
Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i


Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
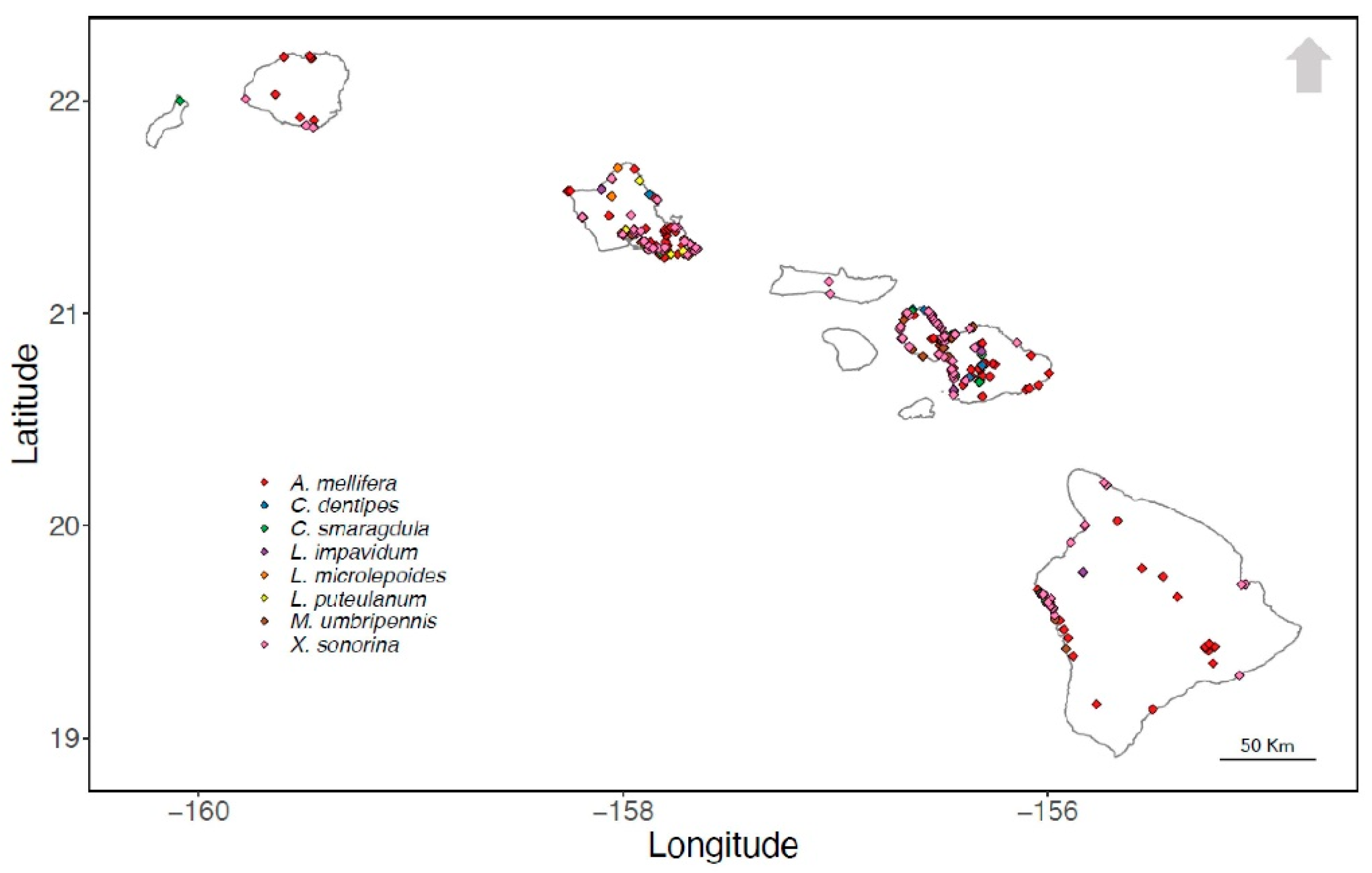
2.1. Species Occurrence Data
2.2. Environmental Data
2.3. Species Distribution Modeling
2.4. Bioclimatic Niche Analysis
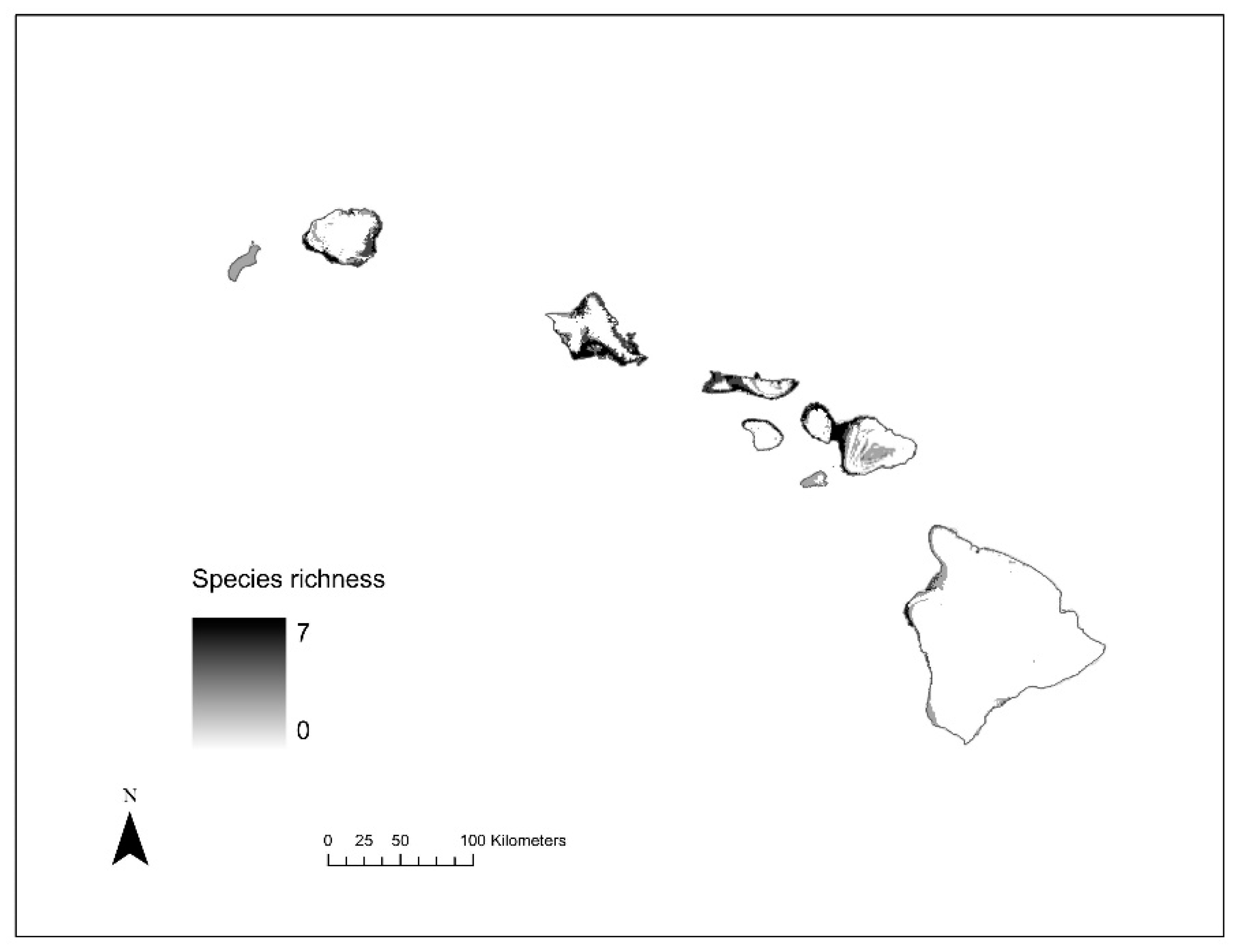
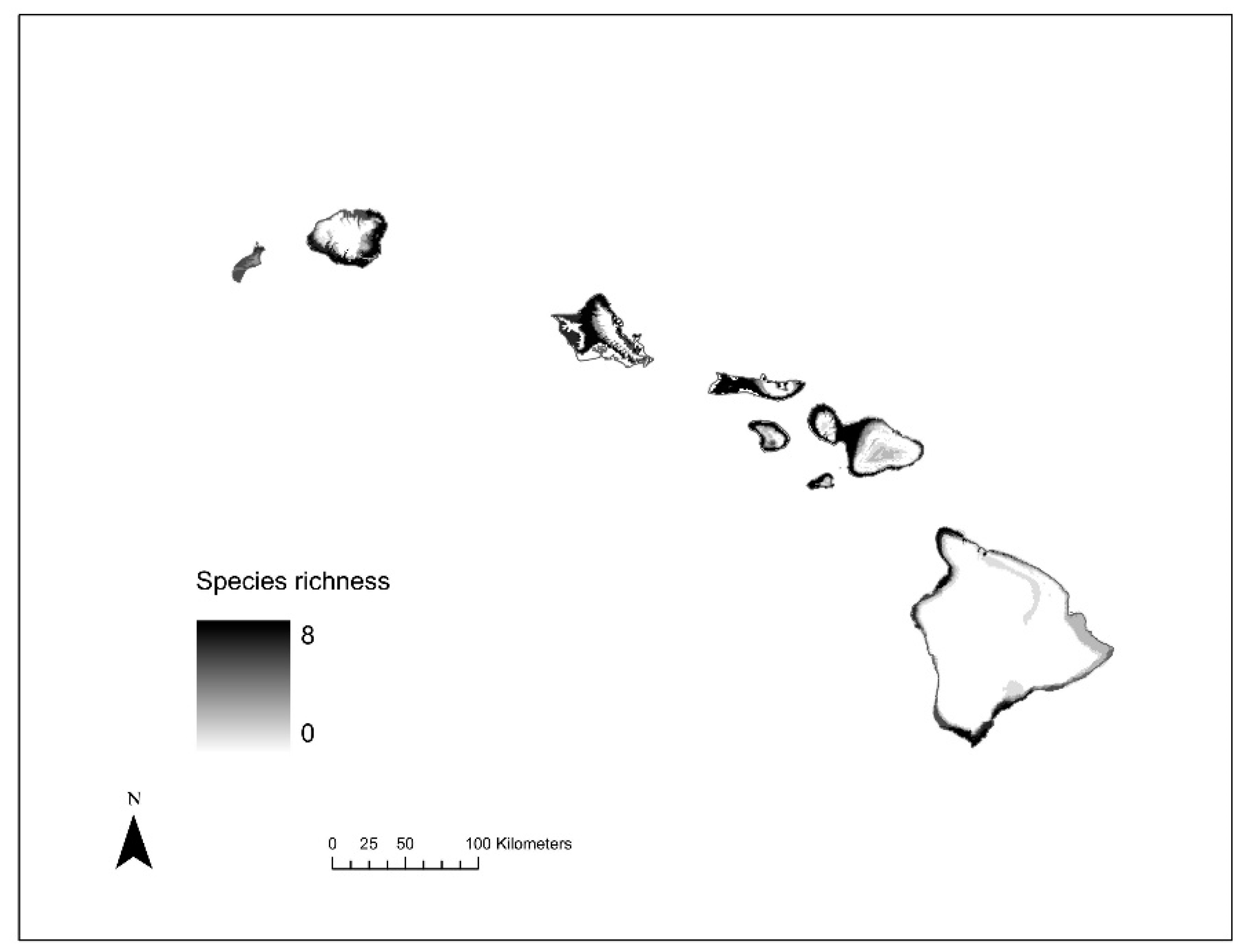
2.5. Species Richness Analysis
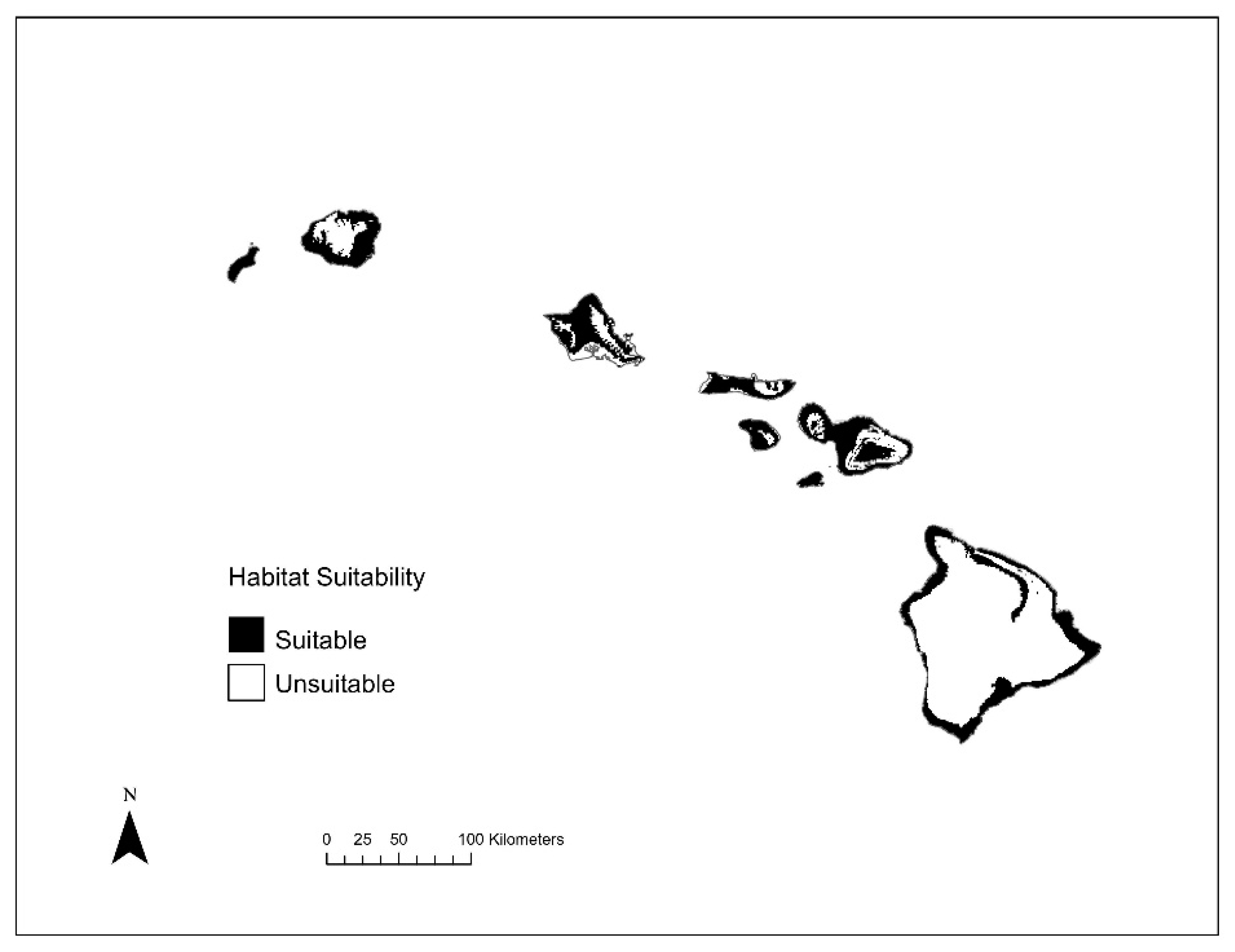
3. Results
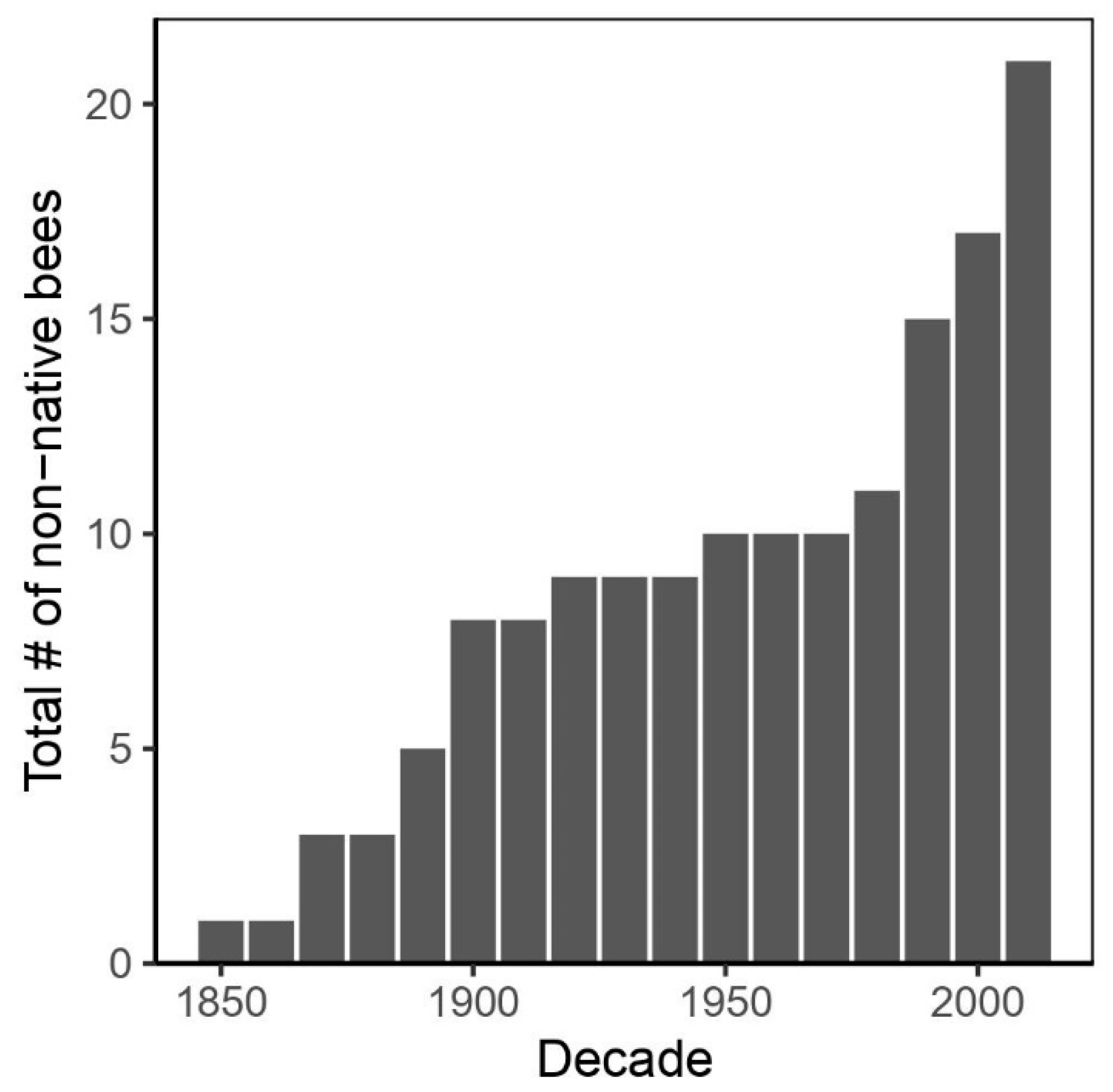
3.1. Species Occurrence Data
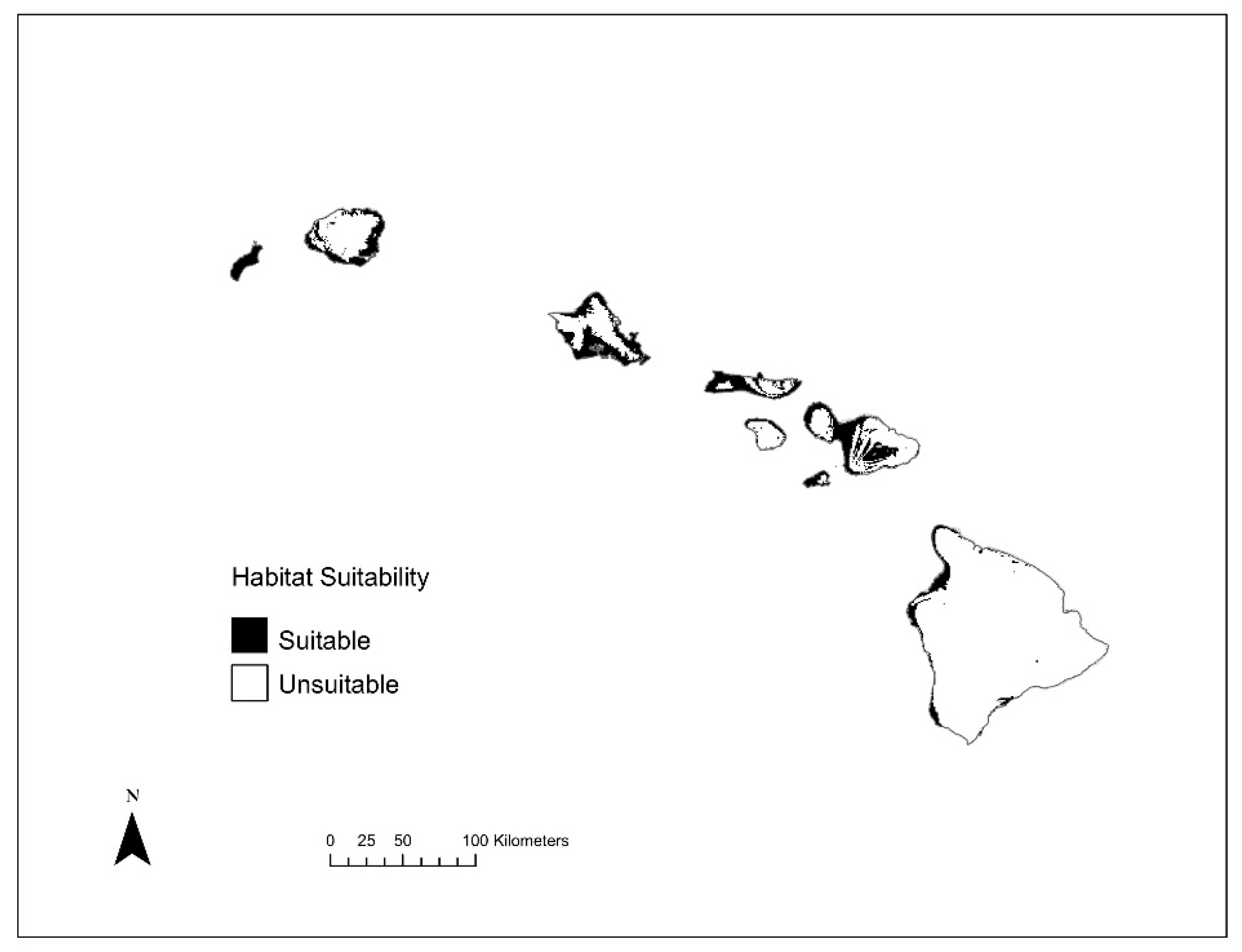
3.2. Species Distribution Modeling
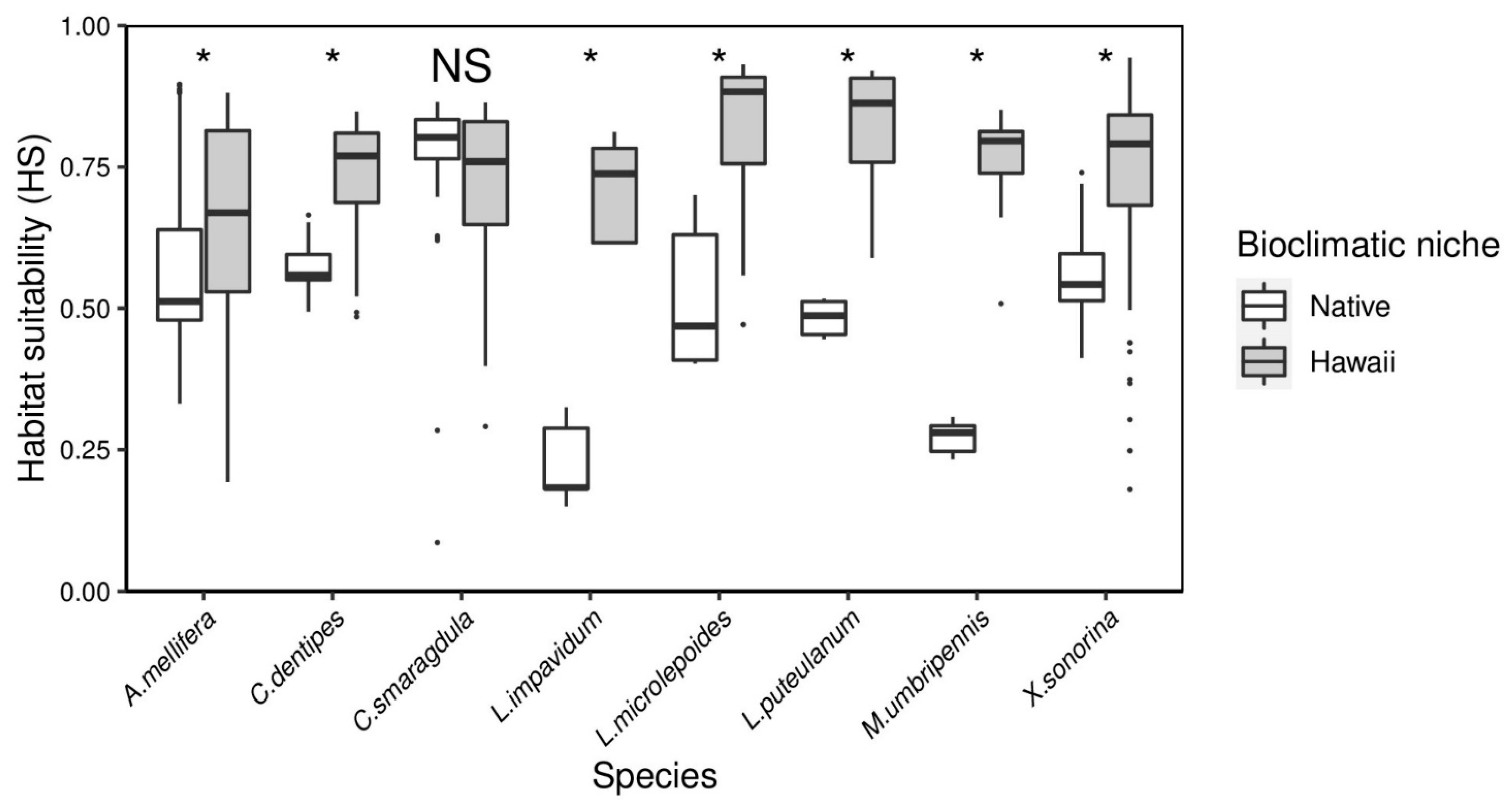
3.3. Bioclimatic Niche Analysis
3.4. Species Richness Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dew, R.M.; Silva, D.P.; Rehan, S.M. Range expansion of an already widespread bee under climate change. Glob. Ecol. Conserv. 2019, 17, e00584. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; Vanbergen, A.J. The Assessment Report on Pollinators, Pollination and Food Production: Summary for Policymakers; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016; p. 36. ISBN 9789280735680. [Google Scholar]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef]
- Bellard, C.; Thuiller, W.; Leroy, B.; Genovesi, P.; Bakkenes, M.; Courchamp, F. Will climate change promote future invasions? Glob. Chang. Biol. 2013, 19, 3740–3748. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Bartomeus, I.; Ascher, J.S.; Wagner, D.; Danforth, B.N.; Colla, S.; Kornbluth, S.; Winfree, R. Climate-associated phenological advances in bee pollinators and bee-pollinated plants. Proc. Natl. Acad. Sci. USA 2011, 108, 20645–20649. [Google Scholar] [CrossRef]
- Rubidge, E.M.; Patton, J.L.; Lim, M.; Cole Burton, A.; Brashares, J.S.; Moritz, C. Climate-induced range contraction drives genetic erosion in an alpine mammal. Nat. Clim. Chang. 2012, 2, 285–288. [Google Scholar] [CrossRef]
- Magnacca, K.N.; Gibbs, J.; Droege, S. Notes on alien and native bees (Hymenoptera: Apoidea) from the Hawaiian Islands. Bish. Mus. Occas. Pap. 2013, 114, 61–65. [Google Scholar]
- Magnacca, K.N. Conservation status of the endemic bees of Hawai’i, Hylaeus (Nesoprosopis) (Hymenoptera: Colletidae). Pac. Sci. 2007, 61, 173–190. [Google Scholar] [CrossRef]
