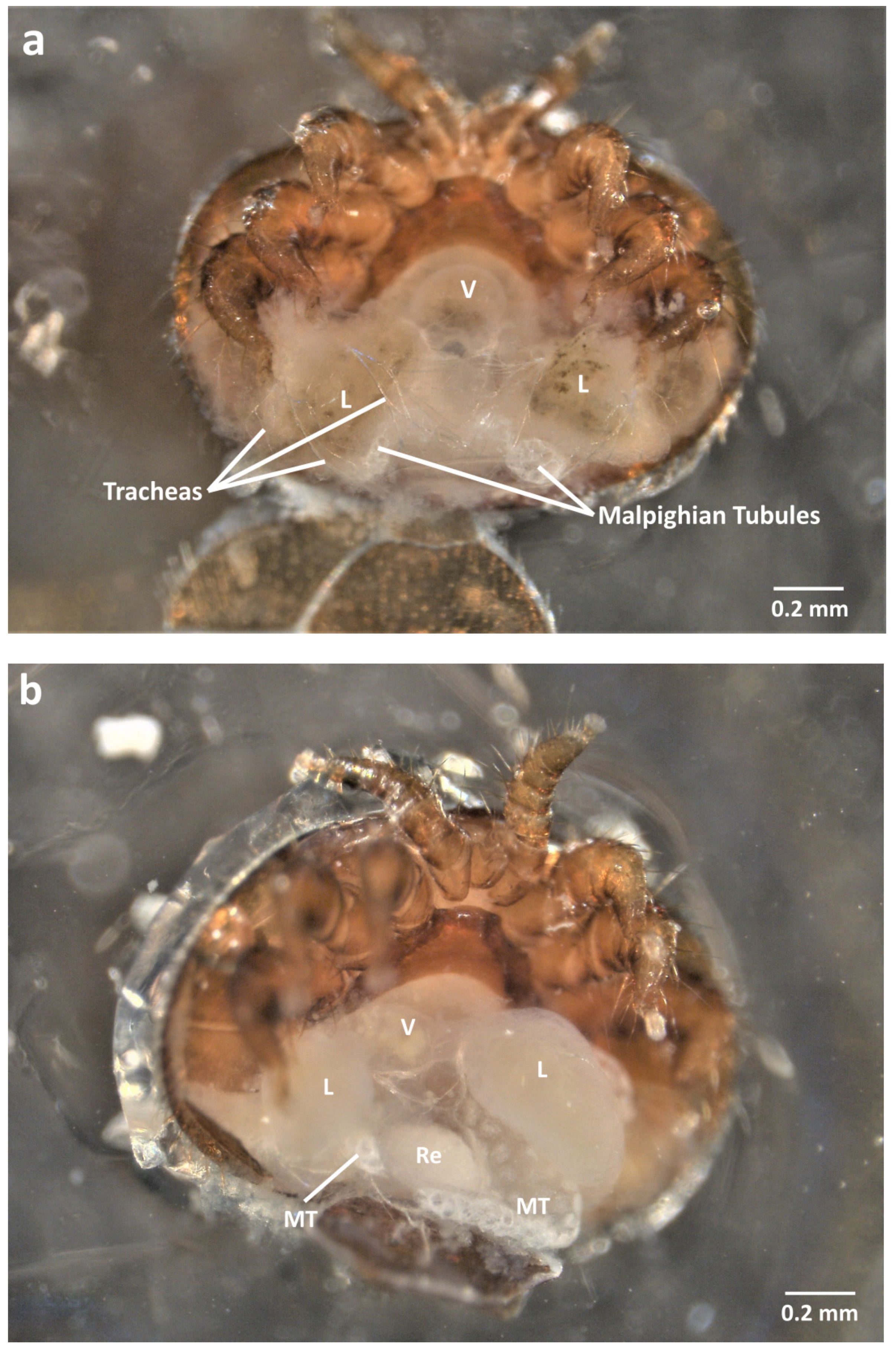
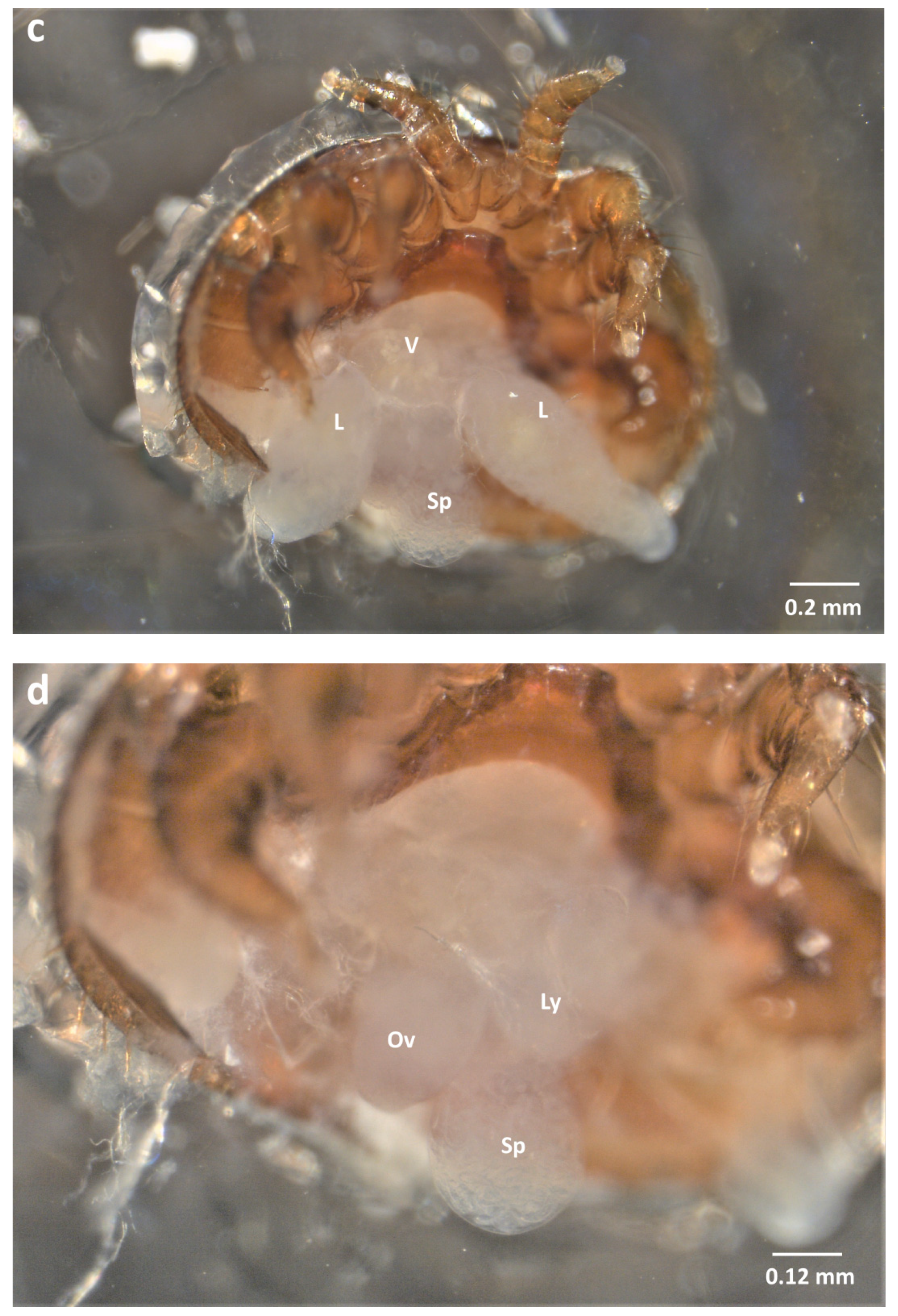
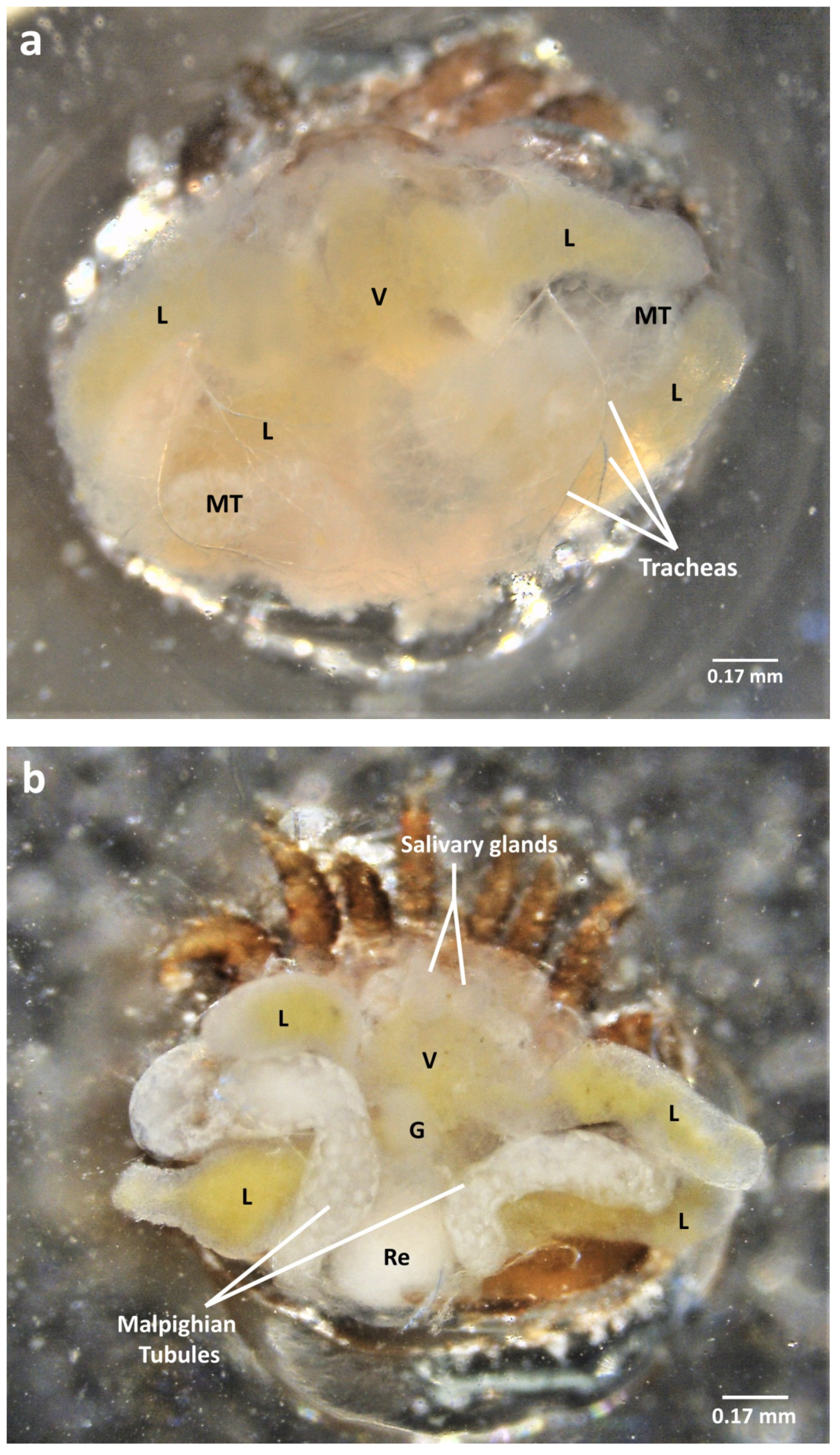
2.3. Dorsal Dissection
2.4. Organ Sampling and Preservation
3. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 10, S96–S119. [Google Scholar] [CrossRef]
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa destructor: A Complex Parasite, Crippling Honey Bees Worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa destructor from the laboratory to the field: Control, biocontrol and ipm perspectives—A review. Insects 2021, 12, 800. [Google Scholar] [CrossRef]
- Corrêa-Marques, M.H.; Medina, L.M.; Martin, S.J.; De Jong, D. Comparing data on the reproduction of Varroa destructor. Genet. Mol. Res. 2003, 2, 1–6. [Google Scholar]
- Francis, R.M.; Nielsen, S.L.; Kryger, P. Varroa-Virus Interaction in Collapsing Honey Bee Colonies. PLoS ONE 2013, 8, e57540. [Google Scholar] [CrossRef]
- Frey, E.; Odemer, R.; Blum, T.; Rosenkranz, P. Activation and interruption of the reproduction of Varroa destructor is triggered by host signals (Apis mellifera). J. Invertebr. Pathol. 2013, 113, 56–62. [Google Scholar] [CrossRef]
- Gisder, S.; Genersch, E. Direct Evidence for Infection of Varroa destructor Mites with the Bee-Pathogenic Deformed Wing Virus Variant B, but Not Variant A, via Fluorescence In Situ Hybridization Analysis. J. Virol. 2021, 95, e01786-20. [Google Scholar] [CrossRef]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed wing virus: Replication and viral load in mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef]
