
The Cotton Mealybug Is Spreading along the Mediterranean: First Pest Detection in Italian Tomatoes




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Sampling
2.2. Morphological Identification
2.3. Molecular Identification
2.4. Phylogenetic Analyses
3. Results
3.1. Insect Sampling
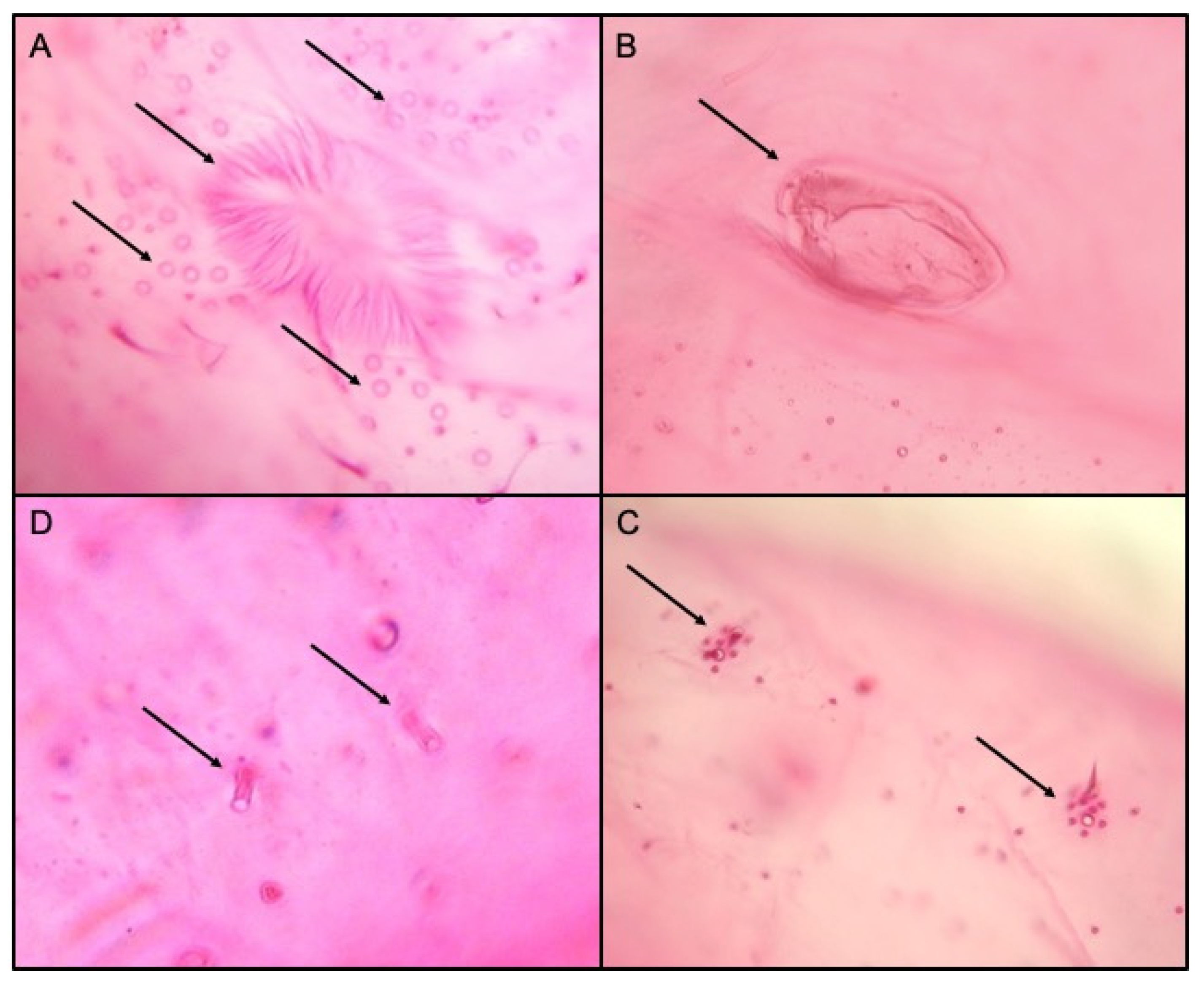
3.2. Morphological Identification
3.3. Molecular Identification
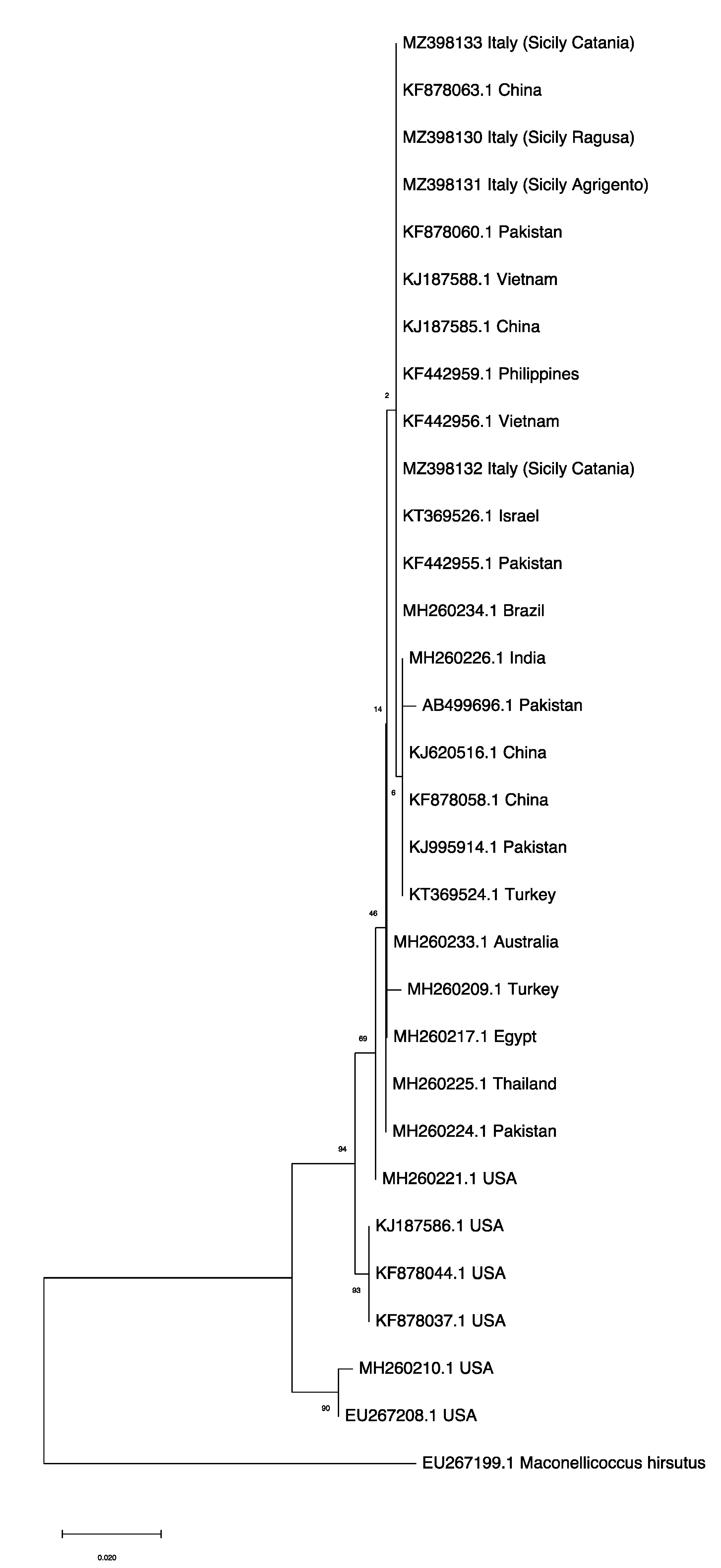
3.4. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E.; Bacher, S.; Kenis, M.; Klotz, S.; Kühn, I.; Minchin, D.; Nentwig, W.; Olenin, S.; Panov, V.; Pergl, J.; et al. Grasping at the routes of biological invasions: A framework for integrating pathways into policy. J. Appl. Ecol. 2008, 45, 403–414. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, Y.; Zhang, H.; Liu, J.; Jiang, Y.; Wyckhuys, K.A.G.; Wu, K. Global warming modifies long-distance migration of an agricultural insect pest. J. Pest Sci. 2020, 93, 569–581. [Google Scholar] [CrossRef]
- Charles, H.; Dukes, J.S. Impacts of Invasive Species on Ecosystem Services. In Biological Invasions; Springer: Berlin/Heidelberg, Germany, 2007; pp. 217–237. [Google Scholar]
- Boughdad, A.; Haddi, K.; El Bouazzati, A.; Nassiri, A.; Tahiri, A.; El Anbri, C.; Eddaya, T.; Zaid, A.; Biondi, A. First record of the invasive spotted wing Drosophila infesting berry crops in Africa. J. Pest Sci. 2021, 94, 261–271. [Google Scholar] [CrossRef]
- Gugliuzzo, A.; Biedermann, P.H.W.; Carrillo, D.; Castrillo, L.A.; Egonyu, J.P.; Gallego, D.; Haddi, K.; Hulcr, J.; Jactel, H.; Kajimura, H.; et al. Recent advances toward the sustainable management of invasive Xylosandrus ambrosia beetles. J. Pest Sci. 2021, 94, 615–637. [Google Scholar] [CrossRef]
- Pellizzari, G.; Germain, J.F. Scales (Hemiptera, Superfamily Coccoidea). Eur. BioRisk 2010, 3, 475–510. [Google Scholar] [CrossRef]
- Miller, D.R.; Miller, G.L.; Watson, G.W. Invasive species of mealybugs (Hemiptera: Pseudococcidae). Proc. Entomol. Soc. Wash. 2002, 104, 825–836. [Google Scholar]
- Fand, B.B.; Suroshe, S.S. The invasive mealybug Phenacoccus solenopsis Tinsley, a threat to tropical and subtropical agricultural and horticultural production systems—A review. Crop Prot. 2015, 69, 34–43. [Google Scholar] [CrossRef]
