Active Human and Murine Tumor Necrosis Factor α Cytokines Produced from Silkworm Baculovirus Expression System


 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Silkworms
2.2. Construction of Recombinant Baculoviruses
2.3. Expression and Purification of rhTNFα and rmTNFα in Silkworm Larvae
2.4. Removal of Terminal Fusion Tags from Purified rh/mTNFα Proteins
2.5. Bioassay of rTNFα Activity
3. Results and Discussion
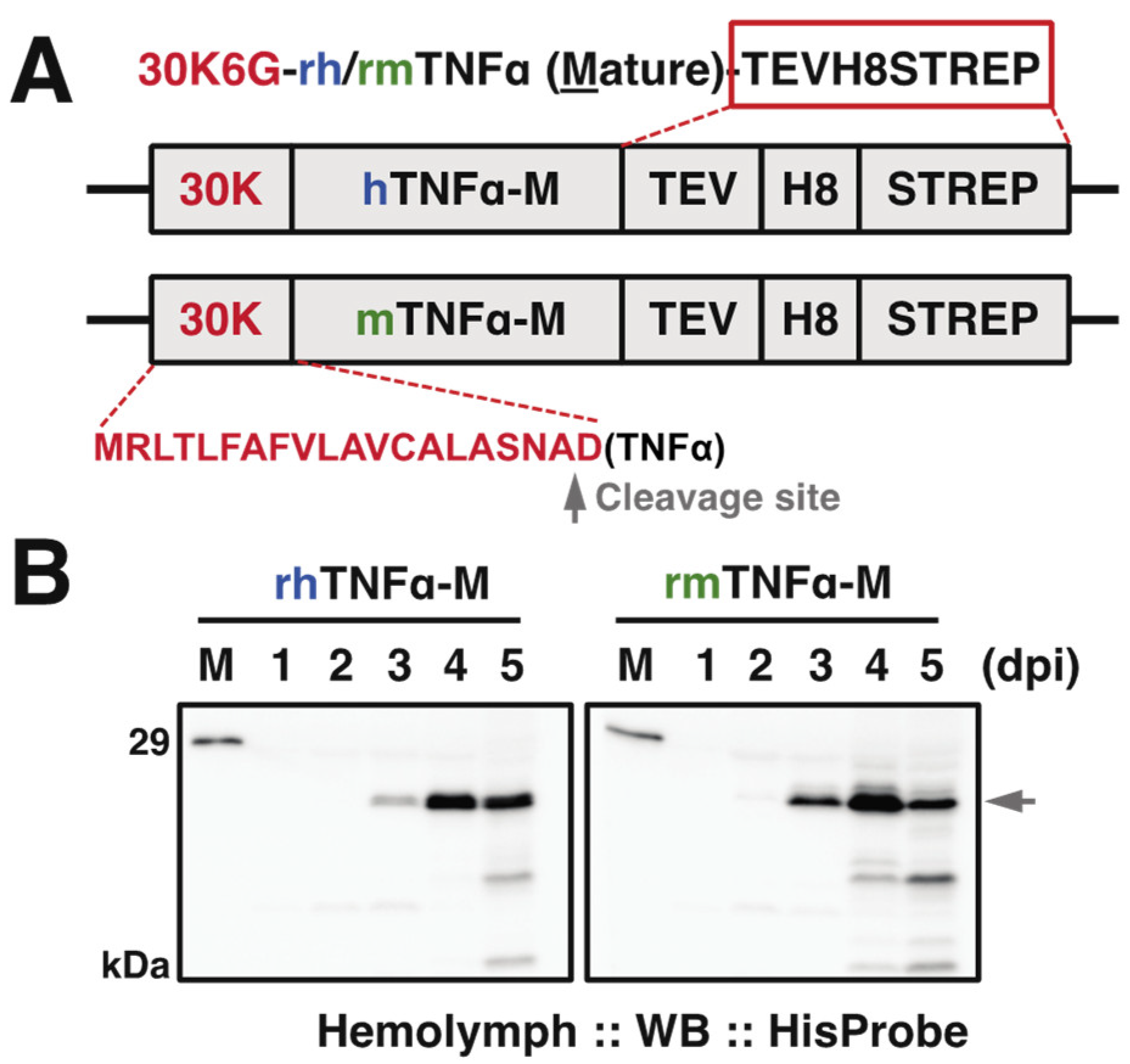
3.1. Construction of rBmNPVs for the Production of rh/rmTNFα
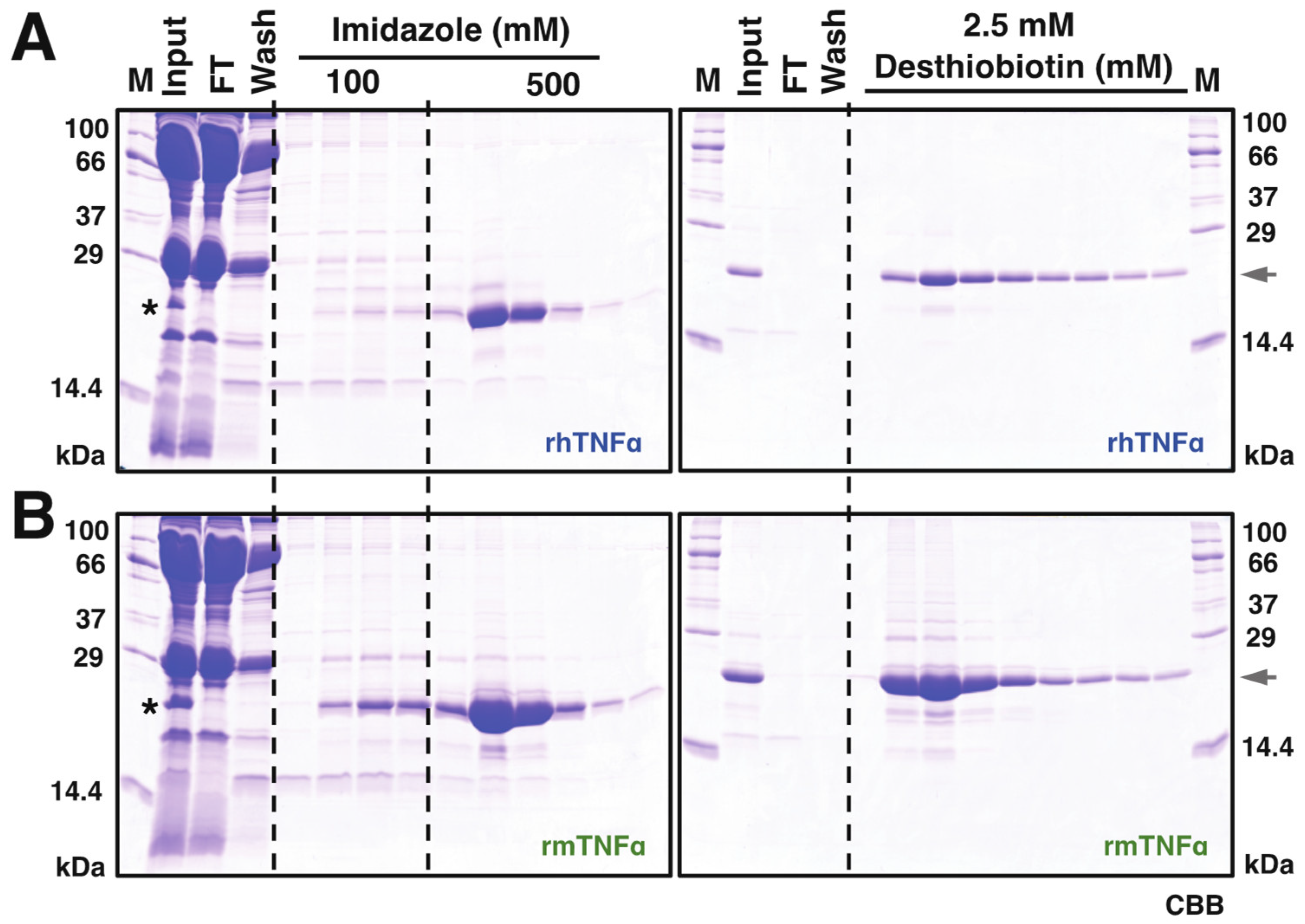
3.2. Purification of rh/rmTNFα from Silkworm Serum
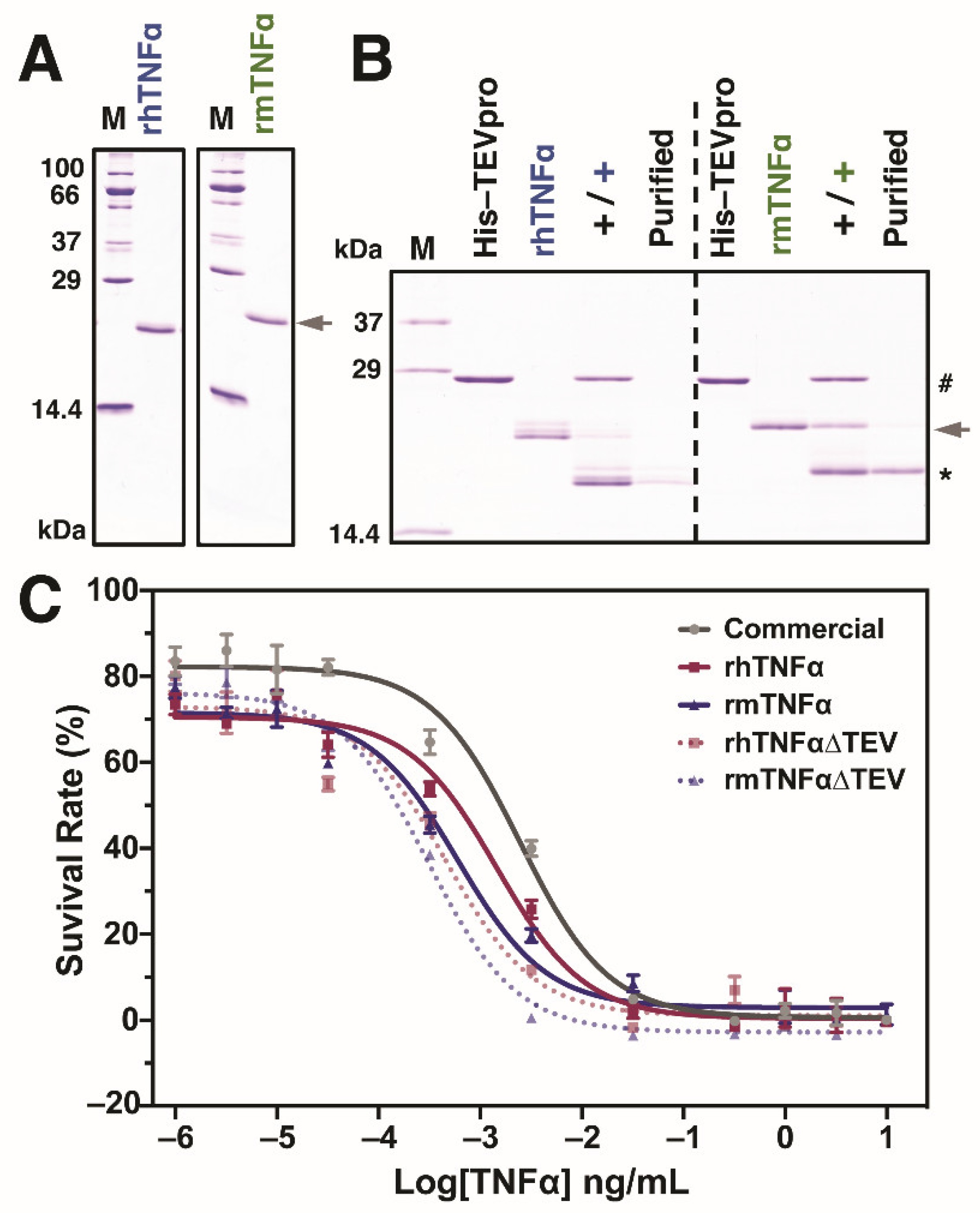
3.3. Biological Activity
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tracey, K.J.; Cerami, A. Tumor necrosis factor: A pleiotropic cytokine and therapeutic target. Annu. Rev. Med. 1994, 45, 491–503. [Google Scholar] [CrossRef]
- Brenner, D.; Blaser, H.; Mak, T.W. Regulation of tumour necrosis factor signalling: Live or let die. Nat. Rev. Immunol. 2015, 15, 362–374. [Google Scholar] [CrossRef]
- Kalliolias, G.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Rohde, F.; Schusser, B.; Hron, T.; Farkašová, H.; Plachý, J.; Härtle, S.; Hejnar, J.; Elleder, D.; Kaspers, B. Characterization of Chicken Tumor Necrosis Factor-α, a Long Missed Cytokine in Birds. Front. Immunol. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Black, R.A.; Rauch, C.T.; Kozlosky, C.J.; Peschon, J.J.; Slack, J.L.; Wolfson, M.F.; Castner, B.J.; Stocking, K.L.; Reddy, P.; Srinivasan, S.; et al. A metalloproteinase disintegrin that releases tumour-necrosis factor-α from cells. Nature 1997, 385, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Van Hauwermeiren, F.; Vandenbroucke, R.; Libert, C. Treatment of TNF mediated diseases by selective inhibition of soluble TNF or TNFR. Cytokine Growth Factor Rev. 2011, 22, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Damough, S.; Sabzalinezhad, M.; Talebkhan, Y.; Nematollahi, L.; Bayat, E.; Torkashvand, F.; Adeli, A.; Jahandar, H.; Barkhordari, F.; Mahboudi, F. Optimization of culture conditions for high-level expression of soluble and active tumor necrosis factor-α in E. coli. Protein Expr. Purif. 2021, 179, 105805. [Google Scholar] [CrossRef]
