
Efficacy of Anystis baccarum against Foxglove Aphids, Aulacorthum solani, in Laboratory and Small-Scale Greenhouse Trials

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing of Pest Insects
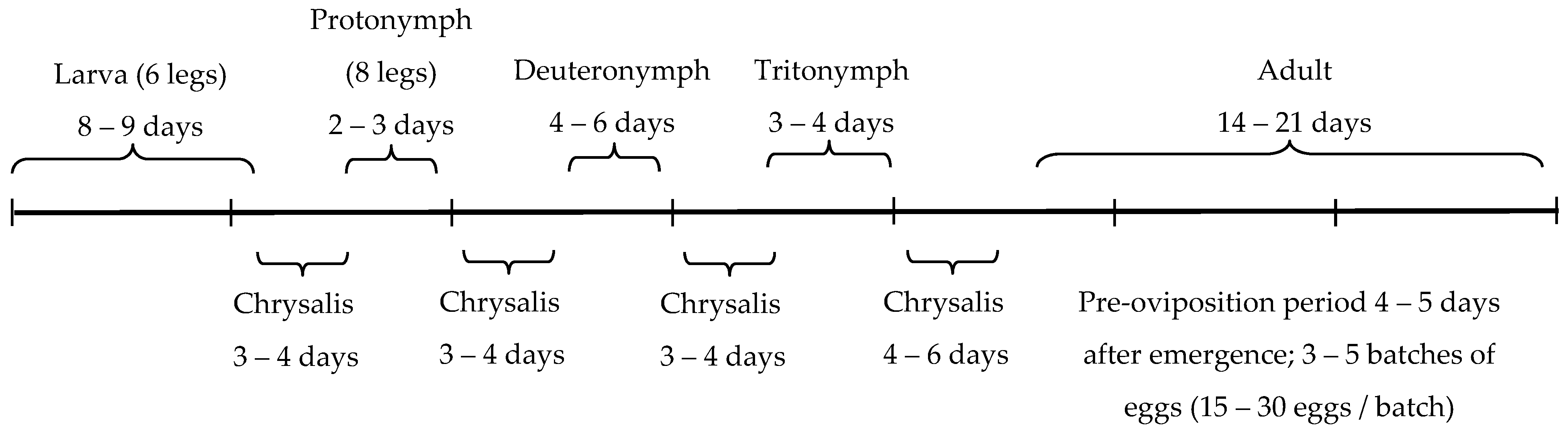
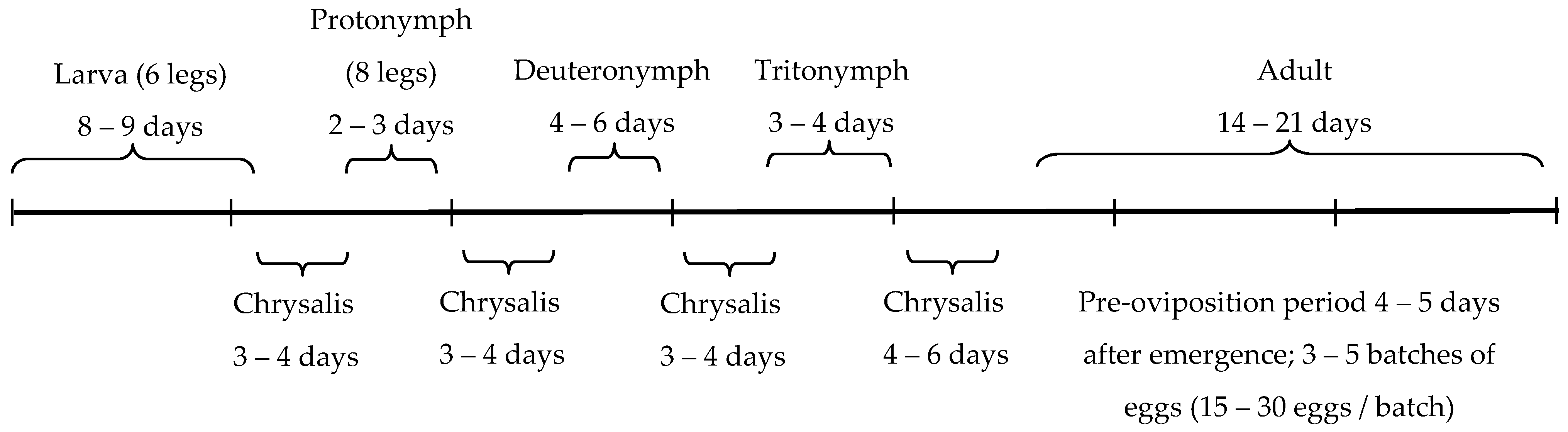
2.2. Predators
2.3. Experimental Designs
2.3.1. Laboratory Cup Trial: Efficacy against Aphids of Mixed Age