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database 2016, 2016, 118. [Google Scholar] [CrossRef]
- Zhou, A.; Lu, Y.; Zeng, L.; Xu, Y.; Liang, G. Does mutualism drive the invasion of two alien species? The case of Solenopsis invicta and Phenacoccus solenopsis. PLoS ONE 2012, 7, e41856. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.S.; Yang, L.; Huang, L.F.; Wang, W.L.; Hu, Y.; Jiang, J.J.; Zhou, Z.S. Temperature- and relative humidity-dependent life history traits of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) on Hibiscus rosa-sinensis (Malvales: Malvaceae). Environ. Entomol. 2015, 44, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Abd-Rabou, S.; Shalaby, H.; Germain, J.F.; Ris, N.; Kreiter, P.; Malausa, T. Identification of mealybug pest species (Hemiptera: Pseudococcidae) in Egypt and France, using a DNA barcoding approach. Bull. Entomol. Res. 2012, 102, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Watson, G.W.; Sun, Y.; Tan, Y.; Xiao, L.; Bai, L. Phenotypic variation and identification of Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in China. Zootaxa 2014, 3802, 109–121. [Google Scholar] [CrossRef][Green Version]
- Thomas, A.; Ramamurthy, V.V. Morphological and molecular studies on the intraspecific variations between populations of the cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Entomol. News 2014, 123, 339–347. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Ma, J.; Qiu, B.L.; He, R.R.; Wu, M.T.; Liang, F.; Zhao, J.P.; Lin, L.; Hu, X.N.; Lv, L.H.; et al. Genetic record for a recent invasion of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) in Asia. Environ. Entomol. 2015, 44, 907–918. [Google Scholar] [CrossRef]
- Wu, F.Z.; Ma, J.; Hu, X.N.; Zeng, L. Homology difference analysis of invasive mealybug species Phenacoccus solenopsis Tinsley in Southern China with COI gene sequence variability. Bull. Entomol. Res. 2015, 105, 32–39. [Google Scholar] [CrossRef]
- Hodgson, C.; Abbas, G.; Arif, M.; Karar, H. Phenacoccus solenopsis Tinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation. Zootaxa 2008, 1913, 1–35. [Google Scholar] [CrossRef]
- Tinsley, J.B. An ants’-nest coccid from New Mexico. Can. Entomol. 1898, 30, 47–48. [Google Scholar] [CrossRef]