- Singha, T.K.; Dagar, V.K.; Gulati, P.; Kumar, S. Kinetic study and optimization of recombinant human tumor necrosis factor-alpha (rhTNF-α) production in Escherichia coli. Prep. Biochem. Biotechnol. 2020, 51, 267–276. [Google Scholar] [CrossRef]
- Pozidis, C.; Lammertyn, E.; Politou, A.S.; Anné, J.; Tsiftsoglou, A.S.; Sianidis, G.; Economou, A. Protein secretion biotechnology using Streptomyces lividans: Large-scale production of functional trimeric tumor necrosis factor alpha. Biotechnol. Bioeng. 2001, 72, 611–619. [Google Scholar] [CrossRef]
- Zhang, X.-W.; Sun, T.; Zeng, X.-Y.; Liu, X.; Gu, D.-X. Expression of recombinant human tumor necrosis factor-α in a baculovirus expression system. Bioresour. Technol. 1999, 70, 299–301. [Google Scholar] [CrossRef]
- Wang, F.; Zeng, L.; Wang, Y.-L.; Cui, S.-Q.; Hu, L.; Zheng, J.-M.; Huang, D.-N.; Hou, G. Construction and characterization of a transmembrane eukaryotic expression vector based on the membrane domain structure of TNF-α. Mol. Med. Rep. 2017, 16, 1021–1030. [Google Scholar] [CrossRef][Green Version]
- Castiñeiras, T.S.; Williams, S.G.; Hitchcock, A.; Cole, J.A.; Smith, D.C.; Overton, T.W. Optimizing host cell physiology and stress avoidance for the production of recombinant human tumour necrosis factor α in Escherichia coli. Microbiology 2018, 164, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Hamzeh-Mivehroud, M.; Farajzadeh, M.; Moosavi-Movahedi, A.; Dastmalchi, S. A simple and rapid method for expression and purification of functional TNF-α using GST fusion system. Curr. Pharm. Biotechnol. 2015, 16, 707–715. [Google Scholar] [CrossRef]
- Han, W.; Zhang, Y.; Yan, Z.; Shi, J. Construction of a new tumour necrosis factor fusion-protein expression vector for high-level expression of heterologous genes in Escherichia coli. Biotechnol. Appl. Biochem. 2003, 37, 109–113. [Google Scholar] [CrossRef]
- Tripathi, N.; Shrivastava, A. Recent developments in bioprocessing of recombinant proteins: Expression hosts and process development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef]