- Snelling, R.R. Bees of the Hawaiian Islands, exclusive of Hylaeus (Nesoprosopis) (Hymenoptera: Apoidea). J. Kans. Entomol. Soc. 2003, 76, 342–356. [Google Scholar]
- Reaser, J.K.; Meyerson, L.A.; Cronk, Q.; de Poorter, M.; Eldrege, L.G.; Green, E.; Kairo, M.; Latasi, P.; Mack, R.N.; Mauremootoo, J.; et al. Ecological and socioeconomic impacts of invasive alien species in island ecosystems. Environ. Conserv. 2007, 34, 98–111. [Google Scholar] [CrossRef]
- Barrows, E.M. Robbing of Exotic Plants by Introduced Carpenter and Honey Bees in Hawaii, with Comparative Notes. Biotropica 1980, 12, 23–29. [Google Scholar] [CrossRef]
- Roddy, K.M.; Arita-Tsutsumi, L. A history of honey bees in the Hawaiian Islands. J. Hawaii Pac. Agric. 1997, 8, 59–70. [Google Scholar]
- Shell, W.A.; Rehan, S.M. Invasive range expansion of the small carpenter bee, Ceratina dentipes (Hymenoptera: Apidae) into Hawai’i with implications for native endangered species displacement. Biol. Invasions 2019, 21, 1155–1166. [Google Scholar] [CrossRef]
- Shell, W.A.; Rehan, S.M. Range Expansion of the Small Carpenter Bee Ceratina smaragdula across the Hawaiian Archipelago with Potential Ecological Implications for Native Pollinator Systems. Pac. Sci. 2017, 71, 1–15. [Google Scholar] [CrossRef]
- Stout, J.C.; Morales, C.L. Ecological impacts of invasive alien species on bees. Apidologie 2009, 40, 388–409. [Google Scholar] [CrossRef]
- Poulsen, N.R.; Rasmussen, C. Island bees: Do wood nesting bees have better island dispersal abilities? Apidologie 2020, 51, 1006–1017. [Google Scholar] [CrossRef]
- Groom, S.V.C.; Hayes, S.E.; Ngo, H.T.; Stevens, M.I.; Schwarz, M.P. Recipe for disruption: Multiple recent arrivals of megachilid bees in Pacific archipelagos. J. Insect Conserv. 2014, 18, 613–622. [Google Scholar] [CrossRef]
- Rasmussen, C.; Carríon, A.L.; Castro-Urgal, R.; Chamorro, S.; Gonzalez, V.H.; Griswold, T.L.; Herrera, H.W.; McMullen, C.K.; Olesen, J.M.; Traveset, A. Megachile timberlakei Cockerell (Hymenoptera: Megachilidae): Yet another adventive bee species to the Galápagos Archipelago. Pan-Pac. Entomol. 2012, 88, 98–102. [Google Scholar] [CrossRef]
