
Sterile Insect Technique in an Integrated Vector Management Program against Tiger Mosquito Aedes albopictus in the Valencia Region (Spain): Operating Procedures and Quality Control Parameters


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Risk Assessment
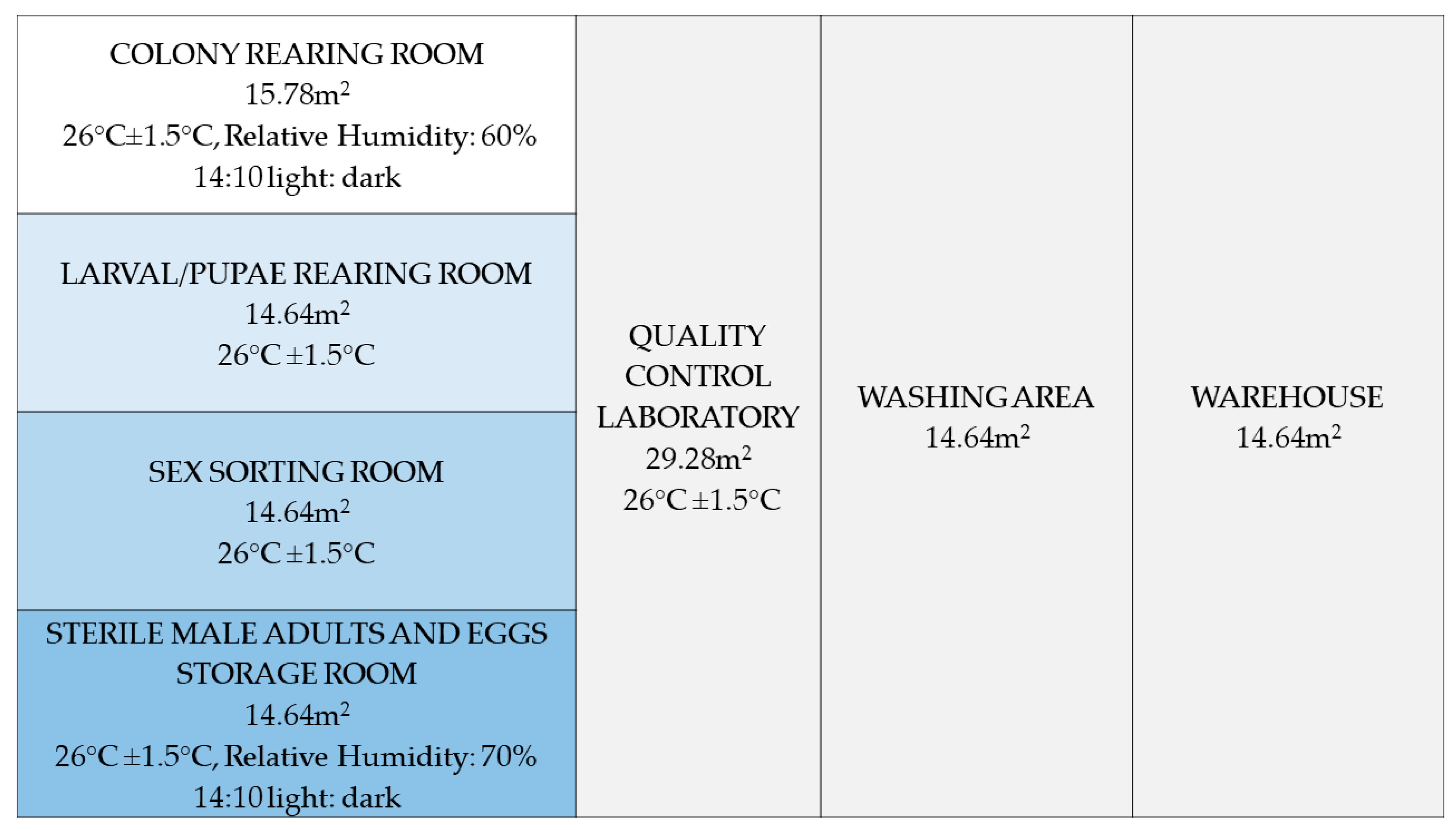
2.2. Rearing Colony
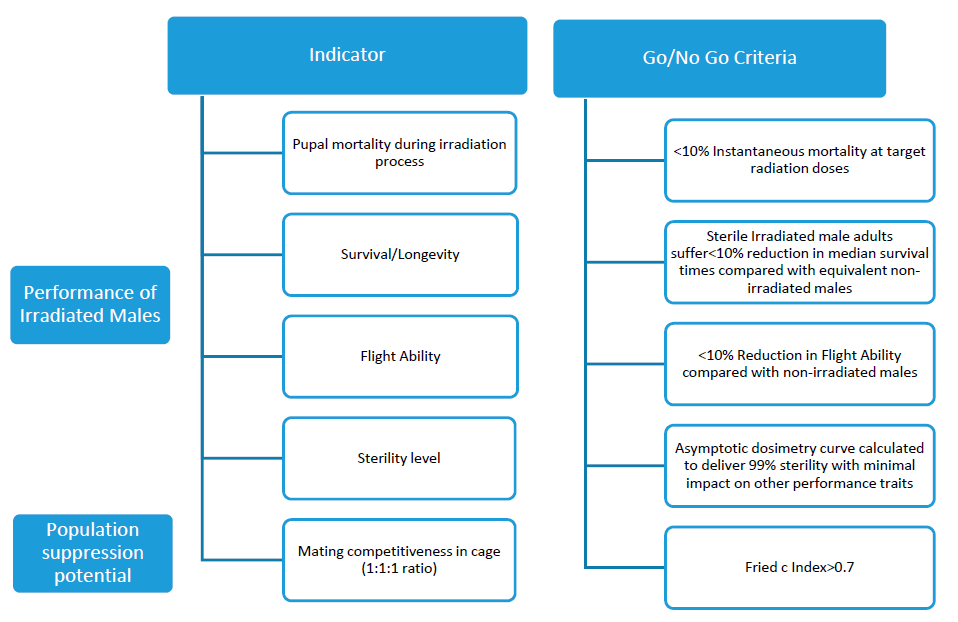
2.3. Laboratory Studies for Evaluating the Entomological Efficacy of SIT Based on WHO-IAEA Phase Conditional Approach
- Determination of irradiation dose. The target irradiation dose for the male pupae was established as the dose required to obtain 99% sterility in males. To estimate the target dose, six batches of 75 pupae that were 24–36 h old were irradiated with different doses (0, 20, 40, 50, 60 and 80 Gy) in a ring-shaped container with inner and outer radii of 2 and 4 cm, placed in the geometrical center of the irradiation chamber. A sample of 50 pupae per batch was selected to emerge in a 24.5 × 24.5 × 24.5-cm BugDorm-4S2222 insect rearing cages (MegaView Science Co., Taichung, Taiwan) and males were allowed to mate with 50 virgin females for 5 days. After three days of blood feeding, the females were isolated in Drosophila vials that contained water and filter paper to stimulate oviposition. The eggs oviposited by each female were counted, dried for four days and allowed to hatch for 48 h by submerging the filter papers in a solution of nutrient broth. The ratio of hatched eggs over the total was estimated for each female.
- Survival. Two batches of irradiated male pupae were selected for the survival analysis. A sample of 50 pupae of each batch was allowed to emerge in BugDorm-4S2222 insect rearing cages (MegaView Science Co., Taichung, Taiwan) with access to a 10% sucrose solution. Simultaneously, 50 non-irradiated male pupae (control) were allowed to emerge under the same conditions. The dead males were counted daily in the following 20 days.
