
Not Seeing the Mites for the Hairs. Comment on Möth et al. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180


1. Synopsis
2. Background
3. Methods
3.1. Characterization and Quantification of Microhabitat Quality
3.2. Statistical Analyses
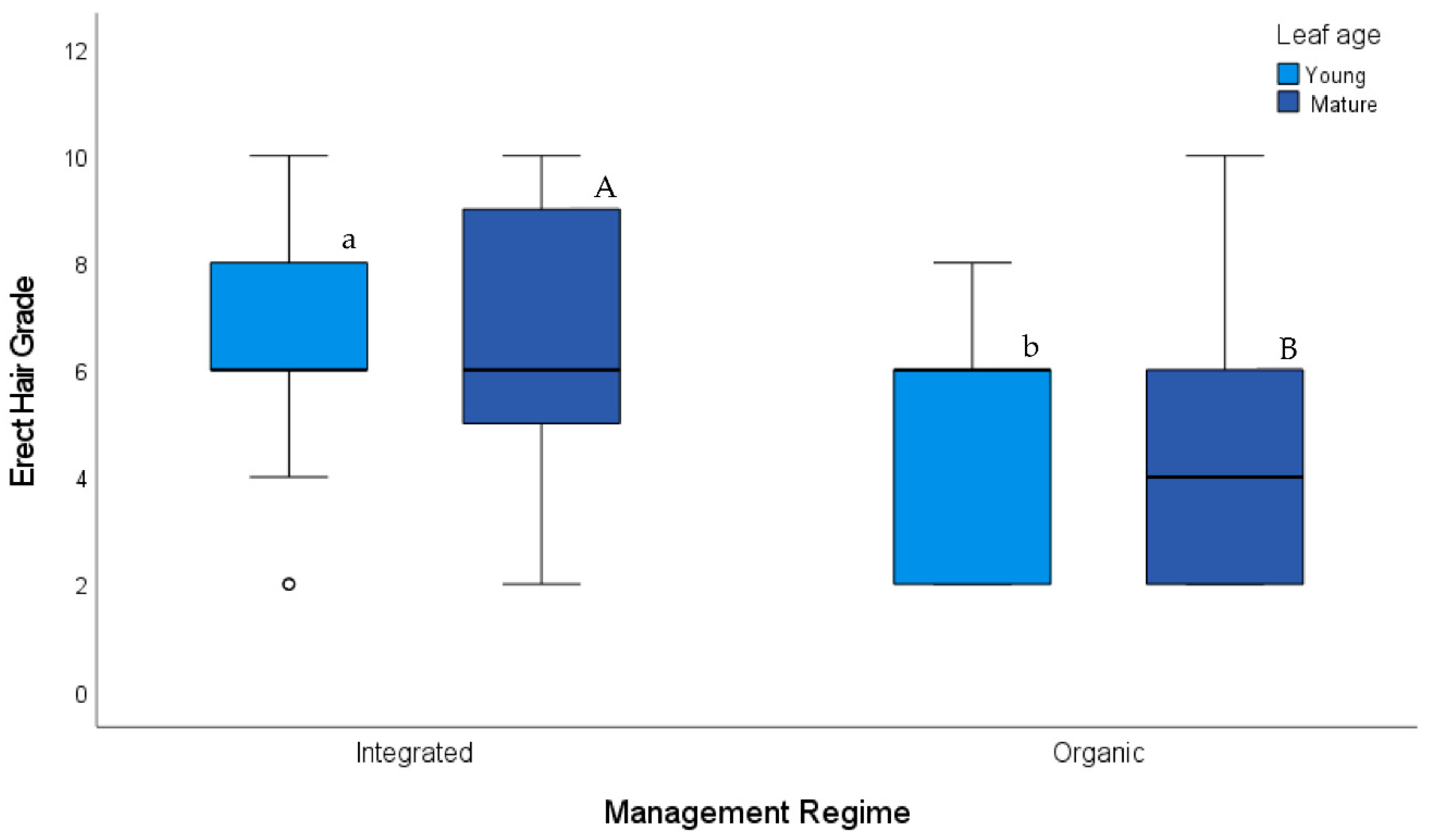
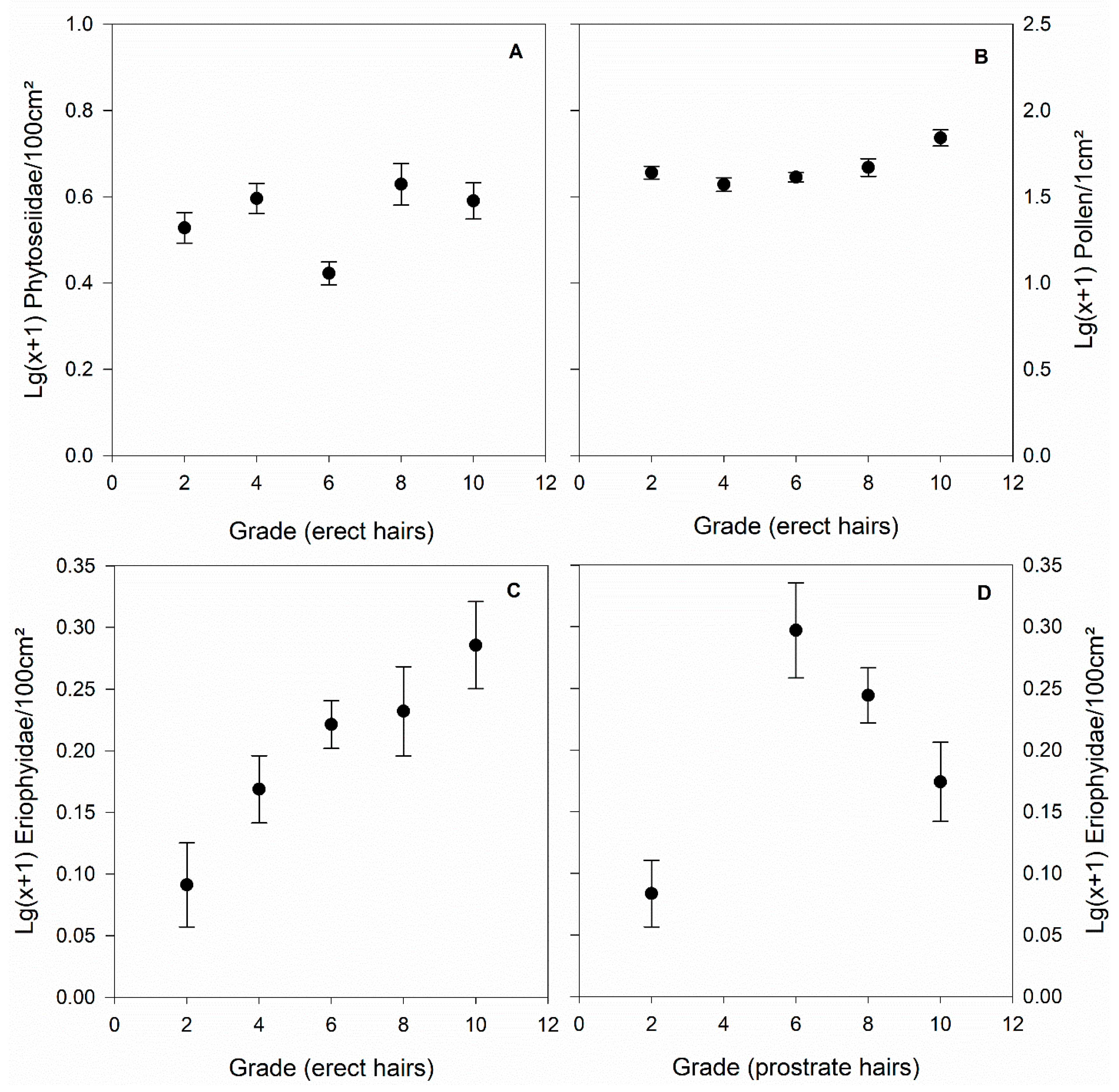
4. Results
5. Discussion
6. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Möth, S.; Walzer, A.; Redl, M.; Petrović, B.; Hoffmann, C.; Winter, S. Unexpected effects of local management and landscape composition on predatory mites and their food resources in vineyards. Insects 2021, 12, 180. [Google Scholar] [CrossRef]
- Price, P.W.; Bouton, C.E.; Gross, P.; McPheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions Among Three Trophic Levels: Influence of Plants on Interactions Between Insect Herbivores and Natural Enemies. Annu. Rev. Ecol. Syst. 1980, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.A. Leaf structures affect predatory mites (Acari: Phytoseiidae) and biological control: A review. Exp. Appl. Acarol. 2013, 62, 1–17. [Google Scholar] [CrossRef]
- Tixier, M.-S. Predatory Mites (Acari: Phytoseiidae) in Agro-Ecosystems and Conservation Biological Control: A Review and Explorative Approach for Forecasting Plant-Predatory Mite Interactions and Mite Dispersal. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009. [Google Scholar]