- Piou, V.; Tabart, J.; Urrutia, V.; Hemptinne, J.L.; Vétillard, A. Impact of the phoretic phase on reproduction and damage caused by Varroa destructor (Anderson and Trueman) to its host, the european honey bee (Apis mellifera L.). PLoS ONE 2016, 11, e0153482. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Tatè, R.; Campbell, E.M.; Gigliotti, S.; Bowman, A.S.; Pennacchio, F. A salivary chitinase of Varroa destructor influences host immunity and mite’s survival. PLoS Pathog. 2020, 16, e1009075. [Google Scholar] [CrossRef]
- Cabrera, A.R.; Shirk, P.D.; Duehl, A.J.; Donohue, K.V.; Grozinger, C.M.; Evans, J.D.; Teal, P.E.A. Genomic organization and reproductive regulation of a large lipid transfer protein in the varroa mite, Varroa destructor (Anderson & Trueman). Insect Mol. Biol. 2013, 22, 505–522. [Google Scholar] [CrossRef]
- Campbell, E.M.; Budge, G.E.; Watkins, M.; Bowman, A.S. Transcriptome analysis of the synganglion from the honey bee mite, Varroa destructor and RNAi knockdown of neural peptide targets. Insect Biochem. Mol. Biol. 2016, 70, 116–126. [Google Scholar] [CrossRef]
- Häußermann, C.K.; Giacobino, A.; Munz, R.; Ziegelmann, B.; Palacio, M.A.; Rosenkranz, P. Reproductive parameters of female Varroa destructor and the impact of mating in worker brood of Apis mellifera. Apidologie 2020, 51, 342–355. [Google Scholar] [CrossRef]
- Häußermann, C.K.; Ziegelmann, B.; Rosenkranz, P. Spermatozoa capacitation in female Varroa destructor and its influence on the timing and success of female reproduction. Exp. Appl. Acarol. 2016, 69, 371–387. [Google Scholar] [CrossRef]
- Wendling, S.; Guillet, B.; Roy, L.; Kreiter, S.; Colin, M.E. Fertilization and fertility in the female of Varroa destructor, a key point for the parasite population dynamics. Apidologie 2014, 45, 722–732. [Google Scholar] [CrossRef][Green Version]
