Darwin Returns to the Galapagos: Genetic and Morphological Analyses Confirm the Presence of Tramea darwini at the Archipelago (Odonata, Libellulidae)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection, DNA Extraction, and Sequencing
2.2. Genetic Analyses
2.3. Morphological Analyses
3. Results
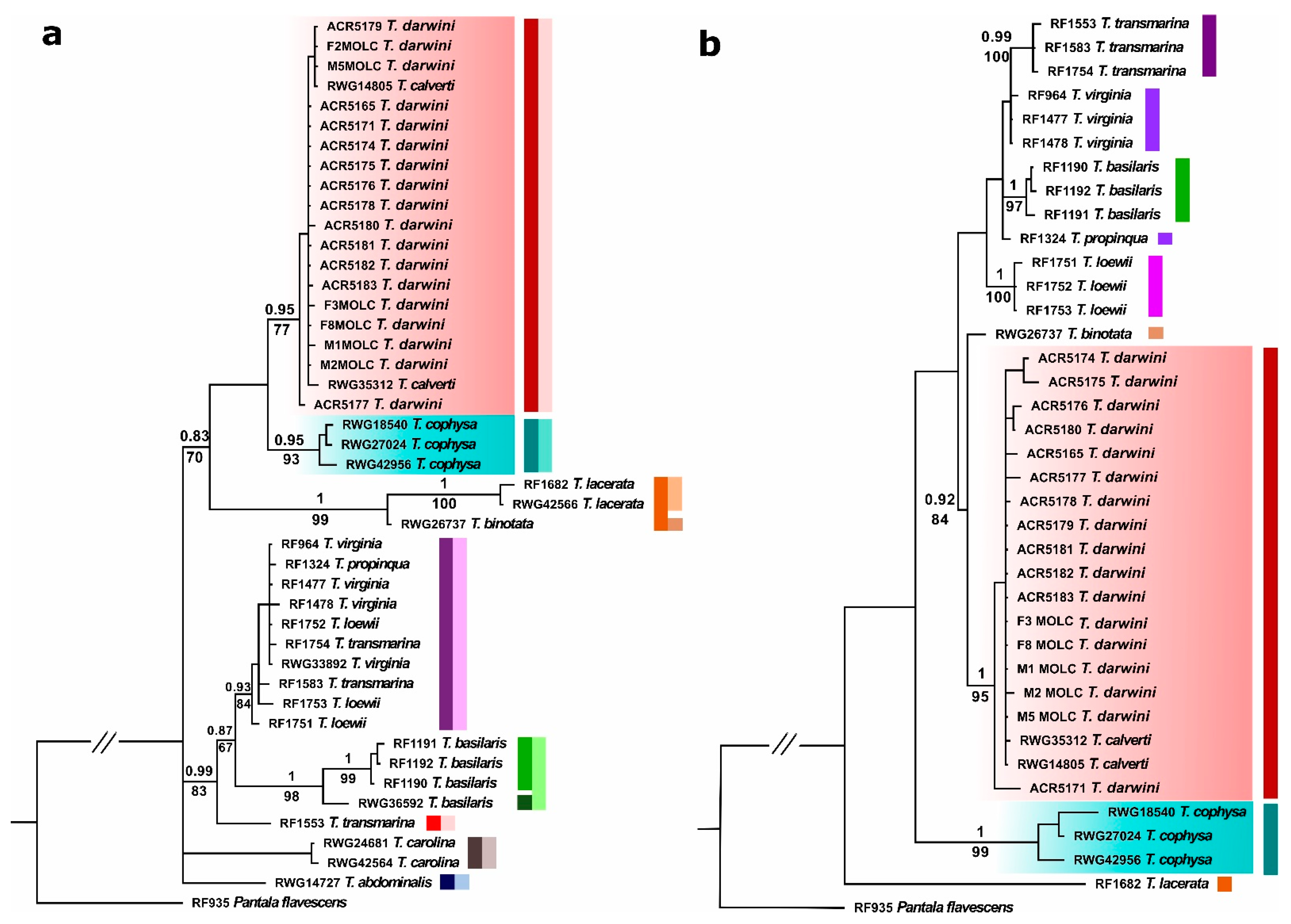
3.1. Genetic Analyses
3.2. Morphological Analyses
4. Discussion
“6a. Underside of abdomen black; abdominal segment 8 all black (Figure 6); hindwing clear with sharply edged dark basal band; male at maturity with frons all violet, male face blackTramea cophysa
6b. Underside of abdomen brown to red; abdominal segment 8 with a semicircular pale basal-lateral spot (Figure 5); hindwing tinged brown with an amber-edged basal band; male with only broad band on top of frons violet; female band on frons top narrower, lower frons and face otherwise pale; male at maturity with lower frons and face redTramea calverti”
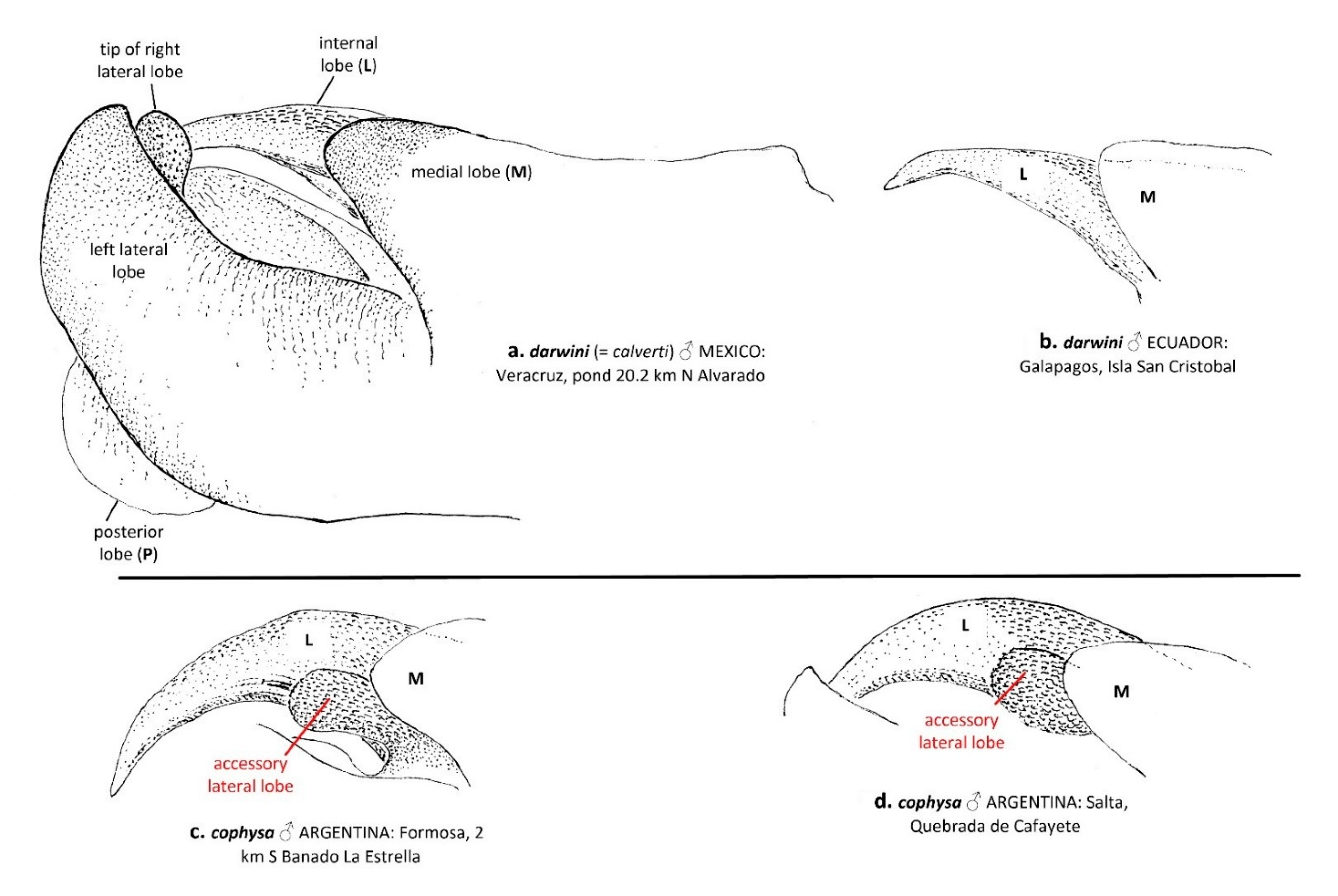
“3. Head in male with dorsal surface of vertex deep metallic violet, same color as postfrons, entire postfrons metallic violet (Figure 3a); labrum mostly dark brown (Figure 3b); vesica spermalis with a small accessory lateral lobe (Figure 2c,d); underside of abdomen black in both sexes (some females with pale brown coloration but often dusted with pruinosity basally, Figure 6c), same color as lateral carina (Figure 6a,b); abdominal segment 8 all black (Figure 7a–c); hindwing clear with sharply edged dark basal band.cophysa Hagen
– Head in male with dorsal surface of vertex orange, basal half of postfrons metallic violet (Figure 3); labrum mostly pale brown (Figure 3d,f,h,j); vesica spermalis lacking a small accessory lateral lobe (Figure 2a,b); underside of abdomen in both sexes brown to red (Figure 6d–g); abdominal segment 8 with a semicircular pale brown basal-lateral spot (Figure 7e–h); hindwing tinged brown with an amber-edged basal band.4”
5. Conclusions
- Our genetic analyses show that the Tramea species from Galapagos belongs to a clade that comprises also the continental species T. calverti, and T. cophysa appears as a closely related species.
