
Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Predator Inoculation Density × Production Period
2.2. Volume of the Rearing Unit
2.3. Statistical Analyses
3. Results
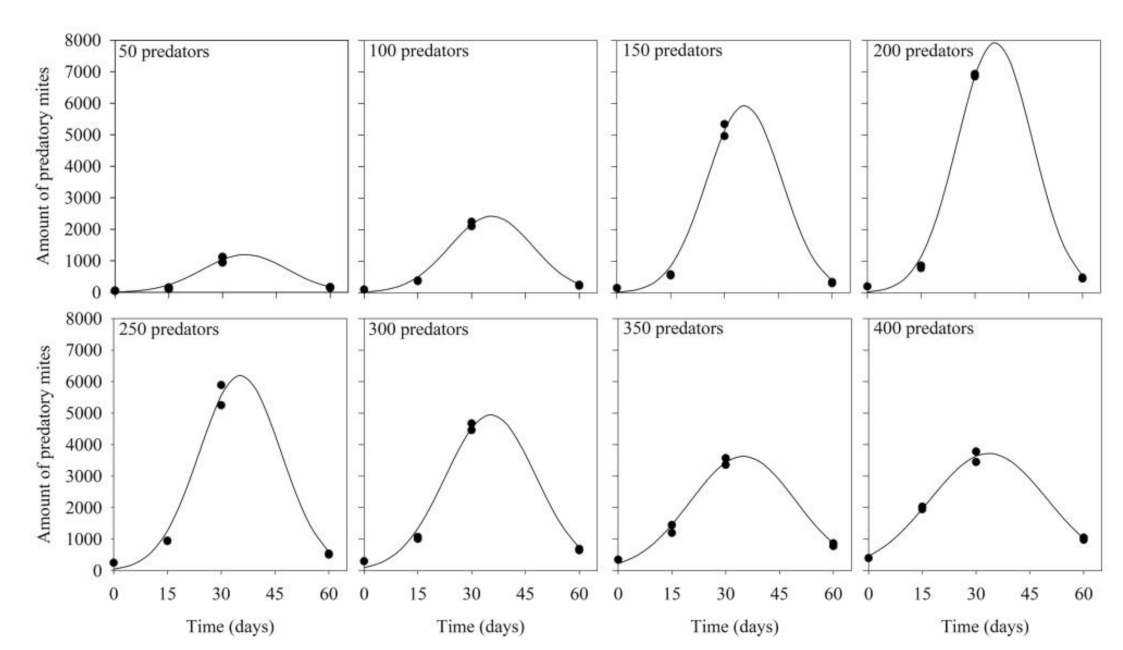
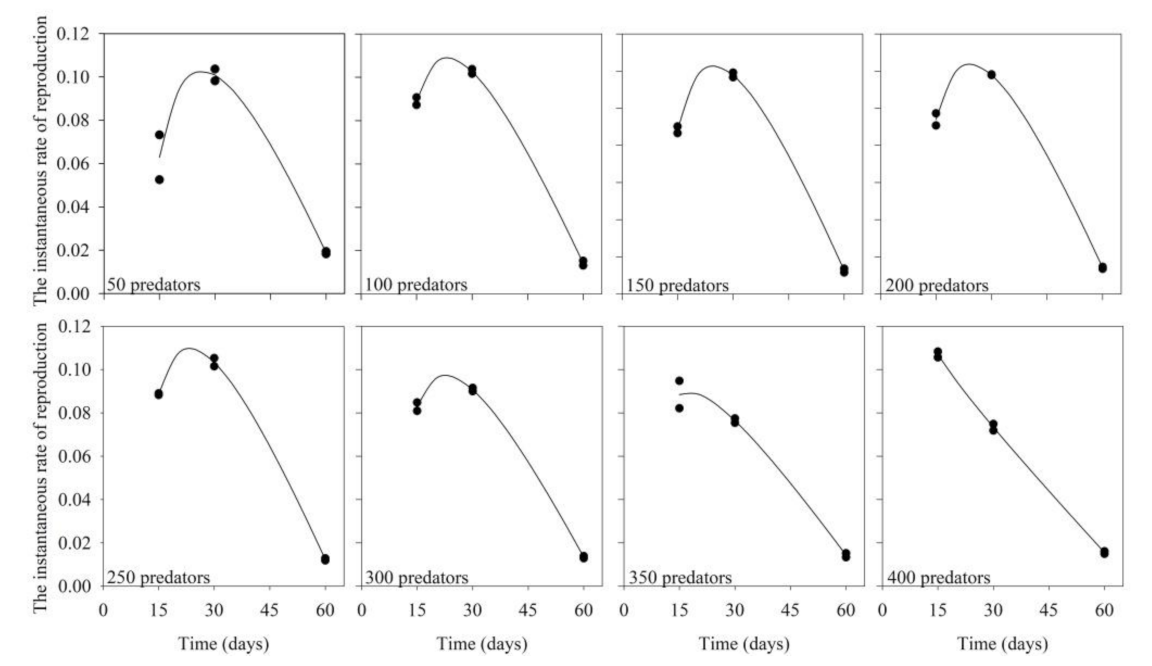
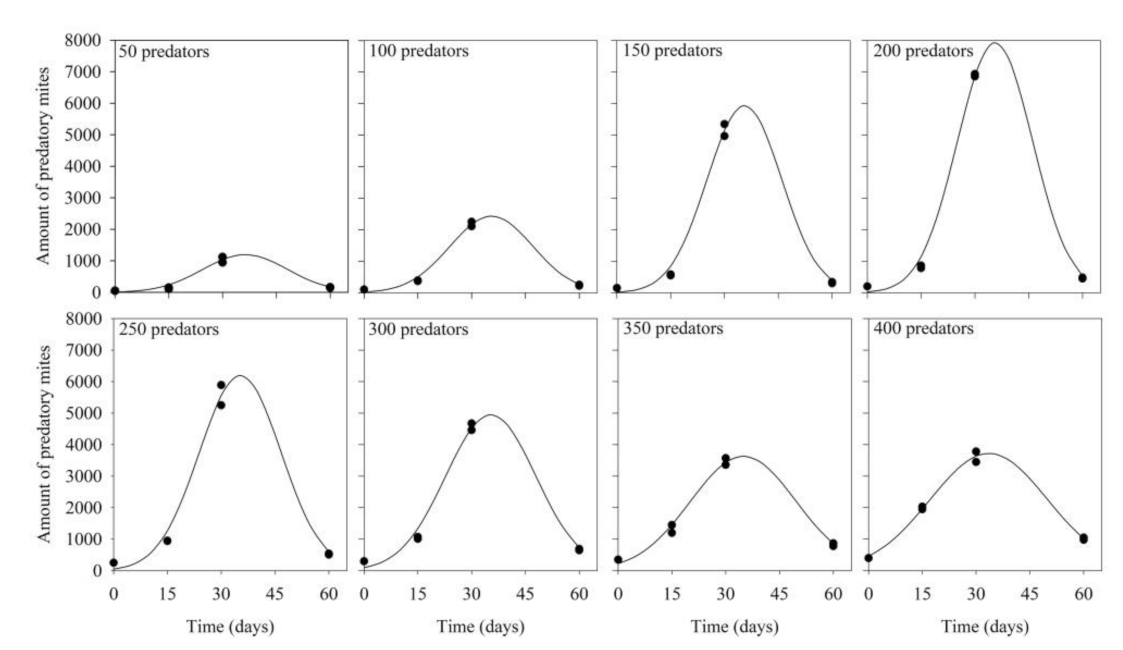
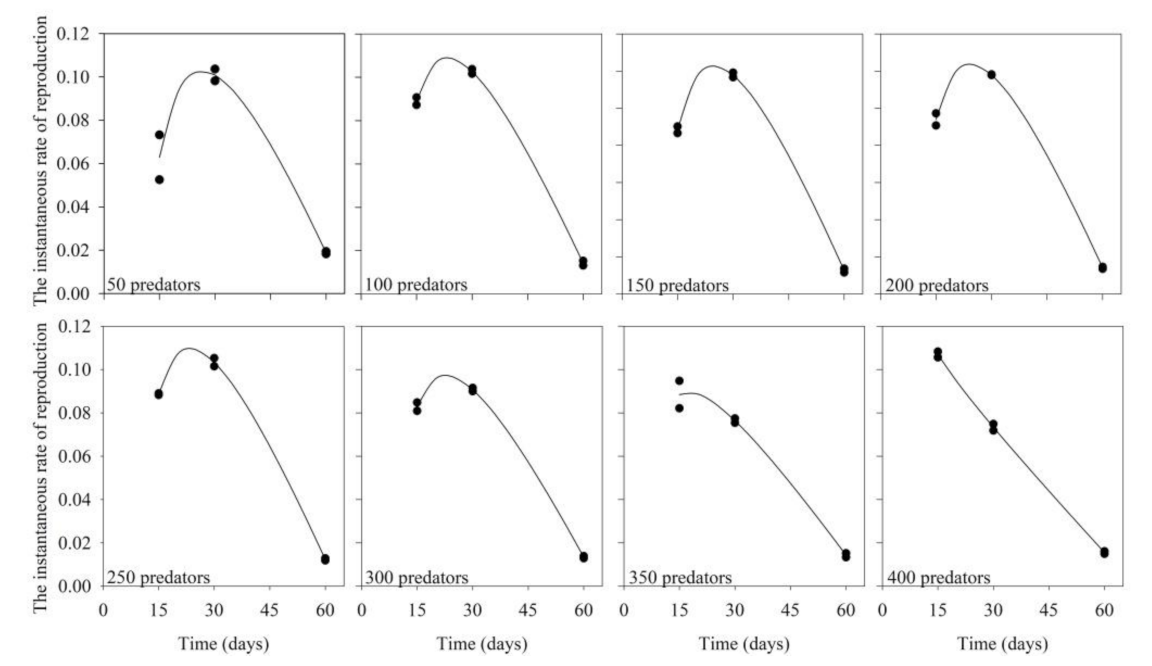
3.1. Predator Inoculation Density × Production Period
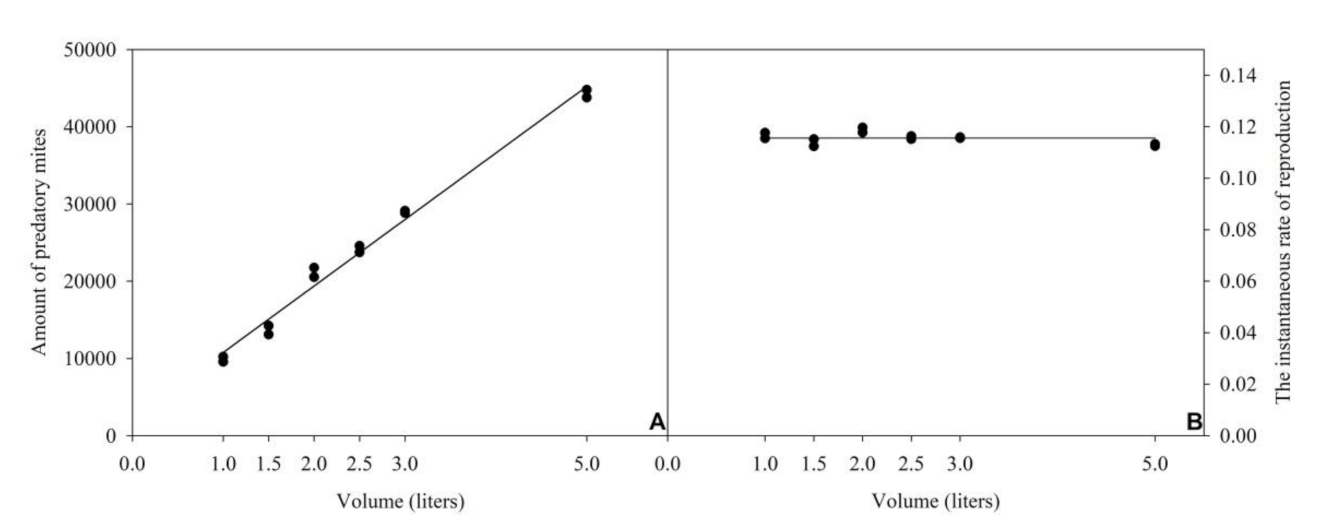
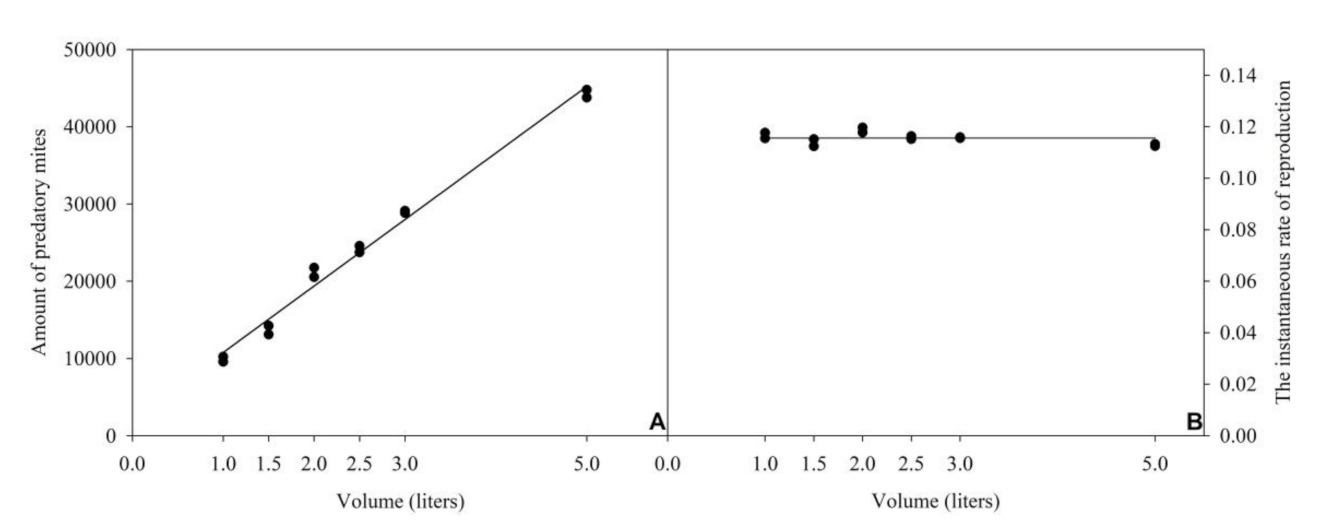
3.2. Volume of the Rearing Unit
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gil, H.K.; Garg, H. Pesticides: Environmental Impacts and Management Strategies. IntechOpen 2014, 187–230. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, D.; Moraes, G.J.; Peña, J.E. Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2015; p. 328. [Google Scholar]