- Garrido, C.; Rosenkranz, P.; Stürmer, M.; Rübsam, R.; Büning, J. Toluidine blue staining as a rapid measure for initiation of oocyte growth and fertility in Varroa jacobsoni Oud. Apidologie 2000, 31, 559–566. [Google Scholar] [CrossRef][Green Version]
- Steiner, J.; Dittmann, F.; Rosenkranz, P.; Engels, W. The first gonocycle of the parasitic mite (Varroa jacobsoni) in relation to preimaginal development of its host, the honey bee (Apis mellifera camica). Invertebr. Reprod. Dev. 1994, 25, 175–183. [Google Scholar] [CrossRef]
- Campbell, E.M.; Budge, G.E.; Bowman, A.S. Gene-knockdown in the honey bee mite Varroa destructor by a non-invasive approach: Studies on a glutathione S-transferase. Parasites Vectors 2010, 3, 73. [Google Scholar] [CrossRef]
- Carreck, N.L.; Andree, M.; Brent, C.S.; Cox-Foster, D.; Dade, H.A.; Ellis, J.D.; Hatjina, F.; Van Englesdorp, D. Standard methods for Apis mellifera anatomy and dissection. J. Apic. Res. 2013, 52, 1–40. [Google Scholar] [CrossRef]
- Hubert, J.; Bicianova, M.; Ledvinka, O.; Kamler, M.; Lester, P.J.; Nesvorna, M.; Kopecky, J.; Erban, T. Changes in the Bacteriome of Honey Bees Associated with the Parasite Varroa destructor, and Pathogens Nosema and Lotmaria passim. Microb. Ecol. 2017, 73, 685–698. [Google Scholar] [CrossRef]
- Wu, J.L.; Zhou, C.X.; Wu, P.J.; Xu, J.; Guo, Y.Q.; Xue, F.; Getachew, A.; Xu, S.F. Brain metabolomic profiling of eastern honey bee (Apis cerana) infested with the mite Varroa destructor. PLoS ONE 2017, 12, e0175573. [Google Scholar] [CrossRef]
- Cabrera, A.R.; Shirk, P.D.; Teal, P.E.A.; Grozinger, C.M.; Evans, J.D. Examining the role of foraging and malvolio in host-finding behavior in the honey bee parasite, Varroa destructor (Anderson & Trueman). Arch. Insect Biochem. Physiol. 2014, 85, 61–75. [Google Scholar] [CrossRef]
