
Breeding Enhancement of Musca domestica L. 1758: Egg Load as a Measure of Optimal Larval Density

 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. House Fly Colony
2.2. Effect of Egg Load on Larval Productivity
2.3. Evaluation
2.4. Statistical Analyses
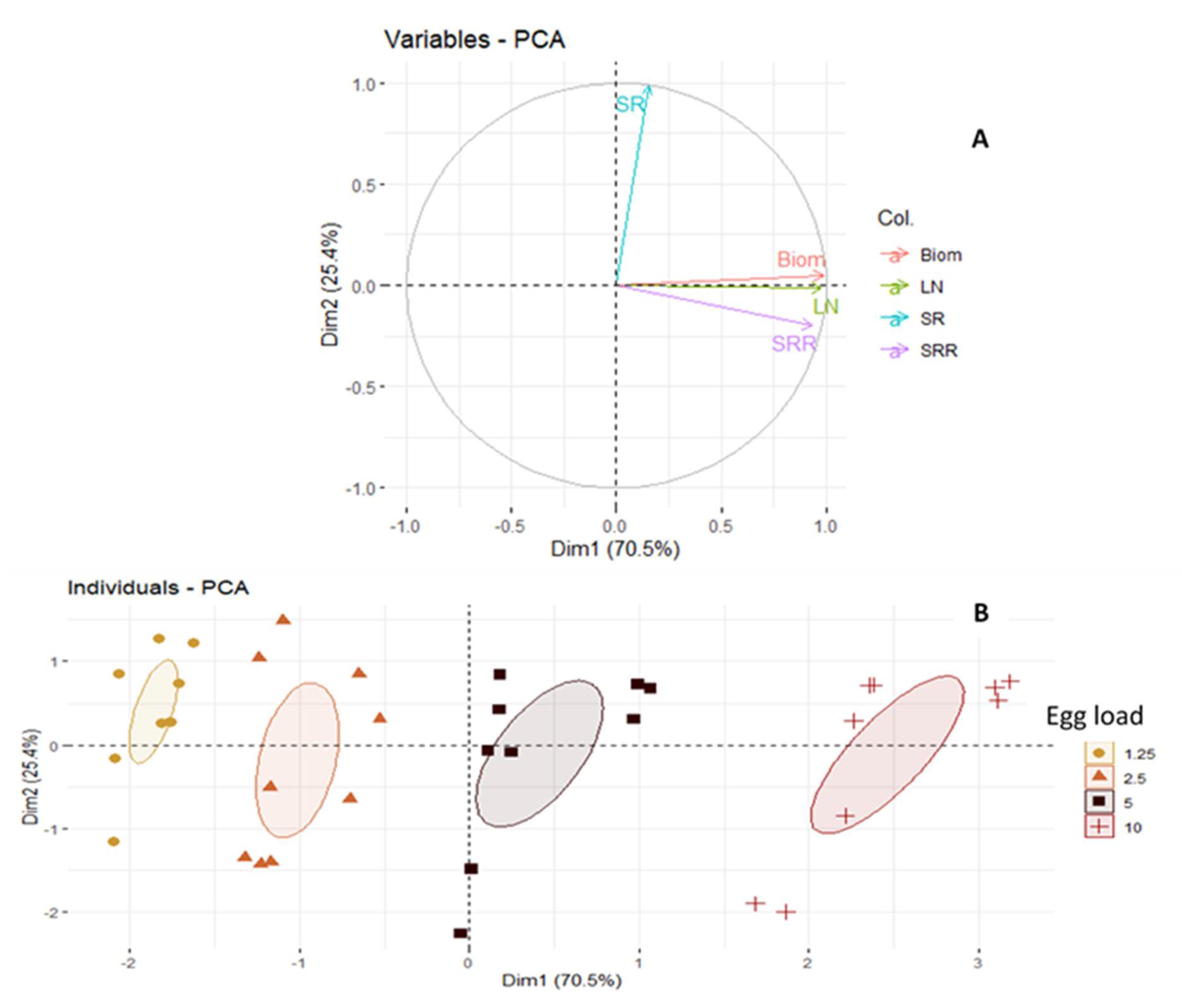
3. Results
Effect of Substrate Type and Egg Load of Musca domestica on Maggot Development and Waste Reduction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaza, S.; Yao, L.C.; Bhada-Tata, P.; Van Woderden, F. What a Waste 2.0: A Global Snapshot of Solid Waste Management to 2050; Urban development; World Bank: Washington, DC, USA, 2018. [Google Scholar]
- Adamou, P.O. Diagnosis of the Household Solid Waste Management System of the 4th District of the City of Niamey (Niger) and Proposal of Improvement Solutions. Master’s Thesis; Burkina Faso, 2015. Available online: www.secheresse.info/spip.php?article57509 (accessed on 3 August 2021).