- Magnacca, K.N.; King, C. Assessing the presence and distribution of 23 Hawaiian yellow-faced bee species on lands adjacent to military installations on O’ahu and Hawai’i Island. Tech. Rep. 2013, 185, 26. [Google Scholar]
- Groom, S.V.C.; Stevens, M.I.; Ramage, T.; Schwarz, M.P. Origins and implications of apid bees (Hymentopera: Apidae) in French Polynesia. Entomol. Sci. 2017, 20, 65–75. [Google Scholar] [CrossRef]
- Manson, D.C.M. The occurrence of the carpenter bee, Xylocopa sonorina in New Zealand. Weta 1988, 11, 4–5. [Google Scholar]
- Imbach, P.; Fung, E.; Hannah, L.; Navarro-Racines, C.E.; Roubik, D.W.; Ricketts, T.H.; Harvey, C.A.; Donatti, C.I.; Läderach, P.; Locatelli, B.; et al. Coupling of pollination services and coffee suitability under climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 10438–10442. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech. 2008, 27, 485–497, 499–510. [Google Scholar] [CrossRef]
- Giannini, T.C.; Maia-Silva, C.; Acosta, A.L.; Jaffé, R.; Carvalho, A.T.; Martins, C.F.; Zanella, F.C.V.; Carvalho, C.A.L.; Hrncir, M.; Saraiva, A.M.; et al. Protecting a managed bee pollinator against climate change: Strategies for an area with extreme climatic conditions and socioeconomic vulnerability. Apidologie 2017, 48, 784–794. [Google Scholar] [CrossRef]
- Giannini, T.C.; Acosta, A.L.; Garófalo, C.A.; Saraiva, A.M.; Alves-dos-Santos, I.; Imperatriz-Fonseca, V.L. Pollination services at risk: Bee habitats will decrease owing to climate change in Brazil. Ecol. Model. 2012, 244, 127–131. [Google Scholar] [CrossRef]
- Giannini, T.C.; Costa, W.F.; Borges, R.C.; Miranda, L.; da Costa, C.P.W.; Saraiva, A.M.; Imperatriz Fonseca, V.L. Climate change in the Eastern Amazon: Crop-pollinator and occurrence-restricted bees are potentially more affected. Reg. Environ. Chang. 2020, 20, 9. [Google Scholar] [CrossRef]
- Taylor, S.; Kumar, L. Global Climate Change Impacts on Pacific Islands Terrestrial Biodiversity: A Review. Trop. Conserv. Sci. 2016, 9, 203–223. [Google Scholar] [CrossRef]
- Magnacca, K.N. Two new species of Hylaeus (Nesoprosopis) (Hymenoptera: Colletidae) from O’ahu, Hawai’i. Zootaxa 2011, 3065, 60–65. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Endangered and threatened wildlife and plants: Endangered status for 49 species from the Hawaiian islands, Proposed Rule. 50 CFR Part 17. Fed. Regist. 2015, 81, 67786–67860. [Google Scholar]
