
Evaluating the Efficacy of 30 Different Essential Oils against Varroa destructor and Honey Bee Workers (Apis mellifera)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material and Essential Oils
2.2. Screening of Essential Oils for Their Acaricidal Activity
2.3. Complete Exposure Bioassay
2.4. Assessment of the Main Components of the Examined EOs
2.5. Statistical Analyses
3. Results
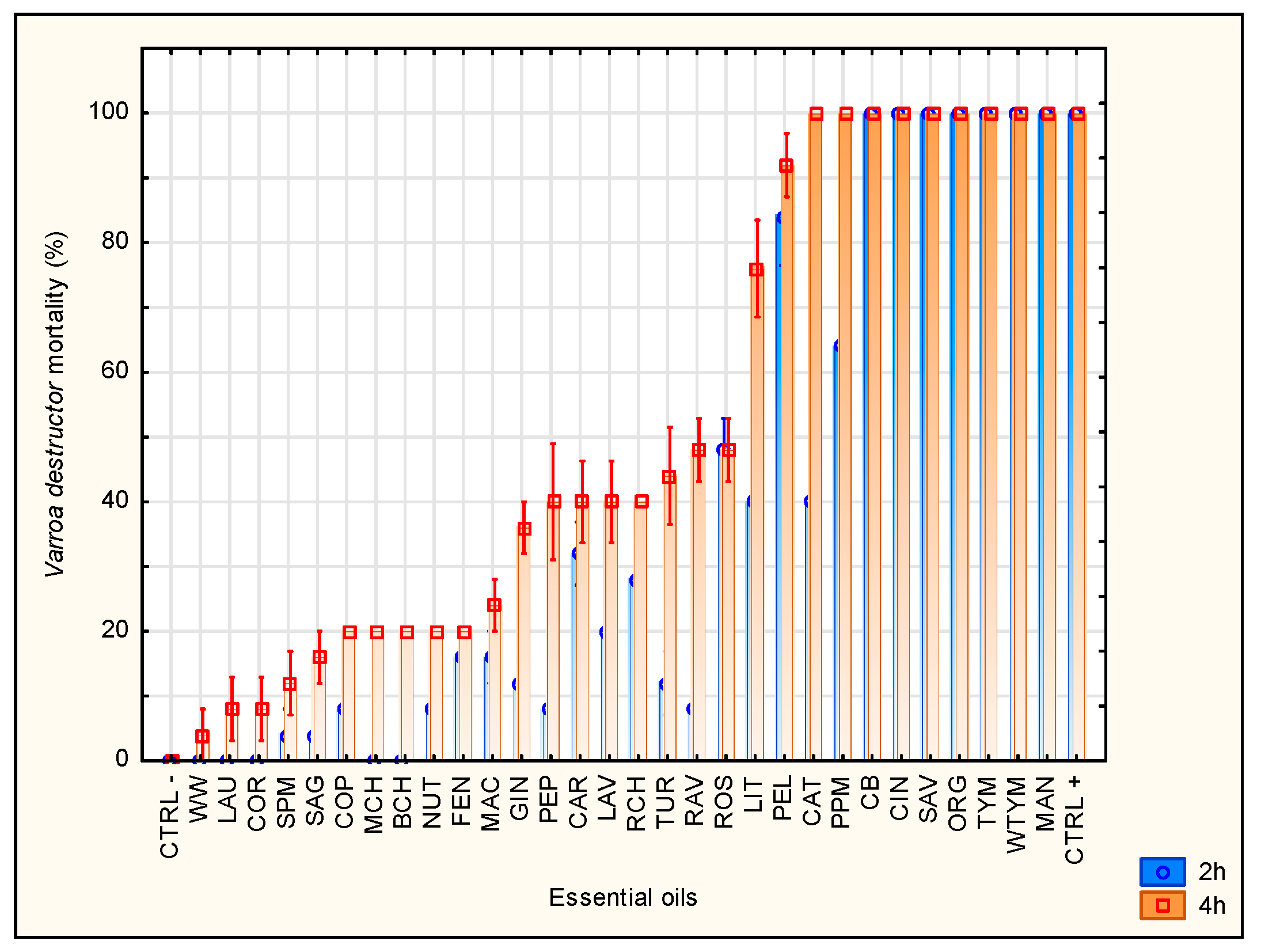
3.1. Screening of Essential Oils for Their Acaricidal Activity
3.2. Complete Exposure Bioassay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Sammataro, D.; Gerson, U.; Needham, G. Parasitic mites of honey bees: Life history, implications, and impact. Annu. Rev. Entomol. 2000, 45, 519–548. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [Green Version]
- Bowen-Walker, P.L.; Gunn, A. The effect of the ectoparasitic mite, Varroa destructor on adult worker honeybee (Apis mellifera) emergence weights, water, protein, carbohydrate, and lipid levels. Entomol. Exp. Appl. 2001, 101, 207–217. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can varroa explain part of the colony losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Bowen-Walker, P.; Martin, S.; Gunn, A. The transmission of deformed wing virus between honeybees (Apis mellifera L.) by the ectoparasitic mite Varroa jacobsoni Oud. J. Invertebr. Pathol. 1999, 73, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhao, Y.; Hammond, J.; Hsu, H.; Evans, J.; Feldlaufer, M. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. J. Invertebr. Pathol. 2004, 87, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Di Prisco, G.; Pennacchio, F.; Caprio, E.; Boncristiani, H.F., Jr.; Evans, J.D.; Chen, Y. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera. J. Gen. Virol. 2011, 92, 151–155. [Google Scholar] [CrossRef]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Shen, M.; Cui, L.; Ostiguy, N.; Cox-Foster, D. Intricate transmission routes and interactions between picorna-like viruses (Kashmir Bee Virus and Sacbrood Virus) with the honeybee host and the parasitic varroa mite. J. Gen. Virol. 2005, 86, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E. Honey bee pathology: Current threats to honey bees and beekeeping. Appl. Microbiol. Biotechnol. 2010, 87, 87–97. [Google Scholar] [CrossRef]
- Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors That may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar]
- Annoscia, D.; Del Piccolo, F.; Nazzi, F. How does the mite Varroa destructor kill the honeybee Apis mellifera? Alteration of cuticular hydrcarbons and water loss in infested honeybees. J. Insect Physiol. 2012, 58, 1548–1555. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J. Ontogenesis of the mite Varroa jacobsoni Oud. in worker brood of the honeybee Apis mellifera L. under natural conditions. Exp. Appl. Acarol. 1994, 18, 87–100. [Google Scholar] [CrossRef]
- Murilhas, A.M. Varroa destructor infestation impact on Apis mellifera Carnica capped worker brood production, bee population and honey storage in a Mediterranean climate. Apidologie 2002, 33, 271–281. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Hood, W.M. Effects of delayed acaricide treatment in honey bee colonies parasitized by Varroa jacobsoni and a late-season treatment threshold for the South-Eastern USA. J. Apic. Res. 1997, 36, 125–132. [Google Scholar] [CrossRef]