- Wang, Y.; Watson, G.W.; Zhang, R. The potential distribution of an invasive mealybug Phenacoccus solenopsis and its threat to cotton in Asia. Agric. For. Entomol. 2010, 12, 403–416. [Google Scholar] [CrossRef]
- Abd-Rabou, S.; Germain, J.-F.; Malausa, T. Phenacoccus parvus Morrison et P. solenopsis Tinsley, deux Cochenilles nouvelles pour l’Egypte (Hemiptera, Pseudococcidae). Bull. Société Entomol. Fr. 2010, 115, 509–510. [Google Scholar]
- Kaydan, M.B.; Çalişkan, A.F.; Ulusoy, M.R. New record of invasive mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Turkey. EPPO Bull. 2013, 43, 169–171. [Google Scholar] [CrossRef]
- Gavrilov-Zimin, I.A.; Danzig, E.M. Some additions to the mealybug fauna (Homoptera: Coccinea: Pseudococcidae) of the Canary Islands. Zoosyst. Ross. 2015, 24, 94–98. [Google Scholar] [CrossRef]
- Pellizzari, G.; Porcelli, F. Aspidiotus bornmuelleri Lindinger, 1911, Rev. Comb. (Hemiptera: Coccomorpha: Diaspididae), a neglected endemic species from Macaronesia, with comments on the genus Cryptophyllaspis, and further notes on the scale insect fauna of Canary Islands, Spain. Zootaxa 2017, 4300, 99–110. [Google Scholar] [CrossRef]
- Spodek, M.; Ben-Dov, Y.; Mondaca, L.; Protasov, A.; Erel, E.; Mendel, Z. The cotton mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Israel: Pest status, host plants and natural enemies. Phytoparasitica 2018, 46, 45–55. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Moharum, F.A.; Abd El-Ghany, N.M. The cotton mealybug Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) as a new insect pest on tomato plants in Egypt. J. Plant Prot. Res. 2015, 55, 48–51. [Google Scholar] [CrossRef]
- Çalışkan-Keçe, A.F.; Ulaşlı, B.T.; Ulusoy, M. Mealybugs (Hemiptera: Coccomorpha: Pseudococcidae) on ornamental plants in eastern Mediterranean region, Turkey. Acta Hortic. 2020, 1269, 231–240. [Google Scholar] [CrossRef]
- Katbeh Bader, A.; Al-Jboory, I.J. First record of cotton mealybug, Phenacoccus solenopsis Tinsley 1898 (Hemiptera: Pseudococcidae), from Saudi Arabia. EPPO Bull. 2020, 50, 557–560. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Jacques, M.A.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; Navas-Cortes, J.A.; et al. Commodity risk assessment of Ficus carica plants from Israel. EFSA J. 2021, 19, 6353. [Google Scholar] [CrossRef]
- FAOSTAT. Production. Crops. 2020. Available online: http://www.fao.org/statistics/en/ (accessed on 20 December 2020).