- Cabrera, A.R.; Shirk, P.D.; Evans, J.D.; Hung, K.; Sims, J.; Alborn, H.; Teal, P.E.A. Three Halloween genes from the Varroa mite, Varroa destructor (Anderson & Trueman) and their expression during reproduction. Insect Mol. Biol. 2015, 24, 277–292. [Google Scholar] [CrossRef]
- Gusachenko, O.N.; Woodford, L.; Balbirnie-Cumming, K.; Campbell, E.M.; Christie, C.R.; Bowman, A.S.; Evans, D.J. Green Bees: Reverse Genetic Analysis of Deformed Wing Virus Transmission, Replication, and Tropism. Viruses 2020, 12, 532. [Google Scholar] [CrossRef]
- Anderson, D.L.; Trueman, J.W. Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- Edwards, K.T.; Goddard, J.; Varela-Stokes, A.S. Examination of the internal morphology of the ixodid tick, Amblyomma maculatum Koch, (Acari: Ixodidae); a “How-to” pictorial dissection guide. Midsouth Entomol. 2009, 2, 28–39. [Google Scholar]
- Grabowski, J.M.; Kissinger, R. Ixodid Tick Dissection and Tick Ex Vivo Organ Cultures for Tick-Borne Virus Research. Curr. Protoc. Microbiol. 2020, 59, e118. [Google Scholar] [CrossRef]
- Akimov, I.A.; Benedyk, S.V.; Zaloznaya, L.M. Complex analysis of morphological characters of gamasid mite Varroa destructor (Parasitiformes, Varroidae). Vestn. Zool. 2004, 38, 57–66. [Google Scholar]
- Yevstafieva, V.O.; Zaloznaya, L.M.; Nazarenko, O.S.; Melnychuk, V.V.; Sobolta, A.G. Morphological variation of Varroa destructor (Parasitiformes, Varroidae) in different seasons. Biosyst. Divers. 2020, 28, 18–23. [Google Scholar] [CrossRef]
- Kirrane, M.J.; De Guzman, L.I.; Rinderer, T.E.; Frake, A.M.; Wagnitz, J.; Whelan, P.M. Asynchronous development of honey bee host and Varroa destructor (Mesostigmata: Varroidae) influences reproductive potential of mites. J. Econ. Entomol. 2011, 104, 1146–1152. [Google Scholar] [CrossRef]