- Meikle, W.G.; Sammataro, D.; Neumann, P.; Pflugfelder, J. Challenges for developing pathogen-based biopesticides against Varroa destructor (Mesostigmata: Varroidae). Apidologie 2012, 43, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Calderone, N.W.; Shimanuki, H.; Allen-Wardell, G. An in vitro evaluation of botanical compounds for the control of the honeybee pathogens Bacillus larvae and Ascosphaera apis, and the secondary invader B. Alvei. J. Essent. Oil Res. 1994, 6, 279–287. [Google Scholar] [CrossRef]
- Cakmak, I.; Aydin, L.; Gulegen, E.; Wells, H. Varroa (Varroa destructor) and tracheal mite (Acarapis woodi) incidence in the Republic of Turkey. J. Apic. Res. 2003, 42, 57–60. [Google Scholar] [CrossRef]
- Price, K.L.; Lummis, S.C. An atypical residue in the pore of Varroa destructor GABA-activated RDL receptors affects picrotoxin block and thymol modulation. Insect Biochem. Mol. Biol. 2014, 55, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinkevich, F.D. Detection of amitraz resistance and reduced treatment efficacy in the varroa mite, Varroa destructor, within commercial beekeeping operations. PLoS ONE 2020, 15, e0227264. [Google Scholar] [CrossRef] [Green Version]
- Wallner, K. Varroacides and their residues in bee products. Apidologie 1999, 30, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Kanga, L.H.; Jones, W.A.; Gracia, C. Efficacy of strips coated with Metarhizium anisopliae for control of Varroa destructor (Acari: Varroidae) in honey bee colonies in Texas and Florida. Exp. Appl. Acarol. 2006, 40, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Milani, N. The resistance of Varroa jacobsoni Oud. to acaricides. Apidologie 1999, 30, 229–234. [Google Scholar] [CrossRef]
- Akyol, E.; Özkök, D. The use of organic acids for varroa (Varroa destructor) control. Uludag Bee J. 2005, 5, 167–174. [Google Scholar]
- Aydın, L. Control of honey bee diseases and pets in autumn. Uludag Bee J. 2005, 5, 159–161. [Google Scholar]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Emsen, B.; Dodoloğlu, A.; Genç, F. Natural compounds for the control of Varroa destructor in honey bees (Apis mellifera L.) and an economic assessment of these products. Mellifera 2010, 10, 32–36. [Google Scholar]
- Bahreini, R. Comparison of two methods of applying oxalic acid for control of varroa. J. Apic. Res. 2003, 42, 82–83. [Google Scholar] [CrossRef]
- Hatjina, F.; Haristos, L. Indirect effects of oxalic acid administered by trickling method on honey bee brood. J. Apic. Res. 2005, 44, 172–174. [Google Scholar] [CrossRef]
- Gregorc, A.; Škerl, M.I.S. Toxicological and immunohistochemical testing of honeybees after oxalic acid and rotenone treatments. Apidologie 2007, 38, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Hernández, R.M.; Pascual, M.H.; Pérez, J.; del Nozal Nalda, M.J.; Meana, A. Short term negative effect of oxalic acid in “Apis mellifera Iberiensis”. Span. J. Agric. Res. 2007, 5, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Silva-Zacarin, E.C.; Gregorc, A.; de Moraes, R.L.S. In situ localization of heat-shock proteins and cell death labelling in the salivary gland of acaricide-treated honeybee larvae. Apidologie 2006, 37, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Underwood, R.; Currie, R. Use of formic acid to control varroa and tracheal mites in indoor overwintering facilities. Am. Bee J. 2003, 143, 323. [Google Scholar]