- Altaba, C.R. Extinction Resilience of Island Species: An Amphibian Case and a Predictive Model. Diversity 2014, 6, 43–71. [Google Scholar] [CrossRef]
- Harter, D.E.V.; Irl, S.D.H.; Seo, B.; Steinbauer, M.J.; Gillespie, R.; Triantis, K.A.; Fernández-Palacios, J.-M.; Beierkuhnlein, C. Impacts of global climate change on the floras of oceanic islands-Projections, implications and current knowledge. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Ing, K.; Mogren, C.L. Evidence of Competition between Honey Bees and Hylaeus anthracinus (Hymenoptera: Colletidae), an Endangered Hawaiian Yellow-Faced Bee. Pac. Sci. 2020, 74, 75–85. [Google Scholar] [CrossRef]
- Plentovich, S.; Graham, J.R.; Haines, W.P.; King, C.B.A. Invasive ants reduce nesting success of an endangered Hawaiian yellow-faced bee, Hylaeus anthracinus. NeoBiota 2021, 64, 137–154. [Google Scholar] [CrossRef]
- Koch, J.B.; Sahli, H.F. Patterns of flower visitation across elevation and successional gradients in Hawai’i. Pac. Sci. 2013, 67, 253–266. [Google Scholar] [CrossRef]
- Sahli, H.F.; Krushelnycky, P.D.; Drake, D.R.; Taylor, A.D. Patterns of Floral Visitation to Native Hawaiian Plants in Presence and Absence of Invasive Argentine Ants1. Pac. Sci. 2016, 70, 309–322. [Google Scholar] [CrossRef]
- Shay, K.; Drake, D.R.; Taylor, A.D.; Sahli, H.F. Alien insects dominate the plant-pollinator network of a Hawaiian coastal ecosystem. Pacifica 2016, 70, 409–429. [Google Scholar] [CrossRef]
- Corlett, R.T. Invasive aliens on tropical East Asian islands. Biodivers. Conserv. 2010, 19, 411–423. [Google Scholar] [CrossRef]
- Jupiter, S.; Mangubhai, S.; Kingsford, R.T. Conservation of Biodiversity in the Pacific Islands of Oceania: Challenges and Opportunities. Pac. Conserv. Biol. 2014, 20, 206–220. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Cronk, Q.C.B. Islands: Stability, diversity, conservation. Biodivers. Conserv. 1997, 6, 477–493. [Google Scholar] [CrossRef]
- Strange, J.P.; Koch, J.B.; Gonzalez, V.H.; Nemelka, L. Global invasion by Anthidium manicatum (Linnaeus) (Hymenoptera: Megachilidae): Assessing potential distribution in North America and beyond. Biologicals 2011, 13, 2115–2133. [Google Scholar] [CrossRef]