- Instant pupal mortality during irradiation. The number of dead pupae after irradiation was counted in four irradiation containers for 3 days (n = 12). Before introduction in the irradiator, the dead pupae were removed, and the dead pupae immediately after irradiation were counted.
- Pupal emergence and flight ability. A flight organ device was used to estimate both pupal emergence and flight ability [28]. Each flight organ was composed of 120 tubes (diameter, 10.1 mm; length, 400 mm). Eight paired simultaneous tests were conducted (control vs. irradiated).
- Mating competitiveness. BugDorm-4S2222 insect rearing cages (MegaView Science Co., Taichung, Taiwan) were filled with different ratios of untreated and irradiated male mosquitoes to estimate the competitiveness of irradiated males. Three cages were used with 30 individuals of each category: sterile males, non-irradiated males and virgin females. Three additional cages with 30 individuals per category contained only sterile males and females and another three contained only non-irradiated males and females. After 5 days, three blood meals were offered to the females, and an oviposition cup with filter paper and water was introduced into each cage. The number of eggs was counted for each cage, and the eggs were allowed to hatch in a solution of nutrient broth for 24 h. The ratio of L1 larvae versus the number of eggs was used as a hatching rate index. The hatching rates of the different mosquito combinations were used to estimate the Fried competitiveness value c [29].
2.4. Rearing Parameters and Routine Quality Control Measures
- Adult colony rearing
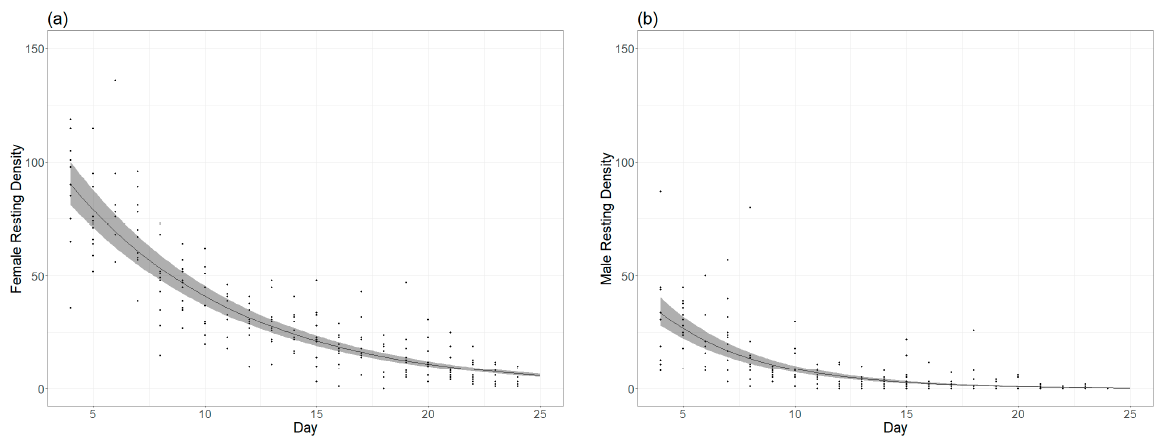
- Survival rate in the adult rearing cages. To estimate the survival of individuals, a triangular, transparent 16 × 16 × 22.6-cm Plexiglas sheet was placed in the upper right corner of a randomly selected cage assembled each day. Every day at the same time, during the lifetime of the cage, the number of resting male and female mosquitoes was estimated as a relative measure of the survival of the mosquitoes.