2.3.2. Laboratory Cup Trial: Efficacy against 1st Instar Aphids
2.3.3. Laboratory Cage Trial: Effects on Aphid Population Growth on Sweet Pepper
2.3.4. Greenhouse Trial: Effects on Foxglove Aphid Population Dynamics on Sweet Pepper
3. Results
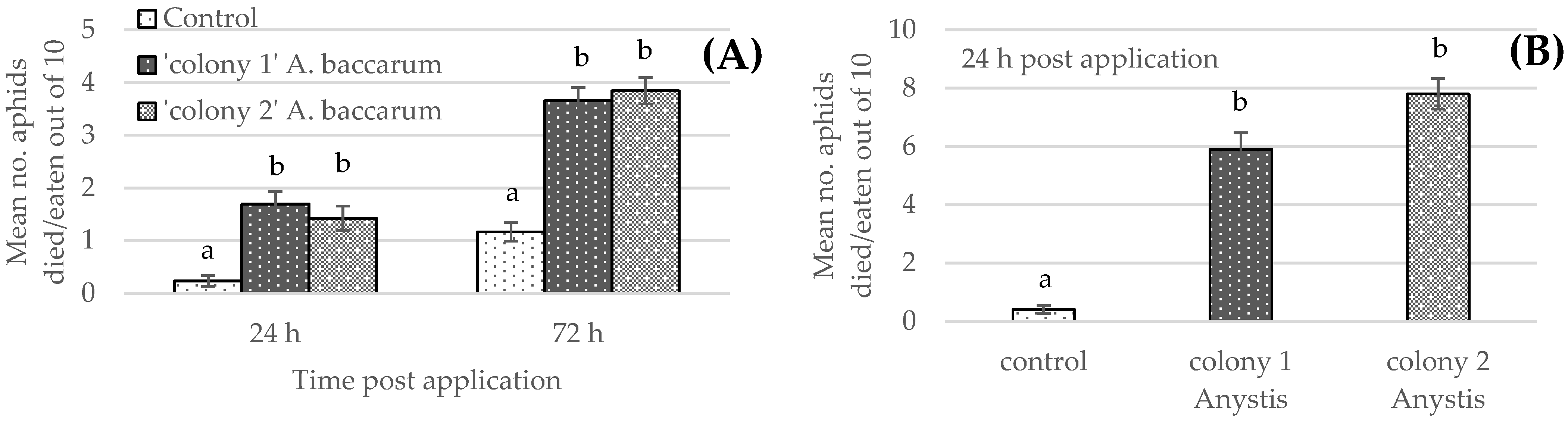
3.1. Laboratory Cup Trial: Efficacy against Aphids of Mixed-Age