- Freeman, B.G.; Scholer, M.N.; Ruiz-Gutierrez, V.; Fitzpatrick, J.W. Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. Proc. Natl. Acad. Sci. USA 2018, 115, 11982–11987. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas: NEW CLIMATE SURFACES FOR GLOBAL LAND AREAS. Int. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.-L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; et al. Long-term climate change: Projections, commitments and irreversibility. In Climate Change 2013—The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 1029–1136. [Google Scholar]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species: The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Eckert, S.; Hamad, A.; Kilawe, C.J.; Linders, T.E.W.; Ng, W.; Mbaabu, P.R.; Shiferaw, H.; Witt, A.; Schaffner, U. Niche change analysis as a tool to inform management of two invasive species in Eastern Africa. Ecosphere 2020, 11. [Google Scholar] [CrossRef]
- Shrestha, U.B.; Shrestha, B.B. Climate change amplifies plant invasion hotspots in Nepal. Divers. Distrib. 2019, 10, e0134665. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS): Assessing the accuracy of distribution models. J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Koch, J.; Looney, C.; Hopkins, B.; Lichtenberg, E.M. Projected climate change will reduce habitat suitability for bumble bees in the Pacific Northwest. BioRxiv 2019. [Google Scholar] [CrossRef]
- Graham, K.K.; Eaton, K.; Obrien, I.; Starks, P.T. Anthidium manicatum, an invasive bee, excludes a native bumble bee, Bombus impatiens, from floral resources. Biol. Invasions 2018. [Google Scholar] [CrossRef]
- Zhu, G.; Gutierrez Illan, J.; Looney, C.; Crowder, D.W. Assessing the ecological niche and invasion potential of the Asian giant hornet. Proc. Natl. Acad. Sci. USA 2020, 117, 24646–24648. [Google Scholar] [CrossRef]
- Moo-Llanes, D.A. Inferring Distributional Shifts of Asian Giant Hornet Vespa mandarinia Smith in Climate Change Scenarios. Neotrop. Entomol. 2021. [Google Scholar] [CrossRef]