- Jia, B.; Jeon, C.O. High-throughput recombinant protein expression in Escherichia coli: Current status and future perspectives. Open Biol. 2016, 6, 6. [Google Scholar] [CrossRef]
- Kato, T.; Kajikawa, M.; Maenaka, K.; Park, E.Y. Silkworm expression system as a platform technology in life science. Appl. Microbiol. Biotechnol. 2009, 85, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus–insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Bieniossek, C.; Richmond, T.J.; Berger, I. MultiBac: Multigene baculovirus-based eukaryotic protein complex production. Curr. Protoc. Protein Sci. 2008, 51, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Berger, I.; Fitzgerald, D.J.; Richmond, T.J. Baculovirus expression system for heterologous multiprotein complexes. Nat. Biotechnol. 2004, 22, 1583–1587. [Google Scholar] [CrossRef]
- Kakino, K.; Masuda, A.; Hino, M.; Ebihara, T.; Xu, J.; Mon, H.; Fujita, R.; Fujii, T.; Kusakabe, T.; Lee, J.M. Efficient production of recombinant T7 endonuclease I using silkworm-baculovirus expression vector system. J. Asia-Pac. Èntomol. 2020, 23, 694–700. [Google Scholar] [CrossRef]
- Kobayashi, M.; Xu, J.; Kakino, K.; Masuda, A.; Hino, M.; Fujimoto, N.; Minamihata, K.; Kamiya, N.; Mon, H.; Iida, H.; et al. Optimal silkworm larva host for high-level production of Mus musculus IL-4 using a baculovirus expression vector system. J. Asia-Pac. Èntomol. 2020, 23, 268–273. [Google Scholar] [CrossRef]
- Fujita, R.; Hino, M.; Ebihara, T.; Nagasato, T.; Masuda, A.; Lee, J.M.; Fujii, T.; Mon, H.; Kakino, K.; Nagai, R.; et al. Efficient production of recombinant SARS-CoV-2 spike protein using the baculovirus-silkworm system. Biochem. Biophys. Res. Commun. 2020, 529, 257–262. [Google Scholar] [CrossRef]
- Sano, K.-I.; Maeda, K.; Oki, M.; Maéda, Y. Enhancement of protein expression in insect cells by a lobster tropomyosin cDNA leader sequence. FEBS Lett. 2002, 532, 143–146. [Google Scholar] [CrossRef]
- Mohanty, A.K.; Wiener, M.C. Membrane protein expression and production: Effects of polyhistidine tag length and position. Protein Expr. Purif. 2004, 33, 311–325. [Google Scholar] [CrossRef]
