
The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Overwintering Study
2.2. Statistical Analysis
2.2.1. Overwintering Assessment
2.2.2. Super-Cooling Point Assessment
3. Results
3.1. Overwintering Results
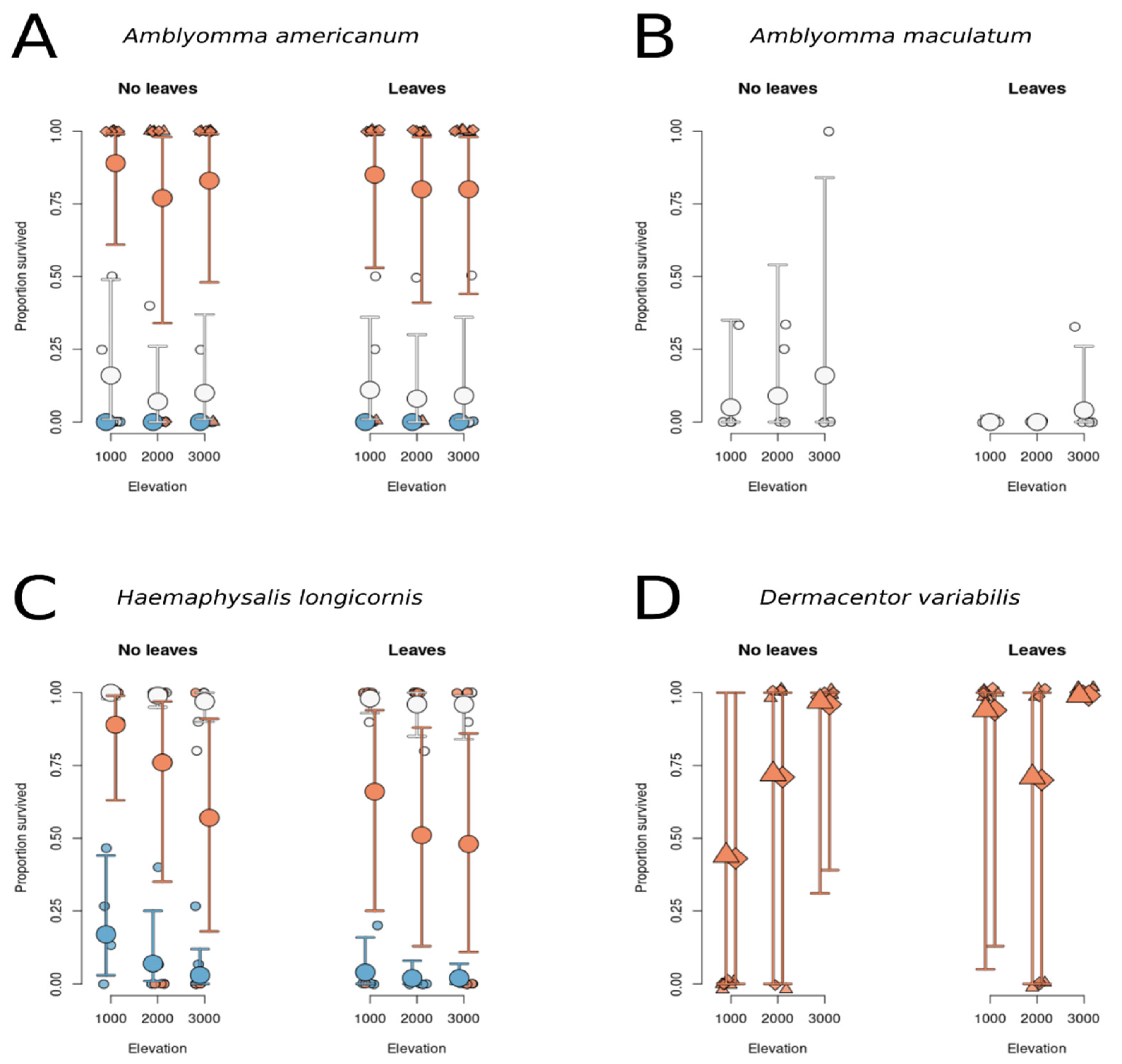
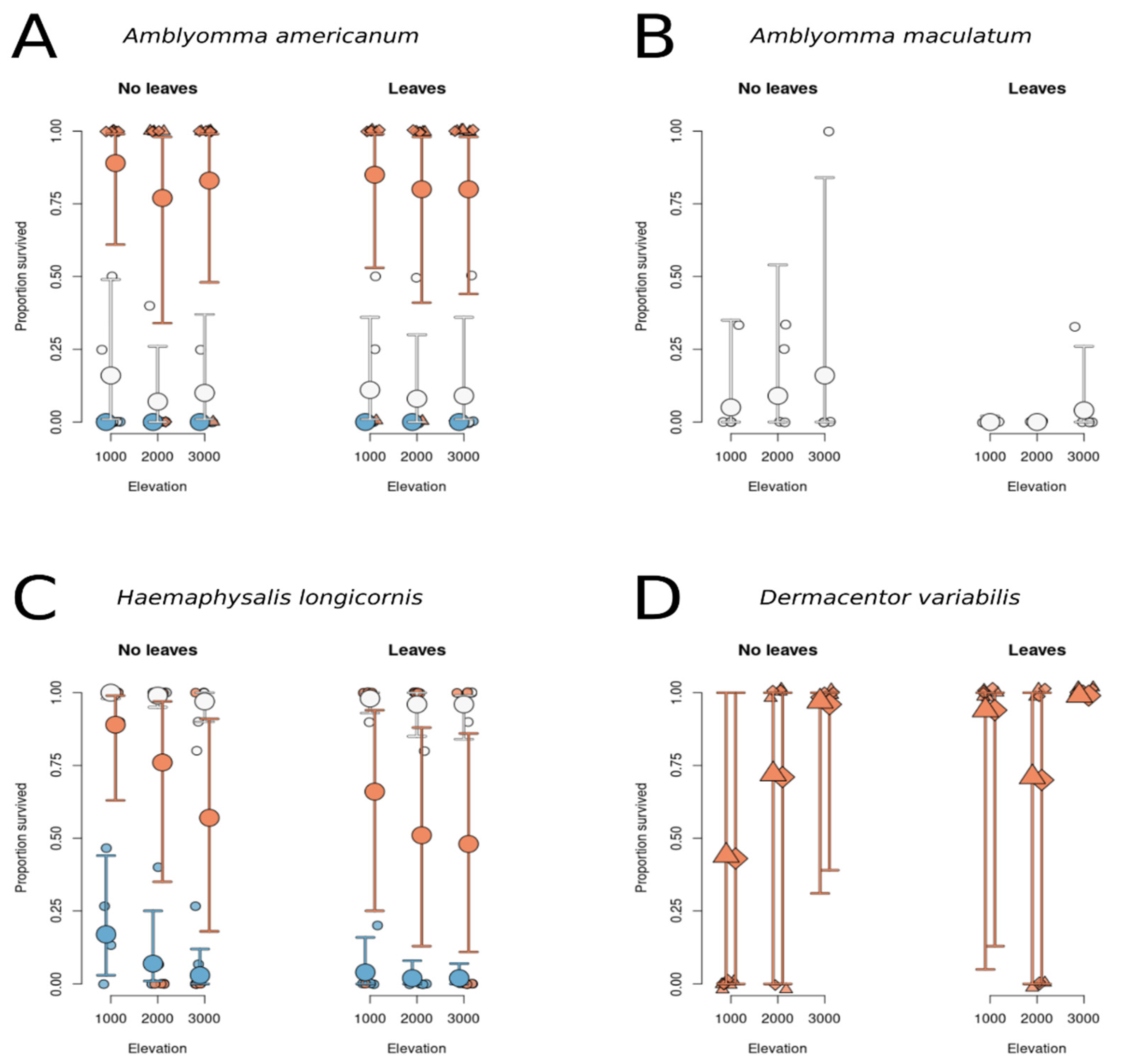
3.1.1. Survival of A. americanum Does Not Depend on Leaf Litter or Elevation but Depends on Life Stage
3.1.2. Survival of H. longicornis Does Not Depend on Leaf Insulation or Elevation but Depends on Life stage
3.1.3. Survival of D. variabilis Is Not Predicted by Elevation or Leaf Litter Treatment
3.2. Temperatures, Relative Humidity, and Precipitation Experienced at Each Elevation
3.3. Super-Cooling Point Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Z.J.; Lu, Y.L.; Yang, X.L.; Chen, J.; Wang, H.; Wang, D.; Liu, J.Z. Cold hardiness and biochemical response to low temperature of the unfed bush tick Haemaphysalis longicornis (Acari: Ixodidae). Parasit Vectors 2014, 7, 346. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, L.R.; Barker, I.K.; Surgeoner, G.A.; McEwen, S.A.; Gillespie, T.J.; Robinson, J.T. Survival and development of Ixodes scapularis (Acari: Ixodidae) under various climatic conditions in Ontario, Canada. J. Med. Entomol. 1995, 32, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.L.; Killilea, M.; Ostfeld, R.S. Overwintering survival of nymphal Ixodes scapularis (Acari: Ixodidae) under natural conditions. J. Med. Entomol. 2012, 49, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Eisen, L.; Ogden, N.H.; Beard, C.B. Linkages of weather and climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), enzootic transmission of Borrelia burgdorferi, and Lyme disease in North America. J. Med. Entomol. 2016, 49, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEnroe, W.D. The effects of snow-cover on an American dog tick, Dermacentor variabilis (Say.) (Acari, Ixodidae) population under a harsh winter environment. Z. Angew. Entomol. 1984, 97, 481–484. [Google Scholar] [CrossRef]
- Linske, M.A.; Stafford, K.C.; Williams, S.C.; Lubelczyk, C.B.; Welch, M.; Henderson, E.F. Impacts of deciduous leaf litter and snow presence on nymphal Ixodes scapularis (Acari: Ixodidae) overwintering survival in coastal New England, USA. Insects 2019, 10, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, H.S.; Zhioua, E. Nymphal survival and habitat distribution of Ixodes scapularis and Amblyomma americanum ticks (Acari: Ixodidae) on Fire Island, New York, USA. Exp. Appl. Acarol. 1996, 20, 533–544. [Google Scholar] [CrossRef]
- Burks, C.S.; Stewart, R.L.; Needham, G.R.; Lee, R.E. The role of direct chilling injury and inoculative freezing in cold tolerance of Amblyomma americanum, Dermacentor variabilis and Ixodes scapularis. Physiol. Entomol. 1996, 21, 44–50. [Google Scholar] [CrossRef]
- Burtis, J.C.; Fahey, T.J.; Yavitt, J.B. Survival and energy use of Ixodes scapularis nymphs throughout their overwintering period. Parasitology 2019, 146, 781–790. [Google Scholar] [CrossRef]
- National Haemaphysalis Longicornis (Asian Longhorned Tick) Situation Report. Available online: https://www.aphis.usda.gov/animal_health/animal_diseases/tick/downloads/longhorned-tick-sitrep.pdf (accessed on 20 August 2021).