- Richard, D.; Colin, M.E.; Lhomme, M. Anatomical organization of the tracheal system of Varroa jacobsoni (Acari: Varroidae). Exp. Appl. Acarol. 1990, 9, 63–72. [Google Scholar] [CrossRef]
- Wendling, S.L.P. Varroa destructor (Anderson et Trueman, 2000), un Acarien Ectoparasite de l’abeille Domestique Apis mellifera Linnaeus, 1758. Revue Bibliographique et Contribution à l’étude de sa Reproduction. Ph.D. Thesis, Ecole Vétérinaire d’Alfort, Maison-Alfort, France, 2012; 190p. [Google Scholar]
- Posada-Florez, F.; Sonenshine, D.E.; Egekwu, N.I.; Rice, C.; Lupitskyy, R.; Cook, S.C. Insights into the metabolism and behaviour of Varroa destructor mites from analysis of their waste excretions. Parasitology 2019, 146, 527–532. [Google Scholar] [CrossRef]
- Alberti, G.; Hänel, H. Fine structure of the genital system in the bee parasite, Varroa jacobsoni (Gamasida: Dermanyssina) with remarks on spermiogenesis, spermatozoa and capacitation. Exp. Appl. Acarol. 1986, 2, 63–104. [Google Scholar] [CrossRef]
- Steiner, J.; Diehl, P.A.; Vlimant, M. Vitellogenesis in Varroa jacobsoni, a parasite of honey bees. Exp. Appl. Acarol. 1995, 19, 411–422. [Google Scholar] [CrossRef]
- Alberti, G.; Zeck-Kapp, G. The Nutrimentary Egg Development of the Mite, Varroa jacobsoni (Acari, Arachnida), an Ectoparasite of Honey Bees. Acta Zool. 1986, 67, 11–25. [Google Scholar] [CrossRef]
- Eliash, N.; Thangarajan, S.; Goldenberg, I.; Sela, N.; Kupervaser, M.; Barlev, J.; Altman, Y.; Knyazer, A.; Kamer, Y.; Zaidman, I.; et al. Varroa chemosensory proteins: Some are conserved across Arthropoda but others are arachnid specific. Insect Mol. Biol. 2019, 28, 321–341. [Google Scholar] [CrossRef]