- Hoy, M.; Castro, D. Two methods for large scale production of pesticide-resistant strains of the spider mite predator Metaseiulus occidentalis (Nesbitt) (Acarina, Phytoseiidae). J. Appl. Entomol. 1982, 94, 1–9. [Google Scholar] [CrossRef]
- Steiner, M.; Goodwin, S.; Welham, T. A simplified rearing method for Stratiolaelaps (=Hypoaspis) miles (Acari: Laelapidae). IOBC/WPRS Bull. 1999, 22, 241–242. [Google Scholar]
- Huang, H.; Xu, X.; LV, J.; Li, G.; Wang, E.; Gao, Y. Impact of proteins and saccharides on mass production of Tyrophagus putrescentiae (Acari: Acaridae) and its predator Neoseiulus barkeri (Acari: Phytoseiidae). Biocontrol Sci. Technol. 2013, 23, 1231–1244. [Google Scholar] [CrossRef]
- Ramakers, P.M.J.; Van Leiburg, M.J. Start of commercial production and introduction of Amblyseius mckenziei Sch. & Pr. (Acarina: Phytoseiidae) for control of Thrips tabaci Lind. (Thysanoptera: Thripidae) in glasshouses. Meded. Fac. Landbouwwet. 1982, 47, 541–545. [Google Scholar]
- Pijnakker, J.; Vangasbek, D.; Duarte, M.; Moerkens, R.; Wäckers, F.L. Predators and parasitoids-in-first: From inundative releases to preventative biological control in greenhouse crops. Front. Sustain. Food. Syst. 2020, 4, 245. [Google Scholar] [CrossRef]
- Brown, J.K.; Frohlich, D.R.; Rosell, R.C. The sweet-potato or silver-leaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization—EPPO. Commercially Used Biological Control Agents: Arachnida, Acarina—2016. Available online: https://archives.eppo.int/EPPOStandards/biocontrol_web/acarina.htm (accessed on 12 June 2020).
- Nappo—North American Plant Protection Organization. Regional Standards for Phytosanitary Measures—2015. Available online: https://www.nappo.org/application/files/5115/8650/2721/RSPM7_05-12-2014-e.pdf (accessed on 15 February 2017).
- Cavalcante, A.C.; Santos, V.L.V.; Rossi, L.C.; Moraes, G.J. Potential of five Brazilian populations of Phytoseiidae (Acari) for the biological control of Bemisia tabaci (Insecta: Hemiptera). J. Econ. Entomol. 2015, 108, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Demite, P.R.; Moraes, G.J.; McMurtry, J.A.; Denmark, H.A.; Castilho, R.C. Phytoseiidae Database. Available online: http://www.lea.usp.br/phytoseiidae/ (accessed on 28 February 2017).