- Higes, M.; Meana, A.; Suárez, M.; Llorente, J. Negative long-term effects on bee colonies treated with oxalic acid against Varroa jacobsoni Oud. Apidologie 1999, 30, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Borsuk, G.; Olszewski, K.; Paleolog, J.; Strachecka, A.; Gryzińska, M. The effect of different varroacides on the acidity of winter stores and honey stores. Ann. UMCS Zootech. 2012, 30, 11–16. [Google Scholar] [CrossRef]
- Conti, B.; Bocchino, R.; Cosci, F.; Ascrizzi, R.; Flamini, G.; Bedini, S. Essential oils against Varroa destructor: A soft way to fight the parasitic mite of Apis mellifera. J. Apic. Res. 2020, 59, 774–782. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A. Chemistry and bioactivity of essential oils. Lipids Essent. Oils Antimicrob. Agents 2011, 25, 203–238. [Google Scholar]
- Athanassiou, C.G.; Rani, P.U.; Kavallieratos, N.G. The use of plant extracts for stored product protection. In Advances in Plant Biopesticides; Springer: Berlin/Heidelberg, Germany, 2014; pp. 131–147. [Google Scholar]
- Bedini, S.; Flamini, G.; Girardi, J.; Cosci, F.; Conti, B. Not just for beer: Evaluation of spent hops (Humulus lupulus L.) as a source of eco-friendly repellents for insect pests of stored foods. J. Pest Sci. 2015, 88, 583–592. [Google Scholar] [CrossRef]
- Bedini, S.; Cosci, F.; Tani, C.; Pierattini, E.C.; Venturi, F.; Lucchi, A.; Ioriatti, C.; Ascrizzi, R.; Flamini, G.; Ferroni, G. Essential oils as post-harvest crop protectants against the fruit fly Drosophila suzukii: Bioactivity and organoleptic profile. Insects 2020, 11, 508. [Google Scholar] [CrossRef]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Echeverria, M.C.; Guidi, L.; Landi, M.; Lucchi, A.; Conti, B. Artemisia spp. essential oils against the disease-carrying blowfly Calliphora vomitoria. Parasites Vectors 2017, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.; Zayed, M. Bioactivity impact of essential oils Allium sativum L. and Citrus reticulata L. against stored product insects Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.). J. Plant Prot. Pathol. 2021, 12, 465–471. [Google Scholar]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef] [Green Version]
- Bedini, S.; Bougherra, H.H.; Flamini, G.; Cosci, F.; Belhamel, K.; Ascrizzi, R.; Conti, B. Repellency of anethole-and estragole-type fennel essential oils against stored grain pests: The different twins. Bull. Insectol. 2016, 69, 149–157. [Google Scholar]
- Conti, B.; Benelli, G.; Leonardi, M.; Afifi, F.U.; Cervelli, C.; Profeti, R.; Pistelli, L.; Canale, A. Repellent effect of Salvia dorisiana, S. longifolia, and S. sclarea (Lamiaceae) essential oils against the mosquito Aedes albopictus Skuse (Diptera: Culicidae). Parasitol. Res. 2012, 111, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.C.; da Silva, T.K.M.; Silva, F.L.; Barbosa-Filho, J.M.; Marques, M.O.M.; Santos, R.L.C.; de Holanda Cavalcanti, S.C.; de Sousa, D.P. Larvicidal activity of Mentha x villosa Hudson essential oil, rotundifolone and derivatives. Chemosphere 2014, 104, 37–43. [Google Scholar] [CrossRef]