- Raghavan, R.K.; Paterson, A.; Cobbs, M.E.; Ganta, R.; Foley, D. Current and future distribution of the Lone Star tick Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLoS ONE 2019, 14, e0209082. [Google Scholar] [CrossRef] [PubMed]
- Nadolny, R.M.; Gaff, H.D. Natural history of Amblyomma maculatum in Virginia. Ticks Tick Borne Dis. 2018, 9, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Oakes, V.J.; Yabsley, M.J.; Schwartz, D.; LeRoith, T.; Bissett, C.; Broaddus, C.; Schlater, J.L.; Todd, S.M.; Boes, K.M.; Brookhart, M.; et al. Theileria orientalis Ikeda Genotype in Cattle, Virginia, USA. Emerg. Infect. Dis. 2019, 25, 1653–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; et al. Distribution of Haemaphysalis longicornis and associated pathogens: Analysis of pooled data from a China field survey and global published data. Lancet Planet 2020, 4, 1653–1659. [Google Scholar] [CrossRef]
- Paddock, C.D.; Goddard, J. The Evolving medical and veterinary importance of the Gulf Coast tick (Acari: Ixodidae). J. Med. Entomol. 2015, 52, 230–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunik, M.E.M.; Galloway, T.D.; Lindsay, L.R. Ability of unfed Dermacentor variabilis (Acari: Ixodidae) to survive a second winter as adults in Manitoba, Canada near the northern limit of their range. J. Med. Entomol. 2015, 52, 138–142. [Google Scholar] [CrossRef]
- Linske, M.A.; Williams, S.C.; Stafford, K.C.; Lubelczyk, C.B.; Henderson, E.F.; Welch, M.; Teel, P.D. Determining effects of winter weather conditions on adult Amblyomma americanum (Acari: Ixodidae) survival in Connecticut and Maine, USA. Insects 2020, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Rosendale, A.J.; Farrow, D.W.; Dunlevy, M.E.; Fieler, A.M.; Benoit, J.B. Cold hardiness and influences of hibernaculum conditions on overwintering survival of American dog tick larvae. Ticks Tick Borne Dis. 2016, 7, 1155–1161. [Google Scholar] [CrossRef]
- Gulf Coast Tick: Amblyomma Maculatum. Available online: https://entnemdept.ufl.edu/creatures/urban/medical/gulf_coast_tick.htm (accessed on 20 August 2021).
- Asian Longhorned Tick: Haemaphysalis Longicornis. Available online: https://entnemdept.ufl.edu/creatures/Livestock/Ticks/Asian_longhorned_tick.html (accessed on 30 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 2 August 2021).
- McElreath, R. Rethinking: Statistical Rethinking. A Bayesian Course with Examples in R and STAN, 2nd ed.; Book Series; Chapman & Hall/CRC: London, UK, 2020. [Google Scholar]
- Kelter, R. Analysis of Bayesian Posterior Significance and Effect Size Indices for the Two-Sample T-Test to Support Reproducible Medical Research. BMC Med. Res. Methodol. 2020, 20, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Cira, T.M.; Venette, R.C.; Agner, J.; Kuhar, T.; Mullins, D.E.; Gabbert, S.E.; Hutchison, W.D. Cold tolerance of Halyomorpha halys (Hemiptera: Pentatomidae) across geographic and temporal scales. Environ. Entomol. 2016, 45, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Climate Data Online: Dataset Discovery. Available online: https://www.ncdc.noaa.gov/cdo-web/datasets (accessed on 22 September 2021).