3.2. Laboratory Cup Trial: Efficacy against 1st Instar Aphids
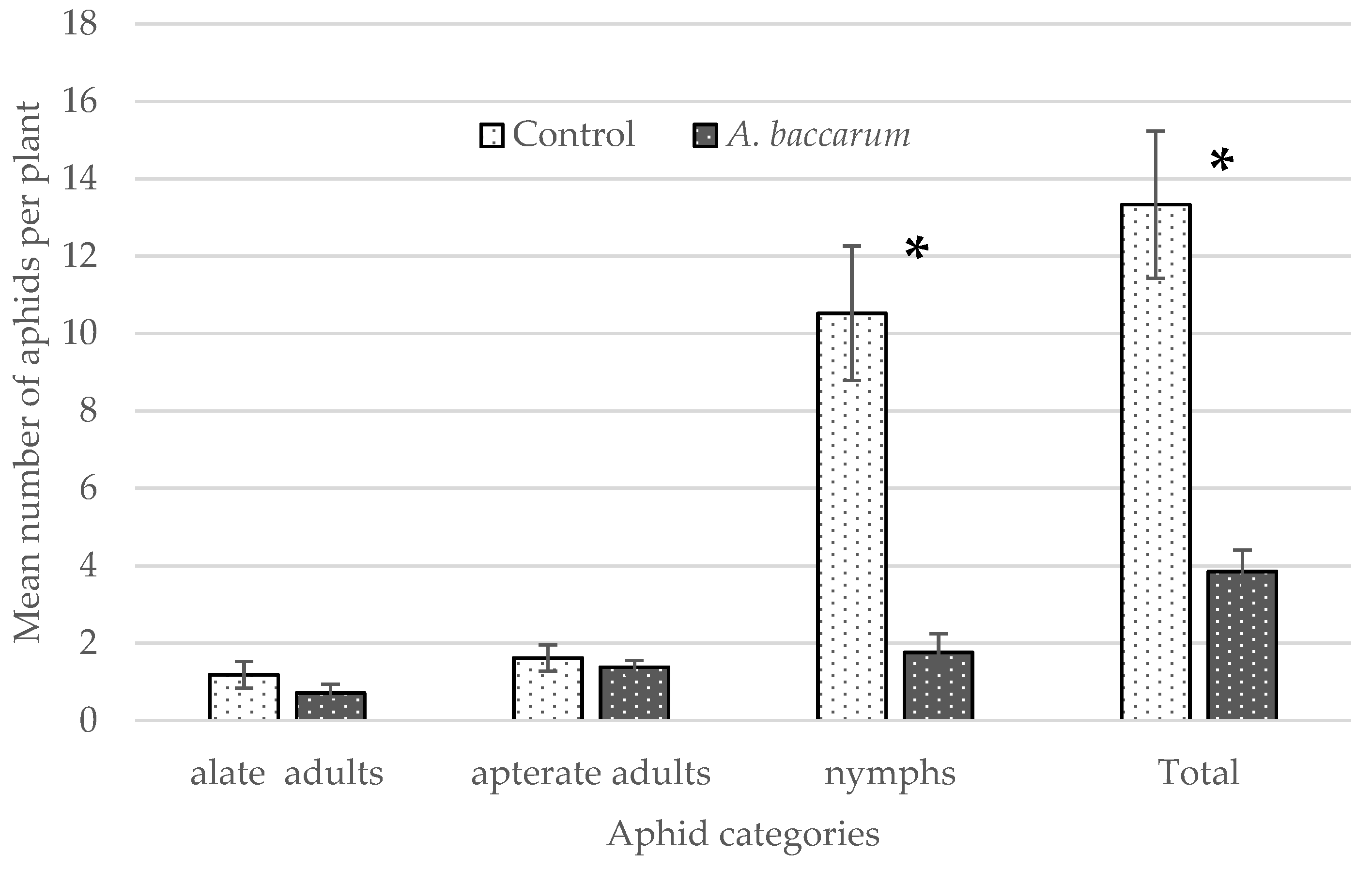
3.3. Laboratory Cage Trial: Effects on Aphid Population Growth
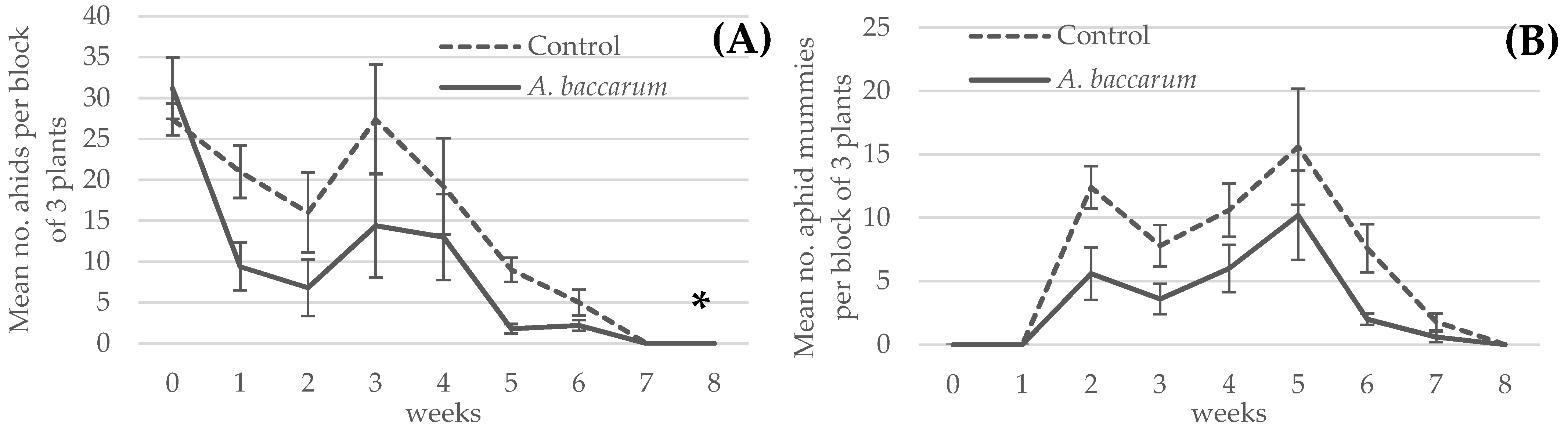
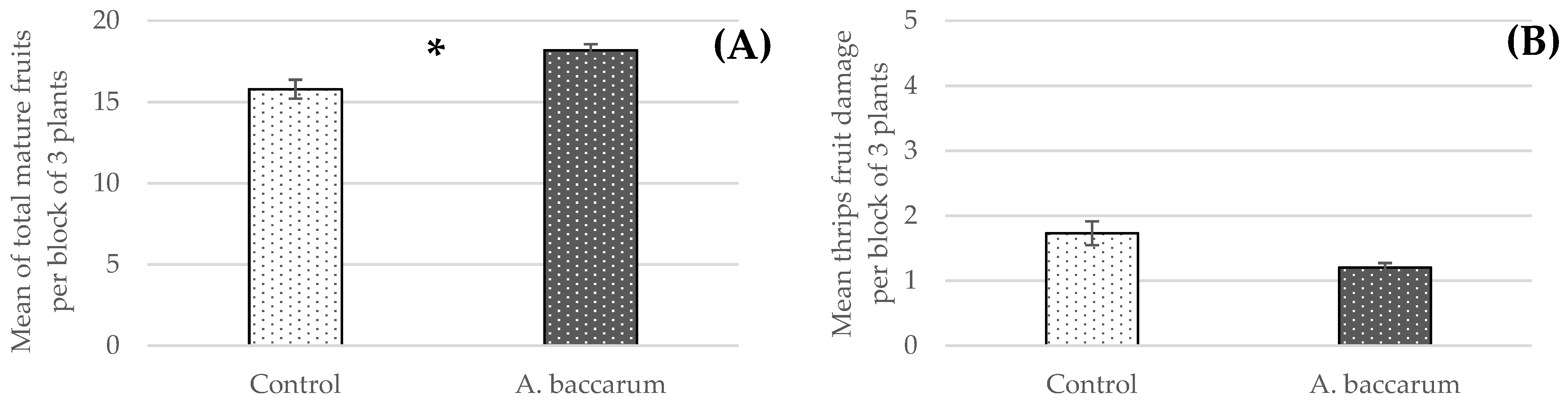
3.4. Greenhouse Trial: Combined Efficacy of the Aphid Parasitoid Aphidius ervi and Anystis baccarum against Foxglove Aphids on Sweet Pepper
4. Discussion
- A. baccarum can be used with A. ervi to control foxglove aphids;
- A. baccarum will readily establish on peppers grown in coco growbags; and
- A. baccarum provides a robust foundation for a biological control program due to its wide host (prey) range and can complement the activity of a more specialized natural enemy (A. ervi).