- Biondi, A.; Guedes, R.N.C.; Wan, F.-H.; Desneux, N. Ecology, Worldwide Spread, and Management of the Invasive South American Tomato Pinworm, Tuta absoluta: Past, Present, and Future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Han, P.; Bayram, Y.; Shaltiel-Harpaz, L.; Sohrabi, F.; Saji, A.; Esenali, U.T.; Jalilov, A.; Ali, A.; Shashank, P.R.; Ismoilov, K.; et al. Tuta absoluta continues to disperse in Asia: Damage, ongoing management and future challenges. J. Pest Sci. 2019, 92, 1317–1327. [Google Scholar] [CrossRef]
- Tropea Garzia, G.; Siscaro, G.; Biondi, A.; Zappalà, L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Williams, D.; de Willink, M.G. Mealybugs of Central and South America; CABI: Wallingford, UK, 1992; 635p. [Google Scholar]
- Granara, M.C.; Szumik, C.A. Phenacoccinae de Centro y Sudamérica (Hemiptera: Coccoidea: Pseudococcidae): Sistemática y Filogenia. Rev. Soc. Entomol. Argent. 2007, 66, 29–129. [Google Scholar]
- Williams, D.J.; Watson, G.W. The Scale Insects of the Tropical South Pacific Region; CAB International Institute of Entomology: London, UK, 1988; 290p. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Cavalieri, V.; Mazzeo, G.; Garzia, G.T.; Buonocore, E.; Russo, A. Identification of Planococcus ficus and Planococcus citri (Hemiptera: Pseudococcidae) by PCR-RFLP of COI gene. Zootaxa 2008, 1816, 65–68. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Varlamov, A.; Vaskin, Y.; Efremov, I.; German Grehov, O.G.; Kandrov, D.; Rasputin, K.; Syabro, M.; et al. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J. Mealybugs of Southern Asia; The Natural History Museum Kuala Lumpur; Southdene SDN, BHD: Kuala Lumpur, Malaysia, 2004; p. 896. [Google Scholar]
- Mazzeo, G.; Russo, A.; Suma, P. Phenacoccus solani Ferris (Homoptera Coccoidea) on ornamental plants in Italy. Boll. Zool. Agrar. Bachic. 1999, 31, 31–35. [Google Scholar]
- Li, H.; Lang, K.L.; Fu, H.B.; Shen, C.P.; Wan, F.H.; Chu, D. Analysis of expressed sequence tags (ESTs) from a normalized cDNA library and isolation of EST simple sequence repeats from the invasive cotton mealybug Phenacoccus solenopsis. Insect Sci. 2015, 22, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Ao, Y.; Li, Z.; Wang, Y.; Jiang, M. Invasion biology of the cotton mealybug, Phenacoccus solenopsis Tinsley: Current knowledge and future directions. J. Integr. Agric. 2019, 18, 758–770. [Google Scholar] [CrossRef]