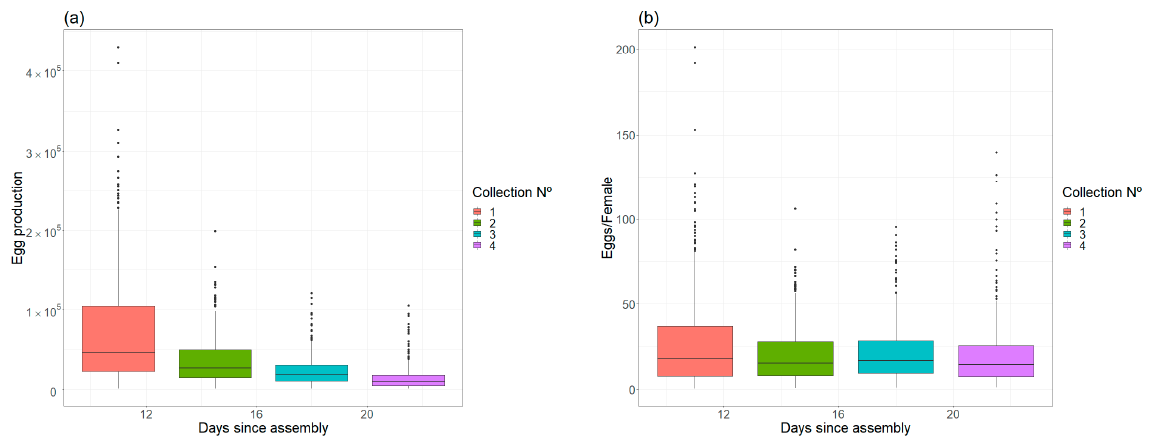
- Egg production and female fecundity. The number of eggs on each oviposition filter paper was visually estimated by an expert technician by comparison with a set of reference oviposition filter papers for which the number of eggs had been estimated [35]. For each oviposition paper, female fecundity was estimated as the egg number divided by the expected number of active females. The expected number of females for a given collection day was approximated using the female survival model. The accumulated fecundity for each cage was estimated as the sum of fecundities of their collected oviposition papers.
- Larval rearing
- Hatching rate. A sample of 2 to 6 oviposition papers was randomly selected after their use in the hatching jars. A random sample of about 200 eggs along the surface of each paper was selected, and the number of hatched and unhatched eggs was counted under a stereomicroscope.
- Pupation at sex sorting and sterile male pupae production. Eight days after eggs hatching, pupae were sexed. Prior to sex sorting, a larval rearing tray was chosen at random and used as a sample for the estimation of pupation parameters. The sample trays were sorted and counted simultaneously with the sex sorting of the production trays. The number of individuals in the three categories of the output (male pupae, female pupae and larvae) was estimated volumetrically. Three tubes (one for each category) were specifically designed for this purpose with a perforated base and graduated/calibrated to 100 individuals. The percentage survival at sex sorting was estimated as the sum of three categories divided by 10,000. The pupation rate at sex sorting was estimated as the sum of male and female pupae divided by the total number of individuals. The percentage of male pupae was estimated as the number of male pupae divided by the total number of pupae at sex sorting. Concerning the total production of male pupae to be sterilized, it was obtained from volumetric dosing into cups for irradiation.
- Sex sorting
- Residual female contamination. The sex sorting of 240,000 males per week involved two technicians five days a week working eight-hour days. As the residual contamination of females in sex sorting depends in part on the skill of the technician, from each of the release batches, 5 cages from each technician were randomly selected on the third day after irradiation and visually checked for females. All females found in the release batches were removed by means of a manual aspirator. A total of 830 batches were visually inspected for the presence of females after the emergence of adults. If the average residual percentage of females for the technician exceeded 1%, 5 more cages were randomly selected. If this percentage was maintained at a level higher than 1%, the entire batch was checked and females were removed before leaving for the field.