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cuthbertson, A.G.S.; Murchie, A.K. Ecological benefits of Anystis baccarum in an orchard ecosystem and the need for its conservation. Int. J. Environ. Sci. Technol. 2010, 7, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Cuthbertson, A.G.S.; Qiu, B.L.; Murchie, A.K. Anystis baccarum: An important generalist predatory mite to be considered in apple orchard pest management strategies. Insects 2014, 5, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Geary, I.J.; Merfield, C.N.; Hale, R.J.; Shaw, M.D.; Hodge, S. Predation of nymphal tomato potato psyllid, Bactericera cockerelli (Šulc) (Hemiptera: Triozidae), by the predatory mite, Anystis baccarum L. (Trombidiformes: Anystidae). N. Z. Entomol. 2016, 39, 110–116. [Google Scholar] [CrossRef]
- Chen, L.L.; Yuan, P.; Pozsgai, G.; Chen, P.; Zhu, H.; You, M.-S. The impact of cover crops on the predatory mite Anystis baccarum (Acari, Anystidae) and the leafhopper pest Empoasca onikii (Hemiptera, Cicadellidae) in a tea plantation. Pest Manag. Sci. 2019, 75, 3371–3380. [Google Scholar] [CrossRef] [PubMed]
- Jandricic, S.E.; Wraight, S.P.; Bennett, K.C.; Sanderson, J.P. Developmental times and life table statistics of Aulacorthum solani (Hemiptera: Aphididae) at six constant temperatures, with recommendations on the application of temperature-dependent development models. Environ. Entomol. 2010, 39, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Buitenhuis, R.; Murphy, G.; Shipp, L. Aphis gossypii Glover, melon/cotton aphid, Aulacorthum solani (Kaltenbach), foxglove aphid, and other arthropod pests in greenhouse crops. In Biological Control Programmes in Canada 2001–2012; Mason, P.G., Gillespie, D.R., Eds.; CABI: Wallingford, UK, 2013; pp. 98–112. [Google Scholar]
- La-Spina, M.; Jandricic, S.E.; Buitenhuis, R. Short-term increases in aphid dispersal from defensive dropping do not necessarily affect long-term biological control by parasitoids. J. Econ. Entomol. 2019, 112, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Ann. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Backer, L.; Megido, C.R.; Haubruge, E.; Verheggen, F.J. Macrolophus pygmaeus (Rambur) as an efficient predator of the tomato leafminer Tuta absoluta (Meyrick) in Europe. A review. Biotechnol. Agron. Soc. Environ. 2014, 18, 536–543. [Google Scholar]
- Saito, T.; Brownbridge, M. From promising, to product: Developmental steps and challenges to bring a new predatory mite to market. IOBC-WPRS Bull. 2017, 124, 195–199. [Google Scholar]
- Blümel, S. Biological control of aphids on vegetable crops. In Biocontrol in Protected Culture; Heinz, K.M., Driesche, V., Parella, M.P., Eds.; Ball Publishing: Batavia, IL, USA, 2004; pp. 297–312. [Google Scholar]
- Badii, M.H.; Hernández-Ortiz, E.; Flores, A.E.; Landeros, J. Prey stage preference and functional response of Euseius hibisci to Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae). Exp. Appl. Acarol. 2004, 34, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Kasap, I.; Atlihan, R. Consumption rate and functional response of the predaceous mite Kampimodromus aberrans to two-spotted spider mite Tetranychus urticae in the laboratory. Exp. Appl. Acarol. 2011, 53, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-Y.; Liu, T.-X. Relative consumption of three aphid species by the lacewing, Chrysoperla rufilabris, and effects on its development and survival. BioControl 2001, 46, 481–491. [Google Scholar] [CrossRef]