- Tini, A. Solid Waste Management in Niamey, Niger Trial for Sustainable Management Strategy. Ph.D. Thesis, Institute of Applied Sciences, Lyon, France, 2003. [Google Scholar]
- United Nation. World Population Prospects—Population Division—United Nations. 2021. Available online: https://population.un.org/wpp/2021 (accessed on 3 August 2021).
- Niger. Summary of the Results of the Pastoral Campaign 2015–2016: Draft Version; Ministry of Livestock: Niamey, Niger, 2015. [Google Scholar]
- Aoike, U. Household waste and local solid waste collection in Niamey, Republique of Niger. Afr. Study Monogr. Suppl. 2019, 58, 93–114. [Google Scholar] [CrossRef]
- Gao, A.; Tian, Z.; Wang, Z.; Wennersten, R.; Sun, Q. Comparison between the technologies for food waste treatment. Energy Procedia 2017, 105, 3915–3921. [Google Scholar] [CrossRef]
- Cheng, J.Y.; Chiu, S.L.; Lo, I.M. Effects of moisture content of food waste on residue separation, larval growth and larval survival in black soldier fly bioconversion. Waste Manag. 2017, 67, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef] [Green Version]
- Pastor, B.; Velásquez, Y.; Gobbi, P.; Rojo, S. Conversion of organic wastes into fly larval biomass: Bottlenecks and challenges. J. Insects Food Feed. 2015, 1, 179–193. [Google Scholar] [CrossRef]
- Adeniji, A.A. Effect of replacing groundnut cake with maggot meal in the diet of broilers. Int. J. Poult. Sci. 2007, 6, 822–825. [Google Scholar] [CrossRef] [Green Version]
- Bouafou, K.G.M. Revue bibliographique sur les asticots et leur emploi dans l’alimentation animale. J. Anim. Plant Sci. 2011, 12, 1543–1551. [Google Scholar]
- Fasakin, E.A.; Balogun, A.M.; Ajayi, O.O. Evaluation of full-fat and defatted maggot meals in the feeding of clariid catfish Clarias gariepinus fingerlings. Aquac. Res. 2003, 34, 733–738. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Chatzifotis, S.; Piccolo, G. Does dietary insect meal affect the fish immune system? The case of mealworm, Tenebrio molitor on European sea bass, Dicentrarchus labrax. Dev. Comp. Immunol. 2018, 81, 204–209. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed. Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Moula, N.; Detilleux, J. A Meta-Analysis of the effects of insects in feed on poultry growth performances. Animals 2019, 9, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunji, J.; Kloas, W.; Wirth, M.; Schulz, C.; Rennert, B. Housefly maggot meal (mag meal) as a protein source for Oreochromis niloticus (Linn). Asian Fish. Sci. 2008, 21, 319–331. [Google Scholar]
- Pieterse, E.; Pretorius, Q. Nutritional evaluation of dried larvae and pupae meal of the housefly (Musca domestica) using chemical- and broiler-based biological assays. Anim. Prod. Sci. 2014, 54, 347–355. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO forestry paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Bloukounon-Goubalan, A.Y.; Saïdou, A.; Obognon, N.; Amadji, G.L.; Igue, A.M.; Clottey, V.A.; Kenis, M. Decomposition and nutrient release pattern of animal manures biodegraded by fly larvae in Acrisols. Can. J. Soil Sci. 2019, 99, 60–69. [Google Scholar] [CrossRef]