- Benelli, G.; Bedini, S.; Cosci, F.; Toniolo, C.; Conti, B.; Nicoletti, M. Larvicidal and ovideterrent properties of neem oil and fractions against the filariasis vector Aedes albopictus (Diptera: Culicidae): A bioactivity survey across production sites. Parasitol. Res. 2015, 114, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Èntomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Rajendran, S.; Sriranjini, V. Plant products as fumigants for stored-product insect control. J. Stored Prod. Res. 2008, 44, 126–135. [Google Scholar] [CrossRef]
- Eguaras, M.J.; Fuselli, S.; Gende, L.; Fritz, R.; Ruffinengo, S.R.; Clemente, G.; Gonzalez, A.; Bailac, P.N.; Ponzi, M.I. An in vitro evaluation of Tagetes minuta essential oil for the control of the honeybee pathogens Paenibacillus larvae and Ascosphaera apis, and the parasitic mite Varroa destructor. J. Essent. Oil Res. 2005, 17, 336–340. [Google Scholar] [CrossRef]
- Imdorf, A.; Bogdanov, S.; Ochoa, R.I.; Calderone, N.W. Use of essential oils for the control of Varroa jacobsoni Oud. in honey bee colonies. Apidologie 1999, 30, 209–228. [Google Scholar] [CrossRef] [Green Version]
- Gashout, H.A.; Guzmán-Novoa, E. Acute toxicity of essential oils and other natural compounds to the parasitic mite, Varroa destructor, and to larval and adult worker honey bees (Apis mellifera L.). J. Apic. Res. 2009, 48, 263–269. [Google Scholar] [CrossRef]
- Ramzi, H.; Ismaili, M.R.; Aberchane, M.; Zaanoun, S. Chemical characterization and acaricidal activity of Thymus satureioides C. & B. and Origanum elongatum E. & M. (Lamiaceae) essential oils against Varroa destructor Anderson & Trueman (Acari: Varroidae). Ind. Crops Prod. 2017, 108, 201–207. [Google Scholar]
- Lindberg, C.M.; Melathopoulos, A.P.; Winston, M.L. Laboratory evaluation of miticides to csontrol Varroa jacobsoni (Acari: Varroidae), a honey bee (Hymenoptera: Apidae) parasite. J. Econ. Entomol. 2000, 93, 189–198. [Google Scholar] [CrossRef]
- Adamczyk, S.; Lázaro, R.; Pérez-Arquillué, C.; Conchello, P.; Herrera, A. Evaluation of residues of essential oil components in honey after different anti-varroa treatments. J. Agric. Food Chem. 2005, 53, 10085–10090. [Google Scholar] [CrossRef] [PubMed]
- Ariana, A.; Ebadi, R.; Tahmasebi, G. Laboratory evaluation of some plant essences to control Varroa destructor (Acari: Varroidae). Exp. Appl. Acarol. 2002, 27, 319–327. [Google Scholar] [CrossRef]
- Aliano, N.P.; Ellis, M.D.; Siegfried, B.D. Acute contact toxicity of oxalic acid to Varroa destructor (Acari: Varroidae) and their Apis mellifera (Hymenoptera: Apidae) hosts in laboratory bioassays. J. Econ. Entomol. 2006, 99, 1579–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffinengo, S.; Eguaras, M.; Floris, I.; Faverin, C.; Bailac, P.; Ponzi, M. LD50 and repellent effects of essential oils from Argentinian wild plant species on Varroa destructor. J. Econ. Entomol. 2005, 98, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Damiani, N.; Gende, L.B.; Bailac, P.; Marcangeli, J.A.; Eguaras, M.J. Acaricidal and insecticidal activity of essential oils on Varroa destructor (Acari: Varroidae) and Apis mellifera (Hymenoptera: Apidae). Parasitol. Res. 2009, 106, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Tihelka, E. Effects of synthetic and organic acaricides on honey bee health: A review. Slov. Veter.-Res. 2018, 55, 114–140. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, G.; Vidau, C.; Ferdy, J.; Tabart, J.; Vetillard, A. Lethal and sub-lethal effects of thymol on honeybee (Apis mellifera) larvae reared in vitro. Pest Manag. Sci. 2014, 70, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.; Underwood, R.; Schwarz, R.; Evans, J.D.; Pettis, J. Direct effect of acaricides on pathogen loads and gene expression levels in honey bees Apis mellifera. J. Insect Physiol. 2012, 58, 613–620. [Google Scholar] [CrossRef]