- Inayat, T.P.; Rana, S.A.; Rana, N.; Ruby, T.; Javaid, M.; Sadiqui, I.; Abbas, M.N. Predation rate in selected coccinellid (coleoptera) predators on some major aphidid and cicadellid (hemipteran) pests. Int. J. Agric. Biol. 2011, 13, 427–430. [Google Scholar]
- Rocca, M.; Messelink, G.J. Combining lacewings and parasitoids for biological control of foxglove aphids in sweet pepper. J. Appl. Entomol. 2017, 141, 402–410. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Complementary suppression of aphids by predators and parasitoids. Biol. Control 2015, 90, 83–91. [Google Scholar] [CrossRef]
- Snyder, W.E.; Ballard, S.N.; Yang, S.; Clevenger, G.M.; Miller, T.D.; Ahn, J.J.; Hatten, T.D.; Berryman, A.A. Complementary biocontrol of aphids by the ladybeetle Harmonia axyridis and the parasitoid Aphelinus asychis on greenhouse roses. Biol. Control 2004, 30, 229–235. [Google Scholar] [CrossRef]
- Henry, L.M.; Gillespie, D.R.; Roitberg, B.D. Does mother really know best? Oviposition preference reduces reproductive performance in the generalist parasitoid Aphidius ervi. Entomol. Exp. Appl. 2005, 116, 167–174. [Google Scholar] [CrossRef]
- Paull, C.A.; Schellhorn, N.A.; HilleRisLAmbers, R.; Austin, A.D. Escape from Parasitoids leave larvae vulnerable to predators and has unexpected outcomes for pest suppression. Basic Appl. Ecol. 2012, 13, 542–550. [Google Scholar] [CrossRef]
- Azzouz, H.; Giordanengo, P.; Wäckers, F.L.; Kaiser, L. Effects of feeding frequency and sugar concentration on behavior and longevity of the adult aphid parasitoid: Aphidius ervi (Haliday) (Hymenoptera: Braconidae). Biol. Control 2004, 31, 445–452. [Google Scholar] [CrossRef]
- Hogervorst, P.A.M.; Wäckers, F.L.; Romeis, J. Effects of honeydew sugar composition on the longevity of Aphidius ervi. Entomol. Exp. Appl. 2007, 122, 223–232. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, T.; Brownbridge, M. Efficacy of Anystis baccarum against Foxglove Aphids, Aulacorthum solani, in Laboratory and Small-Scale Greenhouse Trials. Insects 2021, 12, 709. https://doi.org/10.3390/insects12080709
Saito T, Brownbridge M. Efficacy of Anystis baccarum against Foxglove Aphids, Aulacorthum solani, in Laboratory and Small-Scale Greenhouse Trials. Insects. 2021; 12(8):709. https://doi.org/10.3390/insects12080709
Chicago/Turabian StyleSaito, Taro, and Michael Brownbridge. 2021. "Efficacy of Anystis baccarum against Foxglove Aphids, Aulacorthum solani, in Laboratory and Small-Scale Greenhouse Trials" Insects 12, no. 8: 709. https://doi.org/10.3390/insects12080709
APA StyleSaito, T., & Brownbridge, M. (2021). Efficacy of Anystis baccarum against Foxglove Aphids, Aulacorthum solani, in Laboratory and Small-Scale Greenhouse Trials. Insects, 12(8), 709. https://doi.org/10.3390/insects12080709