- Coulibaly, K.; Sankara, F.; Pousga, S.; Nacoulma, P.; Some, M.; Nacro, H. Effects of poultry litter and the residues of maggot’s production on chemical fertility of a lixisol and maize (Zea mays L.) yield in western of Burkina Faso. Niger. J. Soil Sci. 2020, 30, 95–102. [Google Scholar]
- Coulibaly, K.; Sankara, F.; Pousga, S.; Nacoulma, P.J.; Somé, M.B.; Nacro, H.B. On station maggot production using poultry-litter as substrate: Assessment on the quantity and the chemical quality of the litter before and after maggot production in Burkina Faso. Int. J. Biol. Chem. Sci. 2020, 14, 1689–1697. [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Li, H.; Wang, B.; Zhou, Q.; Zhang, X.; Li, J.; Zhang, Z. Decomposition and humification of dissolved organic matter in swine manure during housefly larvae composting. Waste Manag. Res. 2016, 34, 465–473. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, H.; Zhu, J.; Suneethi, S.; Zheng, J. Swine manure vermicomposting via housefly larvae (Musca domestica): The dynamics of biochemical and microbial features. Bioresour. Technol. 2012, 118, 563–571. [Google Scholar] [CrossRef]
- Fowles, T.M.; Nansen, C. Insect-Based Bioconversion: Value from Food Waste; Palgrave Macmillan: Cham, Switzerland, 2019; pp. 321–346. [Google Scholar] [CrossRef] [Green Version]
- Barnard, D.; Harms, R.; Sloan, D. Biodegradation of poultry manure by house fly (Diptera: Muscidae). Environ. Entomol. 1998, 27, 600–605. [Google Scholar] [CrossRef]
- Koo, S.; Currin, T.; Johson, M.; King, E.; Turk, D. La valeur nutritive et la teneur microbienne des pupes séchées de mouche de visage (Musca autumnalis (De Geer)) une fois alimentées aux poussins. Poult. Sci. 1998, 59, 2514–2518. [Google Scholar] [CrossRef]
- Roffeis, M.; Muys, B.; Almeida, J.; Mathijs, E.; Achten, W.; Pastor, B.; Velásquez, Y.; Martínez-Sánchez, A.; Rojo, S. Pig manure treatment with housefly (Musca domestica) rearing—An environmental life cycle assessment. J. Insects Food Feed. 2015, 1, 195–214. [Google Scholar] [CrossRef] [Green Version]
- Surendra, K.; Olivier, R.; Tomberlin, J.K.; Jha, R.; Khanal, S.K. Bioconversion of organic wastes into biodiesel and animal feed via insect farming. Renew. Energy 2016, 98, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Yehuda, B.; Marchaim, U.; Glatman, L.; Drabkin, V.; Chizov-Ginzburg, A.; Mumcuoglu, K.Y.; Gelman, A. Bioconversion of poultry and fish waste by Lucilia sericata and Sarcophaga carnaria larvae. Asian J. Water Environ. Pollut. 2011, 8, 69–75. [Google Scholar]
- Abraham, K.O.; Richard, A.A.; Aihoon, S. Proximate composition of housefly (Musca domestica) maggots cultured on different substrates as potential feed for Tilapia (Oreochromis niloticus). Int. J. Multidiscip. Res. Dev. 2015, 2, 102–103. [Google Scholar]
- Arong, G.A.; Eyo, V.O. Evaluation of housefly (Musca domestica) maggot meal and termite (Macrotermes subhyalinus) meal as supplementary feed for african catfish Clarias gariepinus (Burchell, 1822). Int. J. Entomol. Nematol. 2017, 3, 43–50. [Google Scholar]
- Djissou, A.S.M.; Tossavi, E.C.; Vodounnou, J.D.; Toguyeni, A.; Fiogbe, E.D. Valorization of agro-alimentary waste for a production of maggots like source of proteins in the animal feeds. Int. J. Agron. Agric. Res. 2015, 7, 42–46. [Google Scholar]
- Ekoue, S.E.; Hadzi, Y.A. Production d’asticots comme source de protéines pour jeunes volailles au Togo—Observations préliminaires. Tropicultura 2000, 18, 2012–2014. [Google Scholar]