- Pellizzari, G.; Porcelli, F. First record of Phenacoccus defectus in Italy, with comments on Phenacoccus solani and Phenacoccus solenopsis. Bull. Insectology 2013, 66, 209–211. [Google Scholar]
- Sciortino, M.; Scurria, A.; Lino, C.; Pagliaro, M.; D’Agostino, F.; Tortorici, S.; Ricupero, M.; Biondi, A.; Zappalà, L.; Ciriminna, R. Silica-Microencapsulated Orange Oil for Sustainable Pest Control. Adv. Sustain. Syst. 2021, 5, 2000280. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Sites | Host Plant | Date | NCBI Accession Number | |
|---|---|---|---|---|---|
| Latitude (N) | Longitude (E) | ||||
| Marina di Ragusa | 36°47′11.7″ | 14°34′21.1″ | Lycopersicon esculentum Mill. (Solanaceae) Portulaca oleracea L. (Portulacaceae) Parietaria sp. L. (Urticaceae) Sesamum indicum L. (Pedaliaceae) | 22 October 2020 | MZ398130 |
| Palma di Montechiaro | 37°11′37″ | 13°45′46″ | Capsicum annuum L. (Solanaceae) | 5 October 2020 | MZ398131 |
| Catania | 37°31′18.0″ | 15°05′49.8″ | Hibiscus sp. L. (Malvaceae) | 4 Novenber 2020 | MZ398132 |
| 37°32′09.9″ | 15°04′06.1″ | Lantana camara L. (Verbenaceae) | 25 Novenber 2020 | MZ398133 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricupero, M.; Biondi, A.; Russo, A.; Zappalà, L.; Mazzeo, G. The Cotton Mealybug Is Spreading along the Mediterranean: First Pest Detection in Italian Tomatoes. Insects 2021, 12, 675. https://doi.org/10.3390/insects12080675
Ricupero M, Biondi A, Russo A, Zappalà L, Mazzeo G. The Cotton Mealybug Is Spreading along the Mediterranean: First Pest Detection in Italian Tomatoes. Insects. 2021; 12(8):675. https://doi.org/10.3390/insects12080675
Chicago/Turabian StyleRicupero, Michele, Antonio Biondi, Agatino Russo, Lucia Zappalà, and Gaetana Mazzeo. 2021. "The Cotton Mealybug Is Spreading along the Mediterranean: First Pest Detection in Italian Tomatoes" Insects 12, no. 8: 675. https://doi.org/10.3390/insects12080675
APA StyleRicupero, M., Biondi, A., Russo, A., Zappalà, L., & Mazzeo, G. (2021). The Cotton Mealybug Is Spreading along the Mediterranean: First Pest Detection in Italian Tomatoes. Insects, 12(8), 675. https://doi.org/10.3390/insects12080675