- Concordant with the results of the genetic analyses, the morphology of the Tramea collected in Galapagos is closer to T. calverti than to T. cophysa.
- Only one morphological character has been found which consistently discriminates between both species: an accessory lobe in the male sperm vesicle, which appears in T. cophysa, but is absent in both T. calverti and the Tramea from Galapagos.
- Given these results, and following the taxonomic principle of priority, the Tramea species currently found in Galapagos should be referred to by its older name, T. darwini.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whittaker, R.J. Island Biogeography. Ecology, Evolution and Conservation; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- de Candolle, A. Géographie Botanique Raisonnée; Ou, Exposition des Faits Principaux et des Lois Concernant la Distribution Géographique des Plantes de L’epoque Actuelle; Masson et cie: Paris, France, 1855. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Darwin, C. On the Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Linsley, E.G.; Usinger, R.L. Insects of the Galápagos Islands. Proc. California Acad. Sci. 4th Ser. 1966, 33, 113–196. [Google Scholar]
- Gerecke, R.; Peck, S.B.; Pehofer, H.E. The invertebrate fauna of the inland waters of the Galapagos Archipelago (Ecuador)—A limnological and zoogeographical summary. Arch. Hydrobiol. Suppl. 1995, 107, 113–147. [Google Scholar]
- Peck, S.B. Smaller Orders of Insects of the Galápagos Islands, Ecuador: Evolution, Ecology, and Diversity; NRC Research Press: Ottawa, ON, Canada, 2001; p. 278. [Google Scholar]
- Muddeman, J. A new species for the Galapagos Islands: Great Pondhawk (Erythemis vesiculosa). Argia 2007, 19, 17–18. [Google Scholar]
- Cordero-Rivera, A.; Encalada, A.C.; Sánchez–Guillén, R.A.; Santolamazza–Carbone, S.; von Ellenrieder, N. The status of Rhionaeschna galapagoensis (Currie, 1901) with notes on its biology and a description of its ultimate instar larva (Odonata, Aeshnidae). Anim. Biodivers. Conserv. 2016, 39, 45–63. [Google Scholar] [CrossRef]
- Lorenzo-Carballa, M.O.; Hassall, C.; Encalada, A.C.; Sanmartín-Villar, I.; Torres-Cambas, Y.; Cordero-Rivera, A. Parthenogenesis did not consistently evolve in insular populations of Ischnura hastata (Odonata, Coenagrionidae). Ecol. Entomol. 2016. [Google Scholar] [CrossRef]
- May, M. A critical overview of progress in studies of migration of dragonflies (Odonata: Anisoptera), with emphasis on North America. J. Insect. Conserv. 2013, 17, 1–15. [Google Scholar] [CrossRef]
- Garrison, R.W.; von Ellenrieder, N.; Louton, J.A. Dragonfly Genera of the New World: An Illustrated and Annotated Key to the Anisoptera; The John Hopkins University Press: Baltimore, MD, USA, 2006; p. 368. [Google Scholar]
- Peck, S.B. The dragonflies and damselflies of the Galapagos Islands, Ecuador (Insecta: Odonata). Psyche 1992, 99, 309–322. [Google Scholar] [CrossRef]
- De Marmels, J.; Rácenis, J. An analysis of the cophysa-group of Tramea Hagen, with descriptions of two new species (Anisoptera: Libellulidae). Odonatologica 1982, 11, 109–128. [Google Scholar]
- Kirby, W.F. A revision of the subfamily Libellulinae, with descriptions of new genera and species. Trans. Zool. Soc. Lond. 1889, 12, 249–348. [Google Scholar] [CrossRef]
- Calvert, P.P. Odonata. In Biologia Centrali Americana: Insecta Neuroptera; R.H. Porter & Dulau Co.: London, UK, 1906; pp. 213–308. [Google Scholar]
- Ris, F. Collections Zoologiques du Baron Edm. de Selys Longchamps. Catal. Syst. Descriptif. Libellulinen 1913, 16, 965–1042. [Google Scholar]
- Currie, R.P. Papers from the Hopkins Stanford Galapagos Expedition, 1898–1899. III. Proc. Wash. Acad. Sci. 1901, 3, 381–389. [Google Scholar]
- Campos, R.F. Catálogo Sistemático y Sinonímico de los Odonatos del Ecuador. Rev. Col. Nac. Vicente Rocafuerte 1922, 4, 1–75. [Google Scholar]
- Calvert, P.P. Odonata of voyages under the auspices of the New York