- Ezewudo, B.I.; Moneb, C.O.; Ugwumba, A.A.A. Production and utilization of Musca domestica maggots in the diet of Oreochromis niloticus (Linnaeus, 1758) fingerlings. Afr. J. Agric. Res. 2015, 10, 2363–2371. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.; Pillai, V.V.; Goddard, J.M.; Park, H.G.; Kothapalli, K.S.; Ross, D.A.; Ketterings, Q.M.; Brenna, J.T.; Milstein, M.B.; Marquis, H.; et al. Sustainable production of housefly (Musca domestica) larvae as a protein-rich feed ingredient by utilizing cattle manure. PLoS ONE 2017, 12, e0171708. [Google Scholar] [CrossRef]
- Koné, N.; Sylla, M.; Nacambo, S.; Kenis, M. Production of Housefly larvae for animal feed through natural oviposition. J. Insects Food Feed 2017, 3, 177–186. [Google Scholar] [CrossRef]
- Mpoame, M.; Téguia, A.; Nguemfo, E.L. Essai comparé de production d’asticots dans les fientes de poule et dans la bouse de vache. Tropicultura 2004, 22, 84–87. [Google Scholar]
- Niu, Y.; Heng, D.; Yao, B.; Cai, Z.; Zhao, Z.; Wu, S.; Cong, P.; Yang, D. A novel bioconversion for value-added products from food waste using Musca domestica. Sci. Directs 2017, 61, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Odesanya, B.O.; Ajayi, S.O.; Agbaogun, B.K.O.; Okuneye, B. Comparative evaluation of nutritive value of maggots. Int. J. Sci. Eng. Res. 2011, 2, 1–5. [Google Scholar]
- Tobin, P.; Pitts, C.W. Geostatistical analysis and the impact of moisture on the spatial and temporal distribution of larval Musca domestica (Diptera: Muscidae). Environ. Entomol. 2002, 31, 273–280. [Google Scholar] [CrossRef]
- Leyo, I.H.; Ousman, Z.M.; Francis, F.; Megido, R.C. Techniques de production d’asticots de mouches domestiques (Musca domestica L. 1758) pour l’alimentation des volailles, synthèse bibliographique. Trop 2021. [Google Scholar] [CrossRef]
- Barnard, D.R.; Geden, C.J. Influence of larval density and temperature in poultry manure on development of the house fly (Diptera: Muscidae). Environ. Entomol. 1993, 22, 971–977. [Google Scholar] [CrossRef]
- Bellows, T.S. The descriptive properties of some models for density dependence. J. Anim. Ecol. 1981, 50, 139. [Google Scholar] [CrossRef]
- Pieterse, E.; Gloy, E. Determination of the influence of a constant nutrient supply on wet yield, dry yield and average weight of Musca domestica larvae maintained at different densities. Afr. Entomol. 2013, 21, 239–242. [Google Scholar] [CrossRef]
- Sullivan, R.L.; Sokal, R.R. The effects of larval density on several strains of the house fly. Ecology 1963, 44, 120–130. [Google Scholar] [CrossRef]
- Teotia, J.S.; Miller, B.F. Environmental conditions affecting development of house fly larvae in poultry manure. Environ. Entomol. 1973, 2, 329–334. [Google Scholar] [CrossRef]
- Kenis, M.; Koné, N.; Chrysostome, C.; Devic, E.; Koko, G.; Clottey, V.; Nacambo, S.; Mensah, G. Insects used for animal feed in West Africa. Entomologia 2014, 2. [Google Scholar] [CrossRef]
- Aniebo, A.O.; Erondu, E.S.; Owen, O.J. Proximate composition of housefly larvae (Musca domestica) meal generated from mixture of cattle blood and wheat bran. Livest. Res. Rural Dev. 2008, 20, 1–5. [Google Scholar]