- de Ruijter, A.; Kaas, J.P. Anatomy of the Varroa-mite. Varroa jacobsoni Oud. affecting honey bees: Present status and needs. In Proceedings of the Meeting of the EC Experts Group, Wageningen, The Netherlands, 7–9 February 1983. [Google Scholar]
- Koutouvela, E.; Papachristoforou, A. The heart of Varroa destructor: Description, function and inhibition following acaricide application. Syst. Appl. Acarol. 2019, 24, 638–644. [Google Scholar] [CrossRef]
- Tabart, J.; Colin, M.E.; Carayon, J.L.; Tene, N.; Payre, B.; Vetillard, A. Artificial feeding of Varroa destructor through a chitosan membrane: A tool for studying the host-microparasite relationship. Exp. Appl. Acarol. 2013, 61, 107–118. [Google Scholar] [CrossRef]
- Posada-Florez, F.; Ryabov, E.V.; Heerman, M.C.; Chen, Y.; Evans, J.D.; Sonenshine, D.E.; Cook, S.C. Varroa destructor mites vector and transmit pathogenic honey bee viruses acquired from an artificial diet. PLoS ONE 2020, 15, e0242688. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piou, V.; Vilarem, C.; Rein, C.; Sprau, L.; Vétillard, A. Standard Methods for Dissection of Varroa destructor Females. Insects 2022, 13, 37. https://doi.org/10.3390/insects13010037
Piou V, Vilarem C, Rein C, Sprau L, Vétillard A. Standard Methods for Dissection of Varroa destructor Females. Insects. 2022; 13(1):37. https://doi.org/10.3390/insects13010037
Chicago/Turabian StylePiou, Vincent, Caroline Vilarem, Carolin Rein, Lina Sprau, and Angélique Vétillard. 2022. "Standard Methods for Dissection of Varroa destructor Females" Insects 13, no. 1: 37. https://doi.org/10.3390/insects13010037
APA StylePiou, V., Vilarem, C., Rein, C., Sprau, L., & Vétillard, A. (2022). Standard Methods for Dissection of Varroa destructor Females. Insects, 13(1), 37. https://doi.org/10.3390/insects13010037