- McDougall, K.L.; Lembrechts, J.; Rew, L.J.; Haider, S.; Cavieres, L.A.; Kueffer, C.; Milbau, A.; Naylor, B.J.; Nuñez, M.A.; Pauchard, A.; et al. Running off the road: Roadside non-native plants invading mountain vegetation. Biol. Invasions 2018, 20, 3461–3473. [Google Scholar] [CrossRef]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Morales-Barbero, J.; Vega-Álvarez, J. Input matters matter: Bioclimatic consistency to map more reliable species distribution models. Methods Ecol. Evol. 2019, 10, 212–224. [Google Scholar] [CrossRef]
- Title, P.O.; Bemmels, J.B. ENVIREM: An expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 2018, 41, 291–307. [Google Scholar] [CrossRef]
- Zhu, G.; Fan, J.; Peterson, A.T. Cautions in weighting individual ecological niche models in ensemble forecasting. Ecol. Model. 2021, 448, 109502. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Rome, Q.; Villemant, C.; Courchamp, F. Can species distribution models really predict the expansion of invasive species? PLoS ONE 2018, 13, e0193085. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Newell, G.; White, M. The effect of sample size on the accuracy of species distribution models: Considering both presences and pseudo-absences or background sites. Ecography 2019, 42, 535–548. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Bradley, A.A.; Schwartz, S.S.; Hashino, T. Sampling Uncertainty and Confidence Intervals for the Brier Score and Brier Skill Score. Weather Forecast. 2008, 23, 992–1006. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Without quality presence-absence data, discrimination metrics such as TSS can be misleading measures of model performance. J. Biogeogr. 2018, 45, 1994–2002. [Google Scholar] [CrossRef]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schröder, B.; Thuiller, W.; et al. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 2017, 40, 913–929. [Google Scholar] [CrossRef]
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pysek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Methods Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | No. of Records on GBIF | No. GeoRef Records on GBIF | Final No. GeoRef Records | Year Documented in Hawai’i | General Native Range |
|---|---|---|---|---|---|---|---|
| Apidae | Apis | mellifera | 150,293 | 134,926 | 50,640 | 1857 | Europe (Snelling 2003) |
| Apidae | Ceratina | arizonensis | 836 | 828 | 101 | 1950 | Southwestern United States (Daly 1973) |