- Soejima, Y.; Lee, J.; Nagata, Y.; Mon, H.; Iiyama, K.; Kitano, H.; Matsuyama, M.; Kusakabe, T. Comparison of signal peptides for efficient protein secretion in the baculovirus-silkworm system. Open Life Sci. 2013, 8, 1–7. [Google Scholar] [CrossRef]
- Ono, C.; Nakatsukasa, T.; Nishijima, Y. Construction of the BmNPV T3 bacmid system and its application to the functional analysis of BmNPV he65. J. Insect Biotechnol. Sericol. 2007, 76, 3_161–3_167. [Google Scholar]
- Morifuji, Y.; Xu, J.; Karasaki, N.; Iiyama, K.; Morokuma, D.; Hino, M.; Masuda, A.; Yano, T.; Mon, H.; Kusakabe, T.; et al. Expression, Purification, and characterization of recombinant human α1-antitrypsin produced using silkworm-baculovirus expression system. Mol. Biotechnol. 2018, 60, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Osthoff, K.; Bakker, A.; Vanhaesebroeck, B.; Beyaert, R.; Jacob, W.; Fiers, W. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 1992, 267, 5317–5323. [Google Scholar] [CrossRef]
- Amer, Y.A.; Erdmann, J.; Alexopoulou, L.; Kollias, G.; Ross, F.P.; Teitelbaum, S.L. Tumor necrosis factor receptors types 1 and 2 differentially regulate osteoclastogenesis. J. Biol. Chem. 2000, 275, 27307–27310. [Google Scholar] [CrossRef]
- Scallon, B.J.; Trinh, H.; Nedelman, M.; Brennan, F.M.; Feldmann, M.; Ghrayeb, J. Functional comparisons of different tumour necrosis factor receptor/IgG fusion proteins. Cytokine 1995, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Fonda, I.; Kenig, M.; Gaberc-Porekar, V.; Pristovaek, P.; Menart, V. Attachment of histidine tags to recombinant tumor necrosis factor-alpha drastically changes its properties. Sci. World J. 2002, 2, 1312–1325. [Google Scholar] [CrossRef]
- Kawakami, N.; Lee, J.M.; Mon, H.; Kubo, Y.; Banno, Y.; Kawaguchi, Y.; Maenaka, K.; Park, E.Y.; Koga, K.; Kusakabe, T. Efficient protein expression in Bombyx mori larvae of the strain d17 highly sensitive to B. mori nucleopolyhedrovirus. Mol. Biotechnol. 2008, 40, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Xu, J.; Mitsudome, T.; Morokuma, D.; Mon, H.; Banno, Y.; Kusakabe, T.; Lee, J.M. Improvement of Endo-β-N-acetylglucosaminidase H production using silkworm–baculovirus protein expression system. J. Asia-Pac. Èntomol. 2015, 18, 175–180. [Google Scholar] [CrossRef]