2.5. Statistical Analysis
3. Results
3.1. Laboratory Studies
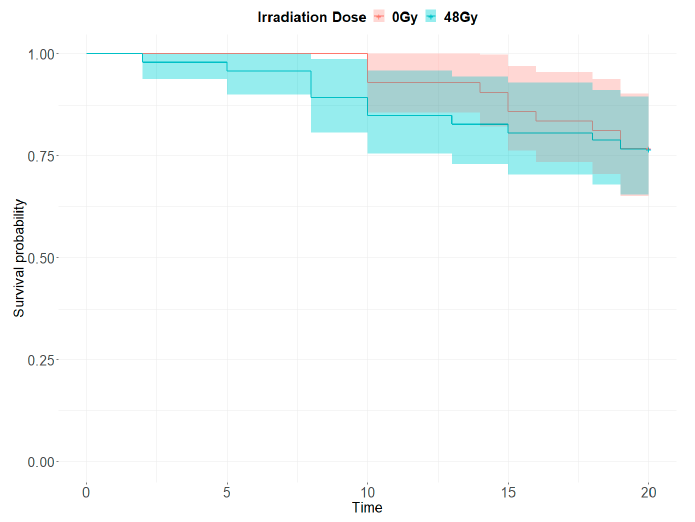
- Survival. In both control and irradiated groups, at least half of the mosquitoes survived more than 20 days. There were no significant differences in the longevity of the irradiated mosquitoes (48 Gy) compared to the control treatment (0 Gy) (hazard ratio = 1.194, z = 0.661, p = 0.509) (Figure 4).
- Instant pupal mortality during irradiation. On average, 0.49% of the pupae died during the irradiation process (SD = 0.27).
- Pupal emergence and flight ability. On average, 97.1% of the irradiated pupae and 97.6% of the non-irradiated emerged to adult. Differences were not statistically significant for untreated and irradiated males (estimate = −0.221, Std. Error = 0.381, z = −0.581, p = 0.561). On average, 85.1% of the irradiated pupae and 90.8% of the non-irradiated succeeded in the flight ability test, which was a statistically significant difference (Estimate = −0.544, SE = 0.201, z = −2.712, p = 0.007).
- Mating competitiveness. The Fried c value was on average 0.92.
3.2. Rearing Parameters and Quality Control Measures
- Adult colony rearing
- ii.
- Larval rearing
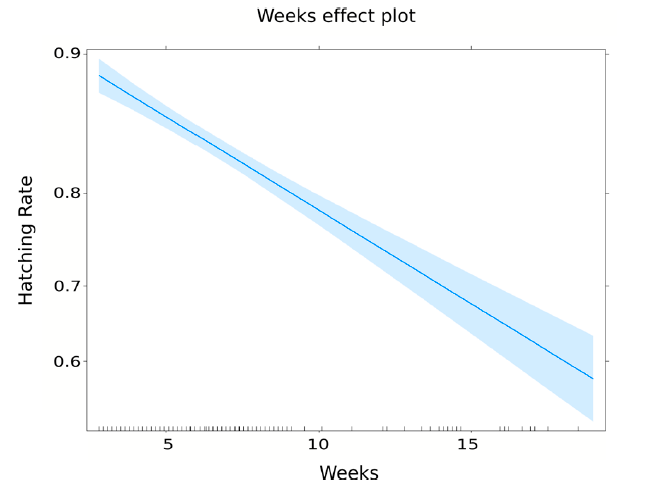
- Hatch rate. Results of egg hatching are shown in Table 5. According to the model, the egg hatching rate was declining with time in storage (Figure 7). After one week of collection, the expected hatching rate was 90.6% (Std. Error = 0.56%). After 5 weeks, it was reduced to 86.2% (Std. Error = 0.43%), and after 20 weeks, it dropped to 54.8% (Std. Error = 3.40%)
- Pupation at sex sorting and male pupae production. A total of 332 trays were used as samples for the estimation of pupation parameters. On average, 0.172 male pupae were recovered at sex sorting from each L1 reared. The specific estimates for the different parameters of pupation are shown in Table 6.
- iii.
- Sex Sorting
- Residual female contamination. The presence of females in the release containers was on average 0.17% of the total individuals (SD = 0.24). Only 0.59% of the batches had a female contamination rate higher than 1%. We estimate that the protocol of inspection reduced the residual presence of females to 0.15%.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the Tiger: Global Risk of Invasion by the Mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Leta, S.; Beyene, T.J.; De Clercq, E.M.; Amenu, K.; Kraemer, M.U.G.; Revie, C.W. Global Risk Mapping for Major Diseases Transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 2018, 67, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Aranda, C.; Eritja, R.; Roiz, D. First Record and Establishment of the Mosquito Aedes albopictus in Spain. Med. Vet. Entomol. 2006, 20, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Goiri, F.; González, M.A.; Goikolea, J.; Oribe, M.; de Castro, V.; Delacour, S.; Lucientes, J.; Ortega-Araiztegi, I.; Barandika, J.F.; García-Pérez, A.L. Progressive Invasion of Aedes albopictus in Northern Spain in The Period 2013–2018 and A Possible Association with The Increase in Insect Bites. Int. J. Environ. Res. Public Health 2020, 17, 1678. [Google Scholar] [CrossRef] [PubMed]
- Monge, S.; García-Ortúzar, V.; López Hernández, B.; Lopaz Pérez, M.Á.; Delacour-Estrella, S.; Sánchez-Seco, M.P.; Fernández Martinez, B.; García San Miguel, L.; García-Fulgueiras, A.; Sierra Moros, M.J. Characterization of the First Autochthonous Dengue Outbreak in Spain (August–September 2018). Acta Trop. 2020, 205, 105402. [Google Scholar] [CrossRef] [PubMed]
- Factsheet about Dengue. Available online: https://www.ecdc.europa.eu/en/dengue-fever/facts/factsheet (accessed on 23 February 2021).