| Apidae | Ceratina | smaragdula | 409 | 318 | 107 | 1998 | Southeast Asia (Hirashima, 1969, Snelling 2003) |
| Apidae | Ceratina | dentipes | 183 | 145 | 70 | 1996 | Southeast Asia (Snelling 2003) |
| Apidae | Xylocopa | sonorina | 155 | 138 | 101 | 1874 | North America (Snelling 2003) |
| Colletidae | Hylaeus | leptocephalus | 589 | 490 | 247 | 1958 | Europe (Snelling 2003) |
| Colletidae | Hylaeus | albonitens | 924 | 172 | 59 | 1995 | Australia (Magnacca & King 2013) |
| Colletidae | Hylaeus | strenuus | 1 | 1 | 2 | 2007 | India (Magnacca 2011) |
| Halictidae | Lasioglossum | imbrex | 0 | 0 | 49 | 2005 | Western North America (Gibbs 2010) |
| Halictidae | Lasioglossum | impavidum | 337 | 326 | 28 | 1994 | Coastal California (Snelling 2003) |
| Halictidae | Lasioglossum | microlepoides | 13,258 | 13,242 | 180 | 2010 | Western North America and Northern Mexico (Magnacca & King 2013) |
| Halictidae | Lasioglossum | puteulanum | 1752 | 1749 | 188 | 2012 | Eastern North America (Gibbs, 2011); USGS (https://www.usgs.gov/media/images/lasioglossum-nr-puteulanum-male-side, Accessed: 3 June 2019) |
| Megachilidae | Lithurgus | scabrosus | 36 | 18 | 14 | 1907 | Europe (Snelling 2003) |
| Megachilidae | Megachile | chlorura | 3 | 1 | 2 | 1988 | Philippines (Snelling, 2003); Southeast Asia (Rasmussen 2012) |
| Megachilidae | Megachile | diligens | 5 | 0 | 15 | 1879 | South Pacific according to Snelling, 2003), Southeast Asia according to Rasmussen 2012 |
| Megachilidae | Megachile | fullawayi | 21 | 17 | 12 | 1921 | Guam according to Cockerell, but likely brought from Asia. (Snelling 2003) |
| Megachilidae | Megachile | gentilis | 731 | 668 | 230 | 1899 | Northwestern USA, species know from southern BC (Snelling 2003); as M. palmarum Perkins |
| Megachilidae | Megachile | lanata | 345 | 302 | 86 | 2012 | Southeast Asia (Gonsalez et al. 2019), India (Magnacca et al. 2013) |
| Megachilidae | Megachile | policaris | 1114 | 891 | 382 | 2018 | Georgia and Florida, west to California and Mexico |
| Megachilidae | Megachile | timberlakei | 10 | 8 | 8 | 1904 | First documented in Hawai’i, probably South Pacific region. (Snelling 2003) |