- Jin, H.; Uddin, M.S.; Huang, Y.L.; Teo, W.K. Purification and renaturation of recombinant human lymphotoxin (tumour necrosis factor beta) expressed in Escherichia coli as inclusion bodies. J. Chem. Technol. Biotechnol. 1994, 59, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, W.; Gao, D.; Wang, L.; Yang, Y.; Bai, Q. One-step refolding and purification of recombinant human tumor necrosis factor-α (rhTNF-α) using ion-exchange chromatography. Biomed. Chromatogr. 2015, 29, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Kawai, T.; Obinata, M.; Fujiwara, H.; Horiuchi, T.; Saeki, Y.; Sato, Y.; Furusawa, M. Production of human α-interferon in silkworm using a baculovirus vector. Nat. Cell Biol. 1985, 315, 592–594. [Google Scholar] [CrossRef]
- Xu, J.; Lee, J.M.; Tatsuke, T.; Ebihara, T.; Masuda, A.; Hino, M.; Morokuma, D.; Fujita, R.; Mon, H.; Kusakabe, T.; et al. Expression and purification of vaccinia virus DNA topoisomerase IB produced in the silkworm–baculovirus expression system. Mol. Biotechnol. 2019, 61, 622–630. [Google Scholar] [CrossRef]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef]
- Sugarman, B.J.; Aggarwal, B.B.; Hass, P.E.; Figari, I.S.; Palladino, M.A.; Shepard, H.M. Recombinant human tumor necrosis factor-alpha: Effects on proliferation of normal and transformed cells in vitro. Science 1985, 230, 943–945. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Y.; Zhao, D.; Li, X.; Yu, R.; Su, Z. Facile purification of Escherichia coli expressed tag-free recombinant human tumor necrosis factor alpha from supernatant. Protein Expr. Purif. 2014, 95, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Kosobokova, E.N.; Skrypnik, K.A.; Kosorukov, V.S. Overview of fusion tags for recombinant proteins. Biochemistry 2016, 81, 187–200. [Google Scholar] [CrossRef]