- Ruche, G.L.; Souarès, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Desprès, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First Two Autochthonous Dengue Virus Infections in Metropolitan France, September 2010. Eurosurveillance 2010, 15, 19676. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid Resistance in Aedes aegypti and Aedes albopictus: Important Mosquito Vectors of Human Diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lees, R.S.; Gilles, J.R.; Hendrichs, J.; Vreysen, M.J.; Bourtzis, K. Back to the Future: The Sterile Insect Technique against Mosquito Disease Vectors. Curr. Opin. Insect Sci. 2015, 10, 156–162. [Google Scholar] [CrossRef]
- Laven, H. Eradication of Culex Pipiens Fatigans through Cytoplasmic Incompatibility. Nature 1967, 216, 383–384. [Google Scholar] [CrossRef]
- Patterson, R.; Weidhaas, D.; Ford, H.; Lofgren, C. Suppression and Elimination of an Island Population of Culex Pipiens Quinquefasciatus with Sterile Males. Science 1970, 168, 1368–1369. [Google Scholar] [CrossRef]
- Lofgren, C.S.; Dame, D.A.; Breeland, S.G.; Weidhaas, D.E.; Jeffery, G.; Kaiser, R.; Ford, H.R.; Boston, M.D.; Baldwin, K.F. Release of Chemosterilized Males for the Control of Anopheles Albimanus in El Salvador. Am. J. Trop. Med. Hyg. 1974, 23, 288–297. [Google Scholar] [CrossRef]
- WHO; IAEA. Guidance Framework for Testing the Sterile Insect Technique as a Vector Control Tool against Aedes-Borne Diseases; WHO: Geneva, Switzerland; IAEA: Geneva, Switzerland, 2020. [Google Scholar]
- Dame, D.A.; Curtis, C.F.; Benedict, M.Q.; Robinson, A.S.; Knols, B.G.J. Historical Applications of Induced Sterilisation in Field Populations of Mosquitoes. Malar. J. 2009, 8 (Suppl. 2), 1–10. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and Sterile Insect Techniques Combined Eliminate Mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Bellini, R.; Medici, A.; Puggioli, A.; Balestrino, F.; Carrieri, M. Pilot Field Trials with Aedes albopictus Irradiated Sterile Males in Italian Urban Areas. J. Med. Entomol. 2013, 50, 317–325. [Google Scholar] [CrossRef]
- Zheng, M.-L.; Zhang, D.-J.; Damiens, D.D.; Lees, R.S.; Gilles, J.R.L. Standard Operating Procedures for Standardized Mass Rearing of the Dengue and Chikungunya Vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)—II—Egg Storage and Hatching. Parasites Vectors 2015, 8, 348. [Google Scholar] [CrossRef]
- Kauffman, E.; Payne, A.; Franke, M.A.; Schmid, M.A.; Harris, E.; Kramer, L.D. Rearing of Culex spp. and Aedes spp. Mosquitoes. Bio Protoc. 2017, 7, e2542. [Google Scholar] [CrossRef]
- Mamai, W.; Maiga, H.; Gárdos, M.; Bán, P.; Somda, N.S.B.; Konczal, A.; Wallner, T.; Parker, A.; Balestrino, F.; Yamada, H.; et al. The Efficiency of a New Automated Mosquito Larval Counter and Its Impact on Larval Survival. Sci. Rep. 2019, 9, 7413. [Google Scholar] [CrossRef]
- Maïga, H.; Mamai, W.; Bimbilé Somda, N.S.; Konczal, A.; Wallner, T.; Herranz, G.S.; Herrero, R.A.; Yamada, H.; Bouyer, J. Reducing the Cost and Assessing the Performance of a Novel Adult Mass-Rearing Cage for the Dengue, Chikungunya, Yellow Fever and Zika Vector, Aedes aegypti (Linnaeus). PLoS Negl. Trop. Dis. 2019, 13. [Google Scholar] [CrossRef]
- FAO; IAEA. Guidelines for Mass-Rearing of Aedes Mosquitoes; FAO: Rome, Italy; IAEA: Vienna, Austria, 2020. [Google Scholar]
- Baughman, T.; Peterson, C.; Ortega, C.; Preston, S.R.; Paton, C.; Williams, J.; Guy, A.; Omodei, G.; Johnson, B.; Williams, H.; et al. A Highly Stable Blood Meal Alternative for Rearing Aedes and Anopheles Mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0006142. [Google Scholar] [CrossRef]
- Lepek, H.; Nave, T.; Fleischmann, Y.; Eisenberg, R.; Tirosh, I. Method and Apparatus for Sex Sorting of Mosquitoes. U.S. Patent US16/628,730, 10 September 2020. [Google Scholar]
- FAO; IAEA. Guidelines for Routine Colony Maintenance of Aedes Mosquito Species; FAO: Rome, Italy; IAEA: Vienna, Austria, 2017. [Google Scholar]
- Puggioli, A.; Balestrino, F.; Damiens, D.; Lees, R.S.; Soliban, S.M.; Madakacherry, O.; Dindo, M.L.; Bellini, R.; Gilles, J.R.L. Efficiency of Three Diets for Larval Development in Mass Rearing Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 819–825. [Google Scholar] [CrossRef]