| Megachilidae | Megachile | umbripennis | 177 | 148 | 39 | 1898 | Northern India and China (Timberlake, 1921) |
| Species | Current Range (km2) | Range Loss (km2) (2070, RCP 8.5) | Range Gain (km2) (2070, RCP 8.5) | % Loss (2070, RCP 8.5) | % Gain (km2) (2070, RCP 8.5) |
|---|---|---|---|---|---|
| A. mellifera | 2562 (12%) * | 168 | 5538 (40.3%) | 6.6% | 216.2% |
| C. dentipes | 889 (4.4%) | 21 | 3987 (24.3%) | 2.4% | 448.5% |
| C. smaragdula | 748 (3.7%) | 0 | 3600 (21.7%) | 0 | 481.3% |
| L. impavidum | 1217 (6.0%) | 171 | 202 (7.0%) | 14.1% | 16.6% |
| L. microlepoides | 364 (1.8%) | 0 | 4827 (25.9%) | 0 | 2425.7% |
| L. puteulanum | 213 (1.0%) | 0 | 4202 (22.0%) | 0 | 1972.8% |
| M. umbripennis | 377 (1.8%) | 0 | 3308 (18.3%) | 0 | 1769.0% |
| X. sonorina | 2958 (14.7%) | 13 | 4390 (36.6%) | 0.4% | 148.4% |
| Species | Invasive Model TSS | Invasive Model AUC | Native Model TSS | Native Model AUC |
|---|---|---|---|---|
| A. mellifera | 0.731 | 0.937 | 0.921 | 0.995 |
| C. dentipes | 0.958 | 0.993 | 0.815 | 0.956 |
| C. smaragdula | 0.917 | 0.987 | 0.759 | 0.939 |
| L. impavidum | 0.94 | 0.975 | 0.807 | 0.936 |
| L. microlepoides | 0.983 | 0.997 | 0.602 | 0.869 |
| L. puteulanum | 0.991 | 0.998 | 0.796 | 0.954 |
| M. umbripennis | 0.983 | 0.998 | 0.907 | 0.976 |
| X. sonorina | 0.753 | 0.943 | 0.63 | 0.89 |
| Species | Native Model | Invasive Model |
|---|---|---|
| A. mellifera | 0.8 | 0.5 |
| C. dentipes | 0.5 | 0.8 |
| C. smaragdula | 0.8 | 0.8 |
| L. impavidum | 0.5 | 0.5 |
| L. microlepoides | 0.5 | 0.8 |
| L. puteulanum | 0.5 | 0.8 |
| M. umbripennis | 0.5 | 0.5 |
| X. sonorina | 0.5 | 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabor, J.A.; Koch, J.B. Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i. Insects 2021, 12, 443. https://doi.org/10.3390/insects12050443
Tabor JA, Koch JB. Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i. Insects. 2021; 12(5):443. https://doi.org/10.3390/insects12050443
Chicago/Turabian StyleTabor, Jesse A., and Jonathan B. Koch. 2021. "Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i" Insects 12, no. 5: 443. https://doi.org/10.3390/insects12050443
APA StyleTabor, J. A., & Koch, J. B. (2021). Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i. Insects, 12(5), 443. https://doi.org/10.3390/insects12050443