- Walter, D.E. Living on leaves: Mites, tomenta, and leaf domatia. Annu. Rev. Entomol. 1996, 41, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Tixier, M.-S.; Kreiter, S.; Auger, P. Colonization of vineyards by phytoseiid mites: Their dispersal patterns in the plot and their fate. Exp. Appl. Acarol. 2000, 24, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.A.; Stenseth, N.C.; Van Horne, B.; Ims, R.A. Ecological Mechanisms and Landscape Ecology. Oikos 1993, 66, 369–380. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. What size is a biologically relevant landscape? Landsc. Ecol. 2012, 27, 929–941. [Google Scholar] [CrossRef]
- Landsman, A.P.; Ladin, Z.S.; Gardner, D.; Bowman, J.L.; Shriver, G.; D’Amico, V.; Delaney, D.A. Local landscapes and micro-habitat characteristics are important determinants of urban–suburban forest bee communities. Ecosphere 2019, 10, e02908. [Google Scholar] [CrossRef] [Green Version]
- English-Loeb, G.; Norton, A.P.; Walker, M.A. Behavioral and population consequences of acarodomatia in grapes on phytoseiid mites (Mesostigmata) and implications for plant breeding. Entomol. Exp. Appl. 2002, 104, 307–319. [Google Scholar] [CrossRef]
- Loughner, R.; Goldman, K.; Loeb, G.; Nyrop, J. Influence of leaf trichomes on predatory mite (Typhlodromus pyri) abundance in grape varieties. Exp. Appl. Acarol. 2008, 45, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Duso, C. Role of Amblyseius aberrans (Oud.), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari, Phytoseiidae) in vineyards. III. Influence of variety characteristics on the success of A. aberrans and T. pyri releases. J. Appl. Entomol. 1992, 114, 455–462. [Google Scholar] [CrossRef]