- Kim, J.; Jung, M.; Kho, J.; Song, H.; Moon, K.; Kim, Y.H.; Lee, D. Characterization of overwintering sites of Haemaphysalis longicornis (Acari: Ixodidae) and tick infection rate with severe fever with thrombocytopenia syndrome virus from eight provinces in South Korea. Ticks Tick Borne Dis. 2020, 11, 101490. [Google Scholar] [CrossRef]
- Minigan, J.W.; Hager, H.H.; Peregrine, A.S.; Newman, J.A. Current and potential future distribution of the American dog tick (Dermacentor variabilis, Say) in North America. Ticks Tick Borne Dis. 2018, 9, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Boorgula, G.D.; Peterson, A.T.; Foley, D.H.; Ganta, R.R.; Raghavan, R.K. Assessing the current and future potential geographic distribution of the American dog tick, Dermacentor variabilis (Say) (Acari: Ixodidae) in North America. PLoS ONE 2020, 15, e0237191. [Google Scholar] [CrossRef] [PubMed]
- Trout Fryxell, R.T.; Moore, J.E.; Collins, M.D.; Kwon, Y.; Jean-Philippe, S.R.; Schaeffer, S.M.; Odoi, A.; Kennedy, M.; Houston, A.E. Habitat and vegetation variables are not enough when predicting tick populations in the southeastern United States. PLoS ONE 2015, 10, e0144092. [Google Scholar] [CrossRef]
- Diyes, C.P.; Dergousoff, S.J.; Yunik, M.E.M.; Chilton, N.B. Reproductive output and larval survival of American dog ticks (Dermacentor variabilis) from a population at the northern distributional limit. Exp. Appl. Carol. 2021, 83, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Climate Change and the 1991–2020 U.S. Climate Normals. Available online: https://www.climate.gov/news-features/understanding-climate/climate-change-and-1991-2020-us-climate-normals (accessed on 29 August 2021).
- Virginia Museum of History & Culture. Available online: https://www.virginiahistory.org/collections-and-resources/virginia-history-explorer/regions-virginia (accessed on 20 August 2021).
- Virginia’s Climate. Available online: https://www.daculaweather.com/climate_reports/ClimateSum_VA.pdf (accessed on 20 August 2021).
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.; Golovljova, I.; Jaenson, T.; Jensen, J.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit Vectors 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Dautel, H.; Estrada-Pena, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef] [PubMed]
- Zajac, Z.; Bartosik, K.; Kulisz, J.; Wozniak, A. Ability of adult Dermacentor reticulatus ticks to overwinter in the temperate climate zone. Biology 2020, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Neelakanta, G.; Sultana, H.; Fish, D.; Anderson, J.F.; Fikrig, E. Anaplasma phagocytophilum induces Ixodes scapularis ticks to express an antifreeze glycoprotein gene that enhances their survival in the cold. J. Clin. Investig. 2010, 120, 3179–3190. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| No Leaves, # of Replicates | No Leaves, Survived/Placed | Leaves, # of Replicates | Leaves, Survived/Placed | |
|---|---|---|---|---|
| Amblyomma americanum | 48 | 26/249 | 48 | 25/253 |
| 1000 ft/305 m | 16 | 10/80 | 16 | 9/86 |
| Larva | 4 | 0/56 | 4 | 0/60 |
| Nymph | 4 | 3/16 | 4 | 3/18 |
| Adult (males) | 4 | 4/4 | 4 | 4/4 |
| Adult (females) | 4 | 3/4 | 4 | 2/4 |
| 2000 ft/610 m | 16 | 8/86 | 16 | 8/84 |
| Larva | 4 | 0/60 | 4 | 0/60 |