- Hoffmann, A.; Müller, M.Q.; Gloser, M.; Sinz, A.; Rudolph, R.; Pfeifer, S. Recombinant production of bioactive human TNF-α by SUMO-fusion system—High yields from shake-flask culture. Protein Expr. Purif. 2010, 72, 238–243. [Google Scholar] [CrossRef]
- Dai, Y.-C.; Yang, S.-M.; Wang, X.; Zhou, Y.-J.; Hou, G.; Huang, D.-N. Antitumor effect and mechanism of action of a tumor-targeting recombinant human tumor necrosis factor-α fusion protein mediated by urokinase. Mol. Med. Rep. 2015, 11, 4333–4340. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Lee, J.M.; Miyata, T.; Sato, T.; Hayashi, S.; Hino, M.; Morokuma, D.; Karasaki, N.; Mon, H.; Kusakabe, T. Purification and characterization of immunogenic recombinant virus-like particles of porcine circovirus type 2 expressed in silkworm pupae. J. Gen. Virol. 2018, 99, 917–926. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebihara, T.; Xu, J.; Tonooka, Y.; Nagasato, T.; Kakino, K.; Masuda, A.; Minamihata, K.; Kamiya, N.; Nakatake, H.; Chieda, Y.; et al. Active Human and Murine Tumor Necrosis Factor α Cytokines Produced from Silkworm Baculovirus Expression System. Insects 2021, 12, 517. https://doi.org/10.3390/insects12060517
Ebihara T, Xu J, Tonooka Y, Nagasato T, Kakino K, Masuda A, Minamihata K, Kamiya N, Nakatake H, Chieda Y, et al. Active Human and Murine Tumor Necrosis Factor α Cytokines Produced from Silkworm Baculovirus Expression System. Insects. 2021; 12(6):517. https://doi.org/10.3390/insects12060517
Chicago/Turabian StyleEbihara, Takeru, Jian Xu, Yoshino Tonooka, Takumi Nagasato, Kohei Kakino, Akitsu Masuda, Kosuke Minamihata, Noriho Kamiya, Hirokazu Nakatake, Yuuka Chieda, and et al. 2021. "Active Human and Murine Tumor Necrosis Factor α Cytokines Produced from Silkworm Baculovirus Expression System" Insects 12, no. 6: 517. https://doi.org/10.3390/insects12060517
APA StyleEbihara, T., Xu, J., Tonooka, Y., Nagasato, T., Kakino, K., Masuda, A., Minamihata, K., Kamiya, N., Nakatake, H., Chieda, Y., Mon, H., Fujii, T., Kusakabe, T., & Lee, J. M. (2021). Active Human and Murine Tumor Necrosis Factor α Cytokines Produced from Silkworm Baculovirus Expression System. Insects, 12(6), 517. https://doi.org/10.3390/insects12060517