- Fay, R.; Morlan, H.B. A Mechanical Device for Separating the Developmental Stages, Sexes and Species of Mosquitoes. Mosq. News 1959, 19, 144–147. [Google Scholar]
- Mehta, K.; Parker, A.G. Dosimetry System for SIT. In Manual for Gafchromic Film; International Atomic Energy Agency: Vienna, Austria, 2004. [Google Scholar]
- Balestrino, F.; Puggioli, A.; Carrieri, M.; Bouyer, J.; Bellini, R. Quality Control Methods for Aedes albopictus Sterile Male Production. PLoS Negl. Trop. Dis. 2017, 11, e0005881. [Google Scholar] [CrossRef]
- Fried, M. Determination of Sterile-Insect Competitiveness. J. Econ. Entomol. 1971, 64, 869–872. [Google Scholar] [CrossRef]
- Culbert, N.J.; Balestrino, F.; Dor, A.; Herranz, G.S.; Yamada, H.; Wallner, T.; Bouyer, J. A Rapid Quality Control Test to Foster the Development of Genetic Control in Mosquitoes. Sci. Rep. 2018, 8, 16179. [Google Scholar] [CrossRef]
- Yeap, H.L.; Endersby, N.M.; Johnson, P.H.; Ritchie, S.A.; Hoffmann, A.A. Body Size and Wing Shape Measurements as Quality Indicators of Aedes aegypti Mosquitoes Destined for Field Release. Am. J. Trop. Med. Hyg. 2013, 89, 78–92. [Google Scholar] [CrossRef]
- Dor, A.; Maggiani-Aguilera, A.M.; Valle-Mora, J.; Bond, J.G.; Marina, C.F.; Liedo, P. Assessment of Aedes aegypti (Diptera: Culicidae) Males Flight Ability for SIT Application: Effect of Device Design, Duration of Test, and Male Age. J. Med. Entomol. 2020, 57, 824–829. [Google Scholar] [CrossRef]
- Culbert, N.J.; Gilles, J.R.; Bouyer, J. Investigating the Impact of Chilling Temperature on Male Aedes aegypti and Aedes albopictus Survival. PLoS ONE 2019, 14, e0221822. [Google Scholar] [CrossRef]
- FAO; IAEA; USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies, Version 6.0; International Atomic Energy Agency: Vienna, Austria, 2014; 164p. [Google Scholar]
- Mains, J.W.; Mercer, D.R.; Dobson, S.L. Digital Image Analysis to Estimate Numbers of Aedes Eggs Oviposited in Containers. J. Am. Mosq. Control Assoc. 2008, 24, 496–501. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; ISBN 0-387-98784-3. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Balestrino, F.; Medici, A.; Candini, G.; Carrieri, M.; Maccagnani, B.; Calvitti, M.; Maini, S.; Bellini, R. Gamma Ray Dosimetry and Mating Capacity Studies in the Laboratory on Aedes albopictus Males. J. Med. Entomol. 2010, 47, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Parker, A.; Oliva, C.; Balestrino, F.; Gilles, J. X-ray-Induced Sterility in Aedes albopictus (Diptera: Culicidae) and Male Longevity Following Irradiation. J. Med. Entomol. 2014, 51, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Bond, J.G.; Osorio, A.R.; Avila, N.; Gómez-Simuta, Y.; Marina, C.F.; Fernández-Salas, I.; Liedo, P.; Dor, A.; Carvalho, D.O.; Bourtzis, K.; et al. Optimization of Irradiation Dose to Aedes aegypti and Ae. Albopictus in a Sterile Insect Technique Program. PLoS ONE 2019, 14, e0212520. [Google Scholar] [CrossRef]
- Du, W.; Hu, C.; Yu, C.; Tong, J.; Qiu, J.; Zhang, S.; Liu, Y. Comparison between Pupal and Adult X-Ray Radiation, Designed for the Sterile Insect Technique for Aedes albopictus Control. Acta Trop. 2019, 199, 105110. [Google Scholar] [CrossRef]
- Yamada, H.; Maiga, H.; Juarez, J.; Carvalho, D.D.O.; Mamai, W.; Ali, A.; Bimbile-Somda, N.S.; Parker, A.G.; Zhang, D.; Bouyer, J. Identification of Critical Factors That Significantly Affect the Dose-Response in Mosquitoes Irradiated as Pupae. Parasites Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef]
- Yamada, H.; Maiga, H.; Bimbile-Somda, N.S.; Carvalho, D.O.; Mamai, W.; Kraupa, C.; Parker, A.G.; Abrahim, A.; Weltin, G.; Wallner, T.; et al. The Role of Oxygen Depletion and Subsequent Radioprotective Effects during Irradiation of Mosquito Pupae in Water. Parasites Vectors 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Alphey, L. Genetic Control of Mosquitoes. Annu. Rev. Entomol. 2014, 59, 205–224. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Y.; Sun, Q.; Zheng, X.; Gilles, J.R.L.; Yamada, H.; Wu, Z.; Xi, Z.; Wu, Y. Establishment of a Medium-Scale Mosquito Facility: Tests on Mass Production Cages for Aedes albopictus (Diptera: Culicidae). Parasites Vectors 2018, 11, 189. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, M.; Wu, Y.; Gilles, J.R.L.; Yamada, H.; Wu, Z.; Xi, Z.; Zheng, X. Establishment of a Medium-Scale Mosquito Facility: Optimization of the Larval Mass-Rearing Unit for Aedes albopictus (Diptera: Culicidae). Parasites Vectors 2017, 10, 569. [Google Scholar] [CrossRef]