| Nymph | 4 | 2/18 | 4 | 2/16 |
| Adult (males) | 4 | 3/4 | 4 | 4/4 |
| Adult (females) | 4 | 3/4 | 4 | 2/4 |
| 3000 ft/914 m | 16 | 8/83 | 16 | 8/83 |
| Larva | 4 | 0/60 | 4 | 0/60 |
| Nymph | 4 | 1/15 | 4 | 2/15 |
| Adult (males) | 4 | 4/4 | 4 | 2/4 |
| Adult (females) | 4 | 3/4 | 4 | 4/4 |
| Amblyomma maculatum | 12 | 6/39 | 12 | 1/38 |
| 1000 ft/305 m | 4 | 1/13 | 4 | 0/13 |
| 2000 ft/610 m | 4 | 2/13 | 4 | 0/13 |
| 3000 ft/914 m | 4 | 3/13 | 4 | 1/12 |
| Haemaphysalis longicornis | 36 | 147/312 | 36 | 127/311 |
| 1000 ft/305 m | 12 | 56/104 | 12 | 44/103 |
| Larva | 4 | 13/60 | 4 | 3/60 |
| Nymph | 4 | 39/40 | 4 | 38/39 |
| Adult | 4 | 4/4 | 4 | 3/4 |
| 2000 ft/610 m | 12 | 48/104 | 12 | 42/104 |
| Larva | 4 | 7/60 | 4 | 0/60 |
| Nymph | 4 | 40/40 | 4 | 38/40 |
| Adult | 4 | 1/4 | 4 | 4/4 |
| 3000 ft/914 m | 12 | 43/104 | 12 | 41/104 |
| Larva | 4 | 5/60 | 4 | 0/60 |
| Nymph | 4 | 37/40 | 4 | 39/40 |
| Adult | 4 | 1/4 | 4 | 2/4 |
| Dermacentor variabilis | 24 | 14/24 | 24 | 20/24 |
| 1000 ft/305 m | 8 | 0/8 | 8 | 8/8 |
| Adult (males) | 4 | 0/4 | 4 | 4/4 |
| Adult (females) | 4 | 0/4 | 4 | 4/4 |
| 2000 ft/610 m | 8 | 6/8 | 8 | 4/8 |
| Adult (males) | 4 | 3/4 | 4 | 2/4 |
| Adult (females) | 4 | 3/4 | 4 | 2/4 |
| 3000 ft/914 m | 8 | 8/8 | 8 | 8/8 |
| Adult (males) | 4 | 4/4 | 4 | 4/4 |
| Adult (females) | 4 | 4/4 | 4 | 4/4 |
| Elevation | Minimum Temp (°C) | Average Temperature (°C) | Maximum Temperature (°C) | Minimum RH | Average RH | Maximum RH |
|---|---|---|---|---|---|---|
| 1000 ft/305 m (Roanoke) | −8.66 | 6.33 | 24.97 | 18.458 | 81.76 | 100 |
| 2000 ft/610 m (Montgomery) | −5.51 | 5.60 | 20.34 | 1.00 | 89.95 | 100 |
| 3000 ft/914 m (Floyd) | −6.96 | 5.97 | 26.09 | 19.44 | 88.85 | 100 |
| Tick Species/Life Stage | Average Super-Cooling Point (°C) | Standard Deviation (SD) | Standard Error (SEM) |
|---|---|---|---|
| Amblyomma americanum Nymph | −23.53 | 1.56 | 0.29 |
| Amblyomma americanum Adult Male | −19.74 | 1.62 | 0.42 |
| Amblyomma americanum Adult Female | −17.30 | 4.38 | 1.13 |
| Haemaphysalis longicornis Nymph | −24.08 | 1.40 | 0.50 |
| Haemaphysalis longicornis Adult Female | −22.64 | 1.44 | 0.38 |
| Dermacentor variabilis Adult Male | −16.04 | 7.21 | 1.86 |
| Dermacentor variabilis Adult Female | −20.16 | 4.91 | 1.23 |
| Amblyomma maculatum Nymph | −22.23 | 2.42 | 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitlow, A.M.; Schürch, R.; Mullins, D.; Eastwood, G. The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region. Insects 2021, 12, 1000. https://doi.org/10.3390/insects12111000
Whitlow AM, Schürch R, Mullins D, Eastwood G. The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region. Insects. 2021; 12(11):1000. https://doi.org/10.3390/insects12111000
Chicago/Turabian StyleWhitlow, Amanda Marie, Roger Schürch, Donald Mullins, and Gillian Eastwood. 2021. "The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region" Insects 12, no. 11: 1000. https://doi.org/10.3390/insects12111000
APA StyleWhitlow, A. M., Schürch, R., Mullins, D., & Eastwood, G. (2021). The Influence of Southwestern Virginia Environmental Conditions on the Potential Ability of Haemaphysalis longicornis, Amblyomma americanum, and Amblyomma maculatum to Overwinter in the Region. Insects, 12(11), 1000. https://doi.org/10.3390/insects12111000