- Hwangbo, J.; Hong, E.C.; Jan, A.; Kang, H.K.; Oh, J.S.; Kim, B.W.; Park, B.S. Utilization of housefly-maggots, a feed supplement in the production of broiler chickens. J. Environ. Biol. 2009, 30, 609–614. [Google Scholar] [PubMed]
- Wang, H.; Zhang, Z.; Czapar, G.F.; Winkler, M.K.; Zheng, J. A full-ccale house fly (Diptera: Muscidae) larvae bioconversion system for value-added swine manure reduction. Waste Manag. Res. J. Sustain. Circ. Econ. 2013, 31, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Recensement, G.A.C. Analyse Des Résultats Des Enquêtes Sur Les Marches à Bétail et Le Cheptel Aviaire. Projet GCP/NER/041/EC; Ministère du Développement Agricole, Ministère des Ressources Animales, Union européenne, FAO: Abuja, Niger, 2008; p. 85. [Google Scholar]
- Holmes, L.A.; VanLaerhoven, S.L.; Tomberlin, J.K. Relative humidity effects on the life history of Hermetia illucens (Diptera: Stratiomyidae). Environ. Entomol. 2012, 41, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Keiding, J.; Arevad, K. Procedure and equipment for rearing a large number of housefly strains. Bull. World Heal. Organ. 1964, 31, 527–528. [Google Scholar]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- R. Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- De Mendiburu, D.F. Una Herramienta de Análisis Estadístico para la Investigación Agrícola. Master’s Thesis, Universidad Nacional de Ingenieria (UNI-PERU), Lima, Peru, 2009. Available online: http://cybertesis.uni.edu.pe/handle/uni/14814 (accessed on 3 August 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Diener, S.; Studt Solano, N.M.; Roa Gutiérrez, F.; Zurbrügg, C.; Tockner, K. Biological treatment of municipal organic waste using black soldier fly larvae. Waste Biomass Valorizat. 2011, 2, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Hem, S.; Touré, S.; Sagbla, S.; Legendre, M. Bioconversion of palm kernel meal for aquaculture: Experiences from the forest region (Republic of Guinea). Afr. J. Biotechnol. 2008, 7, 1192–1198. [Google Scholar]
- Yang, D.; Huang, M.; Liang, D.; Li, Z.; Yang, W. Biodiesel and Preparation Method. Thereof. Patent US20120144732A1, 14 June 2012. [Google Scholar]
- Agunbiade, J.A.; Adeyemi, O.; Ashiru, O.M.; Awojobi, H.A.; Taiwo, A.A.; Oke, D.B.; Adekunmisi, A.A. Replacement of fish meal with maggot meal in cassava-based layers’ diets. J. Poult. Sci. 2007, 44, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Barragan-Fonseca, K.B.; Dicke, M.; Van Loon, J.J. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzepe, D.; Nana, P.; Fotso, A.; Tchuinkam, T.; Djouaka, R. Influence of larval density, substrate moisture content and feedstock ratio on life history traits of black soldier fly larvae. J. Insects Food Feed. 2020, 6, 133–140. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Cheng, Z.; Yu, L.; Li, H.; Xu, X.; Yang, Z. Use of housefly (Musca domestica L.) larvae to bioconversion food waste for animal nutrition and organic fertilizer. Environ. Sci. Pollut. Res. 2021, 28, 48921–48928. [Google Scholar] [CrossRef]
- Green, P.; Simmonds, M.S.; Blaney, W.M. Diet nutriment and rearing density affect the growth of black blowfly larvae, Phomia regina (Diptera: Calliphoridae). Eur. J. Entomol. 2003, 100, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Bryant, E.H.; McCommas, A.S.; Combs, L.M. The effect of an experimental bottleneck upon quantitative genetic variation in the housefly. Genetics 1986, 114, 1191–1211. [Google Scholar] [CrossRef]