- Bogdanov, S.; Imdorf, A.; Kilchenmann, V. Residues in wax and honey after Apilife VAR® treatment. Apidologie 1998, 29, 513–524. [Google Scholar] [CrossRef] [Green Version]
- Floris, I.; Satta, A.; Cabras, P.; Garau, V.L.; Angioni, A. Comparison between two thymol formulations in the control of Varroa destructor: Effectiveness, persistence, and residues. J. Econ. Entomol. 2004, 97, 187–191. [Google Scholar] [CrossRef]
- Tananaki, C.; Goras, G.; Huggett, N.; Karazafiris, E.; Dimou, M.; Thrasyvoulou, A. Evaluation of the impact of Exomite ProTM on varroa mite (Varroa destructor) populations and honeybee (Apis mellifera) colonies: Efficacy, side effects and residues. Parasitol. Res. 2014, 113, 1251–1259. [Google Scholar] [CrossRef]
- Ares, A.M.; Nozal, M.J.; Bernal, J.L.; Bernal, J. Simultaneous determination of carvacrol and thymol in bee pollen by using a simple and efficient solvent extraction method and gas chromatography-mass spectrometry. J. Pharm. Biomed. Anal. 2020, 181, 113124. [Google Scholar] [CrossRef]
- Bergougnoux, M.; Treilhou, M.; Armengaud, C. Exposure to thymol decreased phototactic behaviour in the honeybee (Apis mellifera) in laboratory conditions. Apidologie 2013, 44, 82–89. [Google Scholar] [CrossRef]
- Colin, T.; Lim, M.Y.; Quarrell, S.R.; Allen, G.R.; Barron, A.B. Effects of thymol on European honey bee hygienic behaviour. Apidologie 2019, 50, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Gunes, N.; Aydın, L.; Belenli, D.; Hranitz, J.M.; Mengilig, S.; Selova, S. Stress responses of honey bees to organic acid and essential oil treatments against varroa mites. J. Apic. Res. 2017, 56, 175–181. [Google Scholar] [CrossRef]
- Jeong, E.-Y.; Jeon, J.-H.; Kim, H.-W.; Kim, M.-G.; Lee, H.-S. Antimicrobial activity of leptospermone and its derivatives against human intestinal bacteria. Food Chem. 2009, 115, 1401–1404. [Google Scholar] [CrossRef]
- Jeong, E.; Kim, M.; Lee, H. Acaricidal activity of triketone analogues derived from Leptospermum scoparium oil against house-dust and stored-food mites. Pest Manag. Sci. 2009, 65, 327–331. [Google Scholar] [CrossRef]
- Mattila, H.; Otis, G.; Daley, J.; Schulz, T. Trials of apiguard, a thymol-based miticide part 2. Non-target effects on honey bees. Am. Bee J. 2000, 140, 68–70. [Google Scholar]
- Zambonelli, A.; D’Aulerio, A.Z.; Severi, A.; Benvenuti, S.; Maggi, L.; Bianchi, A. Chemical composition and fungicidal activity of commercial essential oils of Thymus vulgaris L. J. Essent. Oil Res. 2004, 16, 69–74. [Google Scholar] [CrossRef]
- Nazer, I.; Al-Abbadi, A. Control of varroa mite (Varroa destructor) on honeybees by aromatic oils and plant materials. J. Agric. Mar. Sci. [JAMS] 2003, 8, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Thielmann, J.; Muranyi, P.; Kazman, P. Screening essential oils for their antimicrobial activities against the foodborne pathogenic bacteria Escherichia coli and Staphylococcus aureus. Heliyon 2019, 5, e01860. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Khan, K.A.; Alqarni, A.S.; Kaur, M.; Al-Waili, N. In vitro evaluation of the effects of some plant essential oils on Ascosphaera apis, the causative agent of chalkbrood disease. Saudi J. Biol. Sci. 2017, 24, 1001–1006. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Al-Waili, N.; Nuru, A.; Sharma, D.; Khan, K.A.; Kaur, M.; Omer, M. In vitro evaluation of the effects of some plant essential oils on Paenibacillus larvae, the causative agent of American foulbrood. Biotechnol. Biotechnol. Equip. 2016, 30, 49–55. [Google Scholar] [CrossRef]