- Duso, C.; Vettorazzo, E. Mite Population dynamics on different grape varieties with or without phytoseiids released (Acari: Phytoseiidae). Exp. Appl. Acarol. 1999, 23, 741–763. [Google Scholar] [CrossRef]
- Roda, A.; Nyrop, J.; English-Loeb, G. Leaf pubescence mediates the abundance of non-prey food and the density of the predatory mite Typhlodromus pyri. Exp. Appl. Acarol. 2003, 29, 193–211. [Google Scholar] [CrossRef]
- Loughner, R.; Wentworth, K.; Loeb, G.; Nyrop, J. Leaf trichomes influence predatory mite densities through dispersal behavior. Entomol. Exp. Appl. 2010, 134, 78–88. [Google Scholar] [CrossRef]
- Camporese, P.; Duso, C. Different colonization patterns of phytophagous and predatory mites (Acari: Tetranychidae, Phytoseiidae) on three grape varieties: A case study. Exp. Appl. Acarol. 1996, 20, 1–22. [Google Scholar]
- Tixier, M.-S.; Kreiter, S.; Auger, P.; Weber, M. Colonization of Languedoc vineyards by phytoseiid mites (Acari: Phytoseiidae): Influence of wind and crop environment. Exp. Appl. Acarol. 1998, 22, 523–542. [Google Scholar] [CrossRef]
- OIV-Organisation Internationale de la Vigne et du Vin. Description of World Wine Varieties. 2009. Available online: https://www.oiv.int/en/technical-standards-and-documents/description-of-grape-varieties/description-of-grape-varieties-throughout-the-world (accessed on 12 March 2021).