- McMurtry, J.A.; Scriven, G.T. Insectary production of phytoseiid mites. J. Econ. Entomol. 1965, 58, 282–284. [Google Scholar] [CrossRef]
- Massaro, M.; Martin, J.P.I.; Moraes, J.G. Factitious food for mass production of predaceous phytoseiid mites (Acari: Phytoseiidae) commonly found in Brazil. Exp. Appl. Acarol. 2016, 70, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Freire, R.A.P.; Moraes, J.G. Mass production of the predatory mites Stratiolaelaps scimitus (Womersley) (Acari: Laelapidae). Syst. Appl. Acarol. 2007, 12, 117–119. [Google Scholar]
- Hall, D.J. An experimental approach to the dynamics of a natural population of Daphnia galeata mendotae. Ecology 1964, 45, 94–112. [Google Scholar] [CrossRef]
- Cavalcante, A.C.C.; Mandro, M.E.A.; Paes, E.R.; Moraes, G.J. Amblyseius tamatavensis Blommers (Acari: Phytoseiidae) a candidate for biological control of Bemisia tabaci (Gennadius) biotype B (Hemiptera: Aleyrodidae) in Brazil. Int. J. Acarol. 2017, 43, 10–15. [Google Scholar] [CrossRef]
- Barbosa, M.F.d.C.; O’Connor, B.M.; Moraes, G.J. A new species of Thyreophagous (Acari: Acaridae) from Brazil, with notes on species associated with stored food and human habitats and a key to species of this genus. Zootaxa 2016, 4088, 279–291. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Levels of Preda- tor Inoculation | Equation (lny = a + bx + cx2) | F Value | R2 | p-Value | ||
|---|---|---|---|---|---|---|
| A | B | C | ||||
| 50 | 2.41 | 0.25 | −0.003 | 74.47 | 0.96 | 0.00019 |
| 100 | 3.08 | 0.26 | −0.003 | 259.57 | 0.99 | 0.00001 |
| 150 | 2.86 | 0.32 | −0.004 | 316.88 | 0.99 | 0.00001 |
| 200 | 3.20 | 0.32 | −0.004 | 529.79 | 0.99 | <0.00001 |
| 250 | 3.93 | 0.27 | −0.003 | 186.51 | 0.98 | 0.00002 |
| 300 | 4.56 | 0.22 | −0.003 | 193.10 | 0.98 | 0.00002 |
| 350 | 5.39 | 0.16 | −0.002 | 214.83 | 0.98 | 0.00001 |
| 400 | 6.12 | 0.12 | −0.001 | 424.77 | 0.99 | <0.00001 |
| Levels of Predator Inoculation | Equation (y = a + bx + c/x) | F Value | R2 | p-Value | ||
|---|---|---|---|---|---|---|
| a | B | c | ||||
| 50 | 0.34 | −0.004 | −3.15 | 44.33 | 0.96 | 0.00592 |
| 100 | 0.30 | −0.004 | 2.32 | 1241.12 | 0.99 | 0.00004 |
| 150 | 0.38 | −0.005 | −3.28 | 1558.45 | 0.99 | 0.00003 |
| 200 | 0.37 | −0.005 | −3.04 | 814.67 | 0.99 | 0.00008 |
| 250 | 0.31 | −0.004 | −2.41 | 1820.76 | 0.99 | 0.00002 |
| 300 | 0.26 | −0.003 | −1.86 | 1177.54 | 0.99 | 0.00005 |
| 350 | 0.17 | −0.002 | −0.76 | 112.73 | 0.98 | 0.0015 |
| 400 | 0.12 | −0.0018 | 0.18 | 1400.07 | 0.99 | 0.00004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massaro, M.; Montrazi, M.; Melo, J.W.S.; de Moraes, G.J. Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae). Insects 2021, 12, 848. https://doi.org/10.3390/insects12100848
Massaro M, Montrazi M, Melo JWS, de Moraes GJ. Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae). Insects. 2021; 12(10):848. https://doi.org/10.3390/insects12100848
Chicago/Turabian StyleMassaro, Marcela, Matheus Montrazi, José Wagner S. Melo, and Gilberto J. de Moraes. 2021. "Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae)" Insects 12, no. 10: 848. https://doi.org/10.3390/insects12100848
APA StyleMassaro, M., Montrazi, M., Melo, J. W. S., & de Moraes, G. J. (2021). Small-Scale Production of Amblyseius tamatavensis with Thyreophagus cracentiseta (Acari: Phytoseiidae, Acaridae). Insects, 12(10), 848. https://doi.org/10.3390/insects12100848