- Choi, Y.; Woo, S.; Hong, I.; Han, S.; Byoun, G.; Thapa, R.; Lee, M. Effects of limonene composition for controlling of the Varroa destructor and Tropilaelaps clareae in Apis mellifera Hives. Korean J. Apic. 2012, 27, 137–142. [Google Scholar]
- van Vuuren, S.; Viljoen, A.M. Antimicrobial activity of limonene enantiomers and 1, 8-cineole alone and in combination. Flavour Fragr. J. 2007, 22, 540–544. [Google Scholar] [CrossRef]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and antifungal activity of aromatic constituents of Essential oils. Microbios 1997, 89, 39–46. [Google Scholar]
- Leite, A.M.; Lima, E.D.O.; De Souza, E.L.; Diniz, M.D.F.F.M.; Trajano, V.N.; De Medeiros, I.A. Inhibitory effect of beta-pinene, alpha-pinene and eugenol on the growth of potential infectious endocarditis causing gram-positive bacteria. Rev. Bras. Ciênc. Farm. 2007, 43, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Nie, Q.; Ren, M.; Feng, H.; Jiang, X.; Zheng, Y.; Liu, M.; Zhang, H.; Xian, M. Metabolic engineering of Escherichia coli for the biosynthesis of alpha-pinene. Biotechnol. Biofuels 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, M.; Chaves, F.; Almeida, C.A.; Bizzo, H.R.; Duarte, R.S.; Campos-Takaki, G.M.; Alviano, C.S.; Alviano, D.S. Antioxidant and antimicrobial activities of 7-hydroxy-calamenene-rich essential oils from Croton cajucara Benth. Molecules 2013, 18, 1128–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.-T.; Yang, T.-S. Antimicrobial impact of the components of essential oil of Litsea cubeba from Taiwan and antimicrobial Activity of the oil in food systems. Int. J. Food Microbiol. 2012, 156, 68–75. [Google Scholar] [CrossRef]
- Onawunmi, G.O. Evaluation of the antimicrobial activity of citral. Lett. Appl. Microbiol. 1989, 9, 105–108. [Google Scholar] [CrossRef]
- Can Baser, K. Biological and pharmacological activities of carvacrol and carvacrol bearing essential oils. Curr. Pharm. Des. 2008, 14, 3106–3119. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.A.; Stephens, J.C. A review of cinnamaldehyde and its derivatives as antibacterial agents. Fitoterapia 2019, 139, 104405. [Google Scholar] [CrossRef]

{kind=link}
| English Name | Abbreviation | Latin Name | Part of Plant |
|---|---|---|---|
| Black pepper | PEP | Piper nigrum | berry |
| Blue chamomile | BCH | Matricaria chamomilla | flower |
| Carrot | CAT | Daucus carota | seeds |
| Cinnamon | CIN | Cinnamomum zeylanicum | bark |
| Clove Bud | CB | Eugenia caryophyllata | leaves, buds, and twigs |
| Copaiba | COP | Copaifera reticulata | resin |
| Coriander | COR | Coriandrum sativum | seeds |
| Fennel | FEN | Foeniculum vulgare | seeds |
| Ginger | GIN | Zingiber officinale | rhizome |
| Green cardamom | CAR | Elettaria cardamomum | seeds |
| Laurel | LAU | Laurus nobilis | leaves |
| Lavender | LAV | Lavandula angustifolia | flowering herb |
| Litsea | LIT | Litsea cubeba | fruits |
| Mace | MAC | Myristica fragrans | flower |
| Manuka | MAN | Leptospermum scoparium | leaves and twigs |