- Bouafou, K.G.M.; Kouame, K.G.; Amoikon, K.E.; Offoumou, A.M. Potentiel pour la production d’asticots sur des sous-produits en Côte d’Ivoire. Trop 2006, 24, 157–161. [Google Scholar]
- Čičková, H.; Kozánek, M.; Takáč, P. Improvement of survival of the house fly (Musca domestica L.) larvae under mass-rearing conditions. Bull. Entomol. Res. 2012, 103, 119–125. [Google Scholar] [CrossRef]
- Sanou, A.G.; Sankara, F.; Pousga, S.; Coulibaly, K.; Nacoulma, J.P.; Ouedraogo, I.; Nacro, S.; Kenis, M.; Sanon, A.; Somda, I. Production de masse de larves de Musca domestica L. (Diptera: Muscidae) pour l’aviculture au Burkina Faso: Analyse des facteurs déterminants en oviposition naturelle. J. Appl. Biosci. 2019, 134, 13689–13701. [Google Scholar] [CrossRef]
- Bauer, A.; Bauer, A.M.; Tomberlin, J.K. Impact of diet moisture on the development of the forensically important blow fly Cochliomyia macellaria (Fabricius) (Diptera: Calliphoridae). Forensic Sci. Int. 2020, 312, 110333. [Google Scholar] [CrossRef] [PubMed]
- Chia, S.; Tanga, C.M.; Khamis, F.M.; Mohamed, S.A.; Salifu, D.; Sevgan, S.; Fiaboe, K.K.M.; Niassy, S.; Van Loon, J.J.A.; Dicke, M.; et al. Threshold temperatures and thermal requirements of black soldier fly Hermetia illucens: Implications for mass production. PLoS ONE 2018, 13, e0206097. [Google Scholar] [CrossRef] [Green Version]
- Kökdener, M. Impact of diet and moisture content on the development of Musca domestica (Diptera: Muscidae). Environ. Entomol. 2021, 50, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Pastor, B.; Čičková, H.; Kozánek, M.; Martínez-Sánchez, A.; Takáč, P.; Rojo, S. Effect of the size of the pupae, adult diet, oviposition substrate and adult population density on egg production in Musca domestica (Diptera: Muscidae). Eur. J. Entomol. 2011, 108, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.I.; Foglia, M.; Blanckenhorn, W.U. Oviposition site choice in the yellow dung fly Scathophaga stercoraria. Ethology 2001, 105, 423–430. [Google Scholar] [CrossRef]
- Čičková, H.; Pastor, B.; Kozánek, M.; Martínez-Sánchez, A.; Rojo, S.; Takáč, P. Biodegradation of pig manure by the housefly, Musca domestica: A viable ecological strategy for pig manure management. PLoS ONE 2012, 7, e32798. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Parameters | Factors | Df | F Value | p-value |
|---|---|---|---|---|
| Biomass | Egg load | 3 | 1278.95 | p < 0.001 |
| Substrates | 2 | 109.36 | p < 0.001 | |
| Egg load × Substrates | 6 | 21.29 | p < 0.001 | |
| Survival rate | Egg load | 3 | 0.63 | 0.600 |
| Substrates | 2 | 16.31 | p < 0.001 | |
| Egg load × Substrates | 6 | 4.194 | 0.005 | |
| Larval number | Egg load | 3 | 3496.44 | p < 0.001 |
| Substrates | 2 | 20.05 | p < 0.001 | |
| Egg load × Substrates | 6 | 3.16 | 0.019 | |
| Substrate rate reduction | Egg load | 3 | 453.10 | p < 0.001 |
| Substrates | 2 | 58.38 | p < 0.001 | |
| Egg load × Substrates | 6 | 3.03 | 0.023 |
| Parameters | Egg Lad (mg) | Millet Bran | Wheat Bran | Cow Dung | Statistical Analysis |
|---|---|---|---|---|---|