- Balestrino, F.; Puggioli, A.; Gilles, J.R.L.; Bellini, R. Validation of a New Larval Rearing Unit for Aedes albopictus (Diptera: Culicidae) Mass Rearing. PLoS ONE 2014, 9, e91914. [Google Scholar] [CrossRef]
- Mamai, W.; Maiga, H.; Somda, N.S.B.; Wallner, T.; Konczal, A.; Yamada, H.; Bouyer, J. Aedes aegypti Larval Development and Pupal Production in the FAO/IAEA Mass-Rearing Rack and Factors Influencing Sex Sorting Efficiency. Parasite 2020, 27. [Google Scholar] [CrossRef] [PubMed]
- Medici, A.; Carrieri, M.; Scholte, E.-J.; Maccagnani, B.; Luisa Dindo, M.; Bellini, R. Studies on Aedes albopictus Larval Mass-Rearing Optimization. J. Econ. Entomol. 2011, 104, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.O.; Nimmo, D.; Naish, N.; McKemey, A.R.; Gray, P.; Wilke, A.B.B.; Marrelli, M.T.; Virginio, J.F.; Alphey, L.; Capurro, M.L. Mass Production of Genetically Modified Aedes aegypti for Field Releases in Brazil. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, F.; Puggioli, A.; Bellini, R.; Petric, D.; Gilles, J.R.L. Mass Production Cage for Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2014, 51, 155–163. [Google Scholar] [CrossRef]
- Damiens, D.; Marquereau, L.; Lebon, C.; Le Goff, G.; Gaudillat, B.; Habchi-Hanriot, N.; Gouagna, L.-C. Aedes albopictus Adult Medium Mass Rearing for SIT Program Development. Insects 2019, 10, 246. [Google Scholar] [CrossRef]
- Iyaloo, D.P.; Facknath, S. Optimization of Aedes albopictus Rearing Procedures: Preliminary Steps towards Large-Scale Rearing of the Species within the Laboratory in Mauritius. J. Entomol. Zool. Stud. 2017, 5, 46–53. [Google Scholar]
- Maïga, H.; Mamai, W.; Bimbilé Somda, N.S.; Wallner, T.; Poda, B.S.; Salvador-Herranz, G.; Argiles-Herrero, R.; Yamada, H.; Bouyer, J. Assessment of a Novel Adult Mass-Rearing Cage for Aedes albopictus (Skuse) and Anopheles Arabiensis (Patton). Insects 2020, 11, 801. [Google Scholar] [CrossRef]
- Deng, L.; Koou, S.; Png, A.; Ng, L.; Lam-Phua, S. A Novel Mosquito Feeding System for Routine Blood-Feeding of Aedes aegypti and Aedes albopictus. Trop. Biomed. 2012, 29, 169–174. [Google Scholar]
- Del Rosario, A. Studies on the Biology of Philippine Mosquitoes. I. Some Bionomic Features of Aedes aegypti.-Philipp, J. Sci. 90 No. 3 pp. 361–370, 2 refs. Manila, 1961. II. Observations on the Life and Behavior of Aedes albopictus (Skuse) in the Laboratory. Philipp. J. Sci. 1963, 92, 89–103. [Google Scholar]
- Hawley, W.A. The Biology of Aedes albopictus. J. Am. Mosq. Control Assoc. 1988, 1, 1–39. [Google Scholar]
- Oliva, C.F.; Damiens, D.; Vreysen, M.J.; Lemperière, G.; Gilles, J. Reproductive Strategies of Aedes albopictus (Diptera: Culicidae) and Implications for the Sterile Insect Technique. PLoS ONE 2013, 8, e78884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lees, R.S.; Xi, Z.; Bourtzis, K.; Gilles, J.R.L. Combining the Sterile Insect Technique with the Incompatible Insect Technique: III-Robust Mating Competitiveness of Irradiated Triple Wolbachia-Infected Aedes albopictus Males under Semi-Field Conditions. PLoS ONE 2016, 11, e0151864. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S.; et al. Efficient Production of Male Wolbachia -Infected Aedes aegypti Mosquitoes Enables Large-Scale Suppression of Wild Populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef]
- Zacarés, M.; Salvador-Herranz, G.; Almenar, D.; Tur, C.; Argilés, R.; Bourtzis, K.; Bossin, H.; Pla, I. Exploring the Potential of Computer Vision Analysis of Pupae Size Dimorphism for Adaptive Sex Sorting Systems of Various Vector Mosquito Species. Parasites Vectors 2018, 11, 656. [Google Scholar] [CrossRef]