| Maroc chamomile | MCH | Ormenis multicaulis | herb |
| Nutmeg | NUT | Myristica fragrans | seeds |
| Oregano | ORG | Origanum vulgare | herb |
| Pelargonium | PEL | Pelargonium graveolens | leaves and flowers |
| Peppermint | PPM | Mentha piperita | herb |
| Ravensara | RAV | Ravensara aromatica | leaves and twigs |
| Roman chamomile | RCH | Anthemis nobilis | flower |
| Rosemary | ROS | Rosmarinus officinalis | herb |
| Sage | SAG | Salvia officinalis | leaves |
| Savory | SAV | Satureja montana | herb |
| Spearmint | SPM | Mentha spicata crispa | flowering herb |
| Thyme | TYM | Thymus vulgaris | herb |
| Turmeric | TUR | Curcuma longa | root |
| Wild thyme | WTYM | Thymus serpyllum | herb |
| Wormwood | WW | Artemisa absinthium | herb |
| EO | Species | 4 h | 24 h | 48 h | 72 h | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LC50 | 95% CL | LC50 | 95% CL | LC50 | 95% CL | LC50 | 95% CL | ||||||
| THM | V. destructor | 1.505 | 1.180 | 1.937 | 0.834 | 0.629 | 1.052 | 0.660 | 0.475 | 0.846 | 0.660 | 0.475 | 0.846 |
| A. mellifera | 6.181 | 5.074 | 7.847 | 4.090 | 3.189 | 6.759 | 2.427 | 2.097 | 2.871 | 2.112 | 1.940 | 2.320 | |
| Selectivity ratio | 4.107 | 4.903 | 3.675 | 3.198 | |||||||||
| CAT | V. destructor | 10.449 | 6.806 | 34.882 | 4.167 | 2.457 | 6.630 | 3.276 | 1.930 | 4.590 | 2.539 | 1.187 | 3.653 |
| A. mellifera | 18.607 | 13.845 | 64.136 | 13.048 | 9.588 | 27.137 | 11.557 | 8.855 | 19.447 | 11.557 | 8.855 | 19.447 | |
| Selectivity ratio | 1.781 | 3.131 | 3.527 | 4.552 | |||||||||
| PPM | V. destructor | 8.121 | 6.159 | 13.576 | 2.512 | 1.430 | 3.578 | 1.732 | 0.499 | 2.806 | 1.066 | 0.011 | 2.197 |
| A. mellifera | 12.951 | 11.259 | 14.994 | 10.759 | 9.483 | 12.156 | 10.285 | 9.109 | 11.568 | 10.285 | 9.109 | 11.568 | |
| Selectivity ratio | 1.595 | 4.283 | 5.939 | 9.651 | |||||||||
| SAV | V. destructor | 3.825 | 3.165 | 4.918 | 2.008 | 1.323 | 2.754 | 1.459 | 0.626 | 2.075 | 1.364 | 0.417 | 1.996 |
| A. mellifera | 11.657 | 9.247 | 16.335 | 5.786 | 4.607 | 7.218 | 5.275 | 4.273 | 6.467 | 4.621 | 3.884 | 5.897 | |
| Selectivity ratio | 3.048 | 2.881 | 3.615 | 3.386 | |||||||||
| WTYM | V. destructor | 8.185 | 5.218 | 22.390 | 2.549 | 1.495 | 8.207 | 2.013 | 0.926 | 7.327 | 1.861 | 0.825 | 5.487 |
| A. mellifera | 9.074 | 8.106 | 10.780 | 7.517 | 6.606 | 8.265 | 6.512 | 5.958 | 7.494 | 6.250 | 5.603 | 6.897 | |
| Selectivity ratio | 1.109 | 2.949 | 3.236 | 3.358 | |||||||||
| ORG | V. destructor | 3.517 | 2.339 | 7.322 | 0.879 | 0.638 | 1.302 | 0.577 | 0.280 | 0.924 | 0.577 | 0.280 | 0.924 |
| A. mellifera | 6.982 | 6.136 | 7.889 | 3.362 | 2.997 | 3.803 | 3.362 | 2.997 | 3.803 | 3.362 | 2.997 | 3.803 | |
| Selectivity ratio | 1.985 | 3.827 | 5.830 | 5.830 | |||||||||
| PEL | V. destructor | 2.798 | 0.113 | 4.758 | 2.291 | 0.247 | 3.825 | 2.402 | 0.804 | 3.532 | 2.272 | 0.788 | 3.311 |
| A. mellifera | 17.122 | 13.427 | 27.935 | 12.401 | 10.159 | 17.209 | 9.479 | 8.132 | 11.201 | 9.479 | 8.132 | 11.201 | |
| Selectivity ratio | 6.120 | 5.413 | 3.945 | 4.171 | |||||||||
| MAN | V. destructor | 1.262 | 0.848 | 3.192 | 1.029 | 0.558 | 2.880 | 0.265 | 0.020 | 0.540 | 0.158 | 0.011 | 0.497 |
| A. mellifera | 1.975 | 1.662 | 2.681 | 1.415 | 1.218 | 1.666 | 1.472 | 1.277 | 1.720 | 1.472 | 1.277 | 1.720 | |
| Selectivity ratio | 1.565 | 1.375 | 5.551 | 9.333 | |||||||||