- HBLA und Bundesamt für Wein- und Obstbau Klosterneuburg. Rebsortenkatalog-Qualitätsweinsorten. 2021. Available online: http://www.weinobstklosterneuburg.at/service/rebsortenkatalog/qualitaetsweinrebsorten.html (accessed on 12 March 2021).
- Gerrath, J.; Posluszny, U.; Melville, L. Taming the Wild Grape: Botany and Horticulture in the Vitaceae; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Karban, R.; English-Loeb, G.; Walker, M.A.; Thaler, J. Abundance of phytoseiid mites on Vitis species: Effects of leaf hairs, domatia, prey abundance and plant phylogeny. Exp. Appl. Acarol. 1995, 19, 189–197. [Google Scholar] [CrossRef]
- Brewer, M.J.; Butler, A.; Cooksley, S.L. The relative performance of AIC, AICC and BIC in the presence of unobserved heterogeneity. Methods Ecol. Evol. 2016, 7, 679–692. [Google Scholar] [CrossRef]
- Konlechner, C.; Sauer, U. Ultrastructural leaf features of grapevine cultivars (Vitis vinifera L. ssp. vinifera). OENO One 2016, 50, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhu, H.; Ozkan, H.E.; Bagley, W.E.; Krause, C.R. Droplet evaporation and spread on waxy and hairy leaves associated with type and concentration of adjuvants. Pest Manag. Sci. 2011, 67, 842–851. [Google Scholar] [CrossRef]
- Gadino, A.N.; Walton, V.M.; Dreves, A.J. Impact of vineyard pesticides on a beneficial arthropod, Typhlodromus pyri (Acari: Phytoseiidae), in laboratory bioassays. J. Econ. Èntomol. 2011, 104, 970–977. [Google Scholar] [CrossRef]
- Hanna, R.; Zalom, F.G.; Wilson, L.T.; Leavitt, G.M. Sulfur can suppress mite predators in vineyards. Calif. Agric. 1997, 51, 19–21. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Dependent and Explanatory Variables | Wald ꭓ2 | df | p Value |
|---|---|---|---|
| Phytoseiidae | |||
| (Intercept) | 52.344 | 1 | <0.001 |
| Date | 77.757 | 4 | <0.001 |
| Pest Management Regime | 19.055 | 1 | <0.001 |
| Interrow Vegetation Type | 8.618 | 2 | 0.013 |
| Erect Hair Grade | 26.979 | 4 | <0.001 |
| Percent Vineyards | 22.815 | 1 | <0.001 |
| Eriophyidae | |||
| (Intercept) | 0.858 | 1 | 0.354 |
| Date | 81.995 | 4 | <0.001 |
| Interrow Vegetation Type | 7.229 | 2 | 0.027 |
| Prostrate Hair Grade | 21.346 | 3 | <0.001 |
| Erect Hair Grade | 15.254 | 4 | 0.004 |
| Pollen | 3.858 | 1 | 0.050 |
| Tydeidae | |||
| (Intercept) | 184.144 | 1 | <0.001 |
| Date | 138.225 | 4 | <0.001 |
| Interrow Vegetation Type | 25.281 | 2 | <0.001 |
| Pollen | |||
| (Intercept) | 8420.283 | 1 | <0.001 |
| Date | 319.278 | 4 | <0.001 |
| Erect Hair Grade | 23.057 | 4 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schausberger, P. Not Seeing the Mites for the Hairs. Comment on Möth et al. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180. Insects 2021, 12, 671. https://doi.org/10.3390/insects12080671
Schausberger P. Not Seeing the Mites for the Hairs. Comment on Möth et al. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180. Insects. 2021; 12(8):671. https://doi.org/10.3390/insects12080671
Chicago/Turabian StyleSchausberger, Peter. 2021. "Not Seeing the Mites for the Hairs. Comment on Möth et al. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180" Insects 12, no. 8: 671. https://doi.org/10.3390/insects12080671
APA StyleSchausberger, P. (2021). Not Seeing the Mites for the Hairs. Comment on Möth et al. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180. Insects, 12(8), 671. https://doi.org/10.3390/insects12080671