- Gilles, J.R.L.; Schetelig, M.F.; Scolari, F.; Marec, F.; Capurro, M.L.; Franz, G.; Bourtzis, K. Towards Mosquito Sterile Insect Technique Programmes: Exploring Genetic, Molecular, Mechanical and Behavioural Methods of Sex Separation in Mosquitoes. Acta Trop. 2014, 132, S178–S187. [Google Scholar] [CrossRef]
- Papathanos, P.A.; Bourtzis, K.; Tripet, F.; Bossin, H.; Virginio, J.F.; Capurro, M.L.; Pedrosa, M.C.; Guindo, A.; Sylla, L.; Coulibaly, M.B.; et al. A Perspective on the Need and Current Status of Efficient Sex Separation Methods for Mosquito Genetic Control. Parasites Vectors 2018, 11, 165–171. [Google Scholar] [CrossRef]
- Koskinioti, P.; Augustinos, A.; Carvalho, D.; Misbah-ul-Haq, M.; Pillwax, G.; Duran de la Fuente, L.; Salvador-Herranz, G.; Argilés-Herrero, R.; Bourtzis, K. Genetic Sexing Strains for the Population Suppression of the Mosquito Vector Aedes aegypti. Philos. Transact. R Soc. B Biol. Sci. 2020. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Misbah-ul-Haq, M.; Carvalho, D.O.; de la Fuente, L.D.; Koskinioti, P.; Bourtzis, K. Irradiation Induced Inversions Suppress Recombination between the M Locus and Morphological Markers in Aedes aegypti. BMC Genet. 2020, 21, 1–10. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Egg Hatching | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|
| Intercept | 1.694 | 0.033 | 50.40 | <0.0001 |
| Dose (Gy) | −0.133 | 0.001 | −70.02 | <0.0001 |
| Sex | Estimate | Std. Error | z Value | p-Value | |
|---|---|---|---|---|---|
| Female | Intercept | 4.425 | 0.100 | 44.14 | <0.0001 |
| day | −0.226 | 0.005 | −41.69 | <0.0001 | |
| Male | Intercept | 5.022 | 0.056 | 89.09 | <0.0001 |
| day | −0.131 | 0.002 | −61.75 | <0.0001 |
| Average Egg Production/Month | No. Eggs/Cage | No. Eggs/Female Pupae | |||
|---|---|---|---|---|---|
| Year | Average | SD | Average | SD | |
| 2018 | 2,661,583 | 60,836 | 55,306 | 8.79 | 7.99 |
| 2019 | 7,946,500 | 140,854 | 123,474 | 20.35 | 17.84 |
| 2020 | 8,590,583 | 145,603 | 87,262 | 21.03 | 12.61 |
| Estimate | Std. Error | F-Value | p-Value | ||
|---|---|---|---|---|---|
| Egg production | Intercept | 12.002 | 0.112 | 110,483.4 | <0.0001 |
| day | −0.127 | 0.007 | 365.1 | <0.0001 | |
| Fecundity | Intercept | 2.782 | 0.099 | 11,227.690 | <0.0001 |
| day | 0.001 | 0.006 | 0.046 | 0.829 |
| Egg Hatching | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|
| Intercept | 2.379 | 0.075 | 31.52 | <0.0001 |
| Weeks | −0.109 | 0.010 | −10.74 | <0.0001 |
| Parameter | Mean | SD |
|---|---|---|
| % Survival at sex sorting | 83.56 | 17.87 |
| % Pupation rate at sex sorting | 29.03 | 11.97 |
| % Male pupae at sex sorting | 75.66 | 8.6 |
| Sorted male pupae/tray | 1720.18 | 465.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tur, C.; Almenar, D.; Benlloch-Navarro, S.; Argilés-Herrero, R.; Zacarés, M.; Dalmau, V.; Pla, I. Sterile Insect Technique in an Integrated Vector Management Program against Tiger Mosquito Aedes albopictus in the Valencia Region (Spain): Operating Procedures and Quality Control Parameters. Insects 2021, 12, 272. https://doi.org/10.3390/insects12030272
Tur C, Almenar D, Benlloch-Navarro S, Argilés-Herrero R, Zacarés M, Dalmau V, Pla I. Sterile Insect Technique in an Integrated Vector Management Program against Tiger Mosquito Aedes albopictus in the Valencia Region (Spain): Operating Procedures and Quality Control Parameters. Insects. 2021; 12(3):272. https://doi.org/10.3390/insects12030272
Chicago/Turabian StyleTur, Carlos, David Almenar, Sandra Benlloch-Navarro, Rafael Argilés-Herrero, Mario Zacarés, Vicente Dalmau, and Ignacio Pla. 2021. "Sterile Insect Technique in an Integrated Vector Management Program against Tiger Mosquito Aedes albopictus in the Valencia Region (Spain): Operating Procedures and Quality Control Parameters" Insects 12, no. 3: 272. https://doi.org/10.3390/insects12030272
APA StyleTur, C., Almenar, D., Benlloch-Navarro, S., Argilés-Herrero, R., Zacarés, M., Dalmau, V., & Pla, I. (2021). Sterile Insect Technique in an Integrated Vector Management Program against Tiger Mosquito Aedes albopictus in the Valencia Region (Spain): Operating Procedures and Quality Control Parameters. Insects, 12(3), 272. https://doi.org/10.3390/insects12030272