| LIT | V. destructor | 4.801 | 3.522 | 7.436 | 2.716 | 1.322 | 4.311 | 2.116 | 0.243 | 3.761 | 1.807 | 0.243 | 2.989 |
| A. mellifera | 11.660 | 9.524 | 15.222 | 11.590 | 8.994 | 20.096 | 9.207 | 7.721 | 12.115 | 9.678 | 7.255 | 18.278 | |
| Selectivity ratio | 2.429 | 4.267 | 4.352 | 5.354 | |||||||||
| TYM | V. destructor | 1.279 | 0.985 | 1.613 | 0.678 | 0.314 | 0.940 | 0.678 | 0.314 | 0.940 | 0.587 | 0.202 | 0.851 |
| A. mellifera | 8.759 | 6.684 | 14.553 | 3.887 | 3.295 | 4.837 | 3.113 | 2.696 | 3.763 | 2.677 | 2.418 | 2.982 | |
| Selectivity ratio | 6.848 | 5.731 | 4.590 | 4.557 | |||||||||
| CB | V. destructor | 2.337 | 1.829 | 2.962 | 1.690 | 1.237 | 2.143 | 1.490 | 1.207 | 1.776 | 1.490 | 1.207 | 1.776 |
| A. mellifera | 5.965 | 4.620 | 10.868 | 4.860 | 4.023 | 6.546 | 3.305 | 2.790 | 4.179 | 3.305 | 2.790 | 4.179 | |
| Selectivity ratio | 2.553 | 2.875 | 2.218 | 2.218 | |||||||||
| CIN | V. destructor | 4.321 | 3.163 | 5.979 | 2.820 | 1.577 | 4.002 | 2.529 | 1.370 | 3.590 | 1.543 | 0.829 | 2.484 |
| A. mellifera | 10.635 | 9.559 | 11.972 | 7.488 | 6.680 | 8.408 | 7.488 | 6.680 | 8.408 | 7.007 | 5.835 | 8.664 | |
| Selectivity ratio | 2.461 | 2.655 | 2.960 | 4.542 | |||||||||
| EO | Main Components and Their Quantity (%) | ||||
|---|---|---|---|---|---|
| Carrot | Ceratol 30.28 | α-Pinen 15.462 | Sabinen 10.22 | β- Caryophyllen 8.31 | β-bisabolen 5.63 |
| Peppermint | Limonen 38.02 | Menthol 16.41 | α-Pinen 15.92 | β-Pinen 11.46 | Menthon 5.65 |
| Savory | Carvacrol 41.67 | ɣ-Terpinen 35.82 | p-Cymen 11.73 | - | - |
| Wild thyme | Thymol 16.33 | Carvacrol 15.38 | p-Cymen 15.01 | Geraniol 10.62 | ɣ-Terpinen 10.30 |
| Oregano | Carvacrol 73.50 | p-Cymen 6.97 | ɣ-Terpinen 6.02 | - | - |
| Pelargonium | Citronellol 33.51 | Geraniol 15.36 | Citronellylformiat 7.81 | Isomenthon 5.61 | 10-epi-g-Eudesmol 5.37 |
| Manuka | Calamenene 17.92 | Leptospermone 16.02 | Flaveson 5.95 | α-Selinene 4.62 | - |
| Litsea | Citral A 39.03 | Citral B 29.35 | Limonen 13.74 | - | - |
| Thyme | Thymol 40.96 | p-Cymen 16.76 | - | - | - |
| Clove Bud | Eugenol 86.62 | β-Caryophyllen 10.21 | - | - | - |
| Cinnamon | trans-Cinnamaldehyde 77.69 | Eugenol 7.50 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hýbl, M.; Bohatá, A.; Rádsetoulalová, I.; Kopecký, M.; Hoštičková, I.; Vaníčková, A.; Mráz, P. Evaluating the Efficacy of 30 Different Essential Oils against Varroa destructor and Honey Bee Workers (Apis mellifera). Insects 2021, 12, 1045. https://doi.org/10.3390/insects12111045
Hýbl M, Bohatá A, Rádsetoulalová I, Kopecký M, Hoštičková I, Vaníčková A, Mráz P. Evaluating the Efficacy of 30 Different Essential Oils against Varroa destructor and Honey Bee Workers (Apis mellifera). Insects. 2021; 12(11):1045. https://doi.org/10.3390/insects12111045
Chicago/Turabian StyleHýbl, Marian, Andrea Bohatá, Iva Rádsetoulalová, Marek Kopecký, Irena Hoštičková, Alena Vaníčková, and Petr Mráz. 2021. "Evaluating the Efficacy of 30 Different Essential Oils against Varroa destructor and Honey Bee Workers (Apis mellifera)" Insects 12, no. 11: 1045. https://doi.org/10.3390/insects12111045
APA StyleHýbl, M., Bohatá, A., Rádsetoulalová, I., Kopecký, M., Hoštičková, I., Vaníčková, A., & Mráz, P. (2021). Evaluating the Efficacy of 30 Different Essential Oils against Varroa destructor and Honey Bee Workers (Apis mellifera). Insects, 12(11), 1045. https://doi.org/10.3390/insects12111045