| Biomass (mg) | 1.25 | 412.67 ± 11.93 c (b) | 488.33 ± 9.60 d (b) | 396.33 ± 23.02 d (a) | p < 0.001; Df = 2; F = 28.35 |
| 2.5 | 506.00 ± 95.39 c (c) | 855.33 ± 49.90 c (b) | 726.33 ± 17.09 c (a) | p = 0.001; Df = 2; F = 23.63 | |
| 5 | 1267.00 ± 146.24 b (b) | 1712.33 ± 184.85 b (a) | 1281.33 ± 1.52 b (b) | p = 0.011; Df = 2; F = 10.37 | |
| 10 | 2194.67 ± 61.04 a (c) | 3240.00 ± 76.21 a (b) | 2458.66 ± 63.10 a (a) | p < 0.001; Df = 2; F = 196.80 | |
| Statistical analysis | p < 0.001; Df = 3; F = 238.70 | p < 0.001; Df = 3; F = 421.90 | p < 0.001; Df = 3; F = 2047.00 | ||
| Survival rate (%) | 1.25 | 95.00 ± 5.00 a (a) | 93.33 ± 5.77 a (a) | 85.00 ± 10.00 a (a) | p = 0.272; Df = 2; F = 1.63 |
| 2.5 | 75.00 ± 1.00 b (b) | 91.33 ± 7.63 a (a) | 94.66 ± 10.40 a (a) | p = 0.037; Df = 2; F = 5.98 | |
| 5 | 77.33 ± 10.69 b (b) | 97.00 ± 1.73 a (a) | 93.66 ± 4.50 a (a) | p = 0.025; Df = 2; F = 7.24 | |
| 10 | 75.33 ± 6.65 b (b) | 98.00 ± 1.00 a (a) | 96.66 ± 2.30 a (a) | p < 0.001; Df = 2; F = 28.74 | |
| Statistical analysis | p = 0.018; Df = 3; F = 6.03 | p = 0.363; Df = 3; F = 1.22 | P = 0.322; Df = 3; F = 1.36 | ||
| Larval number | 1.25 | 19.00 ± 1.00 d (a) | 19.66 ± 0.57 d (a) | 20.00 ± 1.00 d (a) | p = 0.422 Df = 2; F = 1.00 |
| 2.5 | 30.66 ± 1.51 c (b) | 37.00 ± 2.00 c (a) | 38.66 ± 1.52 c (a) | p = 0.001; Df = 2; F = 20.91 | |
| 5 | 63.33 ± 8.50 b (b) | 78.01 ± 2.05 b (a) | 75.33 ± 2.51 b (a) | p = 0.030; Df = 2; F = 6.64 | |
| 10 | 150.00 ± 2.00 a (b) | 156.66 ± 1.52 a (a) | 155.00 ± 3.46 a (a) | p = 0.038; Df = 2; F = 5.99 | |
| Statistical analysis | p < 0.001; Df = 3; F = 534.90 | p < 0.001; Df = 3; F = 4187.00 | p < 0.001; Df = 3; F = 1978.00 | ||
| Substrate rate reduction (%) | 1.25 | 05 ± 0.01 d (a) | 05.33 ± 0.57 d (b) | 02.00 ± 1.01 d (a) | p = 0.006; Df = 2; F = 13.00 |
| 2.5 | 12.66 ± 1.52 c (a) | 15.00 ± 1.00 c (b) | 07.66 ± 1.52 c (a) | p = 0.001; Df = 2; F = 22.29 | |
| 5 | 18.66 ± 2.08 b (b) | 23.00 ± 1.02 b (b) | 15.66 ± 0.57 b (a) | p = 0.005; Df = 2; F = 14.20 | |
| 10 | 24.33 ± 0.57 a (b) | 26.00 ± 1.00 a (c) | 22.33 ± 0.58 a (a) | p = 0.002; Df = 2; F = 18.20 | |
| Statistical analysis | p < 0.001; Df = 3; F = 102.90 | p < 0.001; Df = 3; F = 162.10 | p < 0.001; Df = 3; F = 239.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamidou Leyo, I.; Moussa Ousmane, Z.; Noël, G.; Francis, F.; Caparros Megido, R. Breeding Enhancement of Musca domestica L. 1758: Egg Load as a Measure of Optimal Larval Density. Insects 2021, 12, 956. https://doi.org/10.3390/insects12110956
Hamidou Leyo I, Moussa Ousmane Z, Noël G, Francis F, Caparros Megido R. Breeding Enhancement of Musca domestica L. 1758: Egg Load as a Measure of Optimal Larval Density. Insects. 2021; 12(11):956. https://doi.org/10.3390/insects12110956
Chicago/Turabian StyleHamidou Leyo, Idriss, Zakari Moussa Ousmane, Gregoire Noël, Frédéric Francis, and Rudy Caparros Megido. 2021. "Breeding Enhancement of Musca domestica L. 1758: Egg Load as a Measure of Optimal Larval Density" Insects 12, no. 11: 956. https://doi.org/10.3390/insects12110956
APA StyleHamidou Leyo, I., Moussa Ousmane, Z., Noël, G., Francis, F., & Caparros Megido, R. (2021). Breeding Enhancement of Musca domestica L. 1758: Egg Load as a Measure of Optimal Larval Density. Insects, 12(11), 956. https://doi.org/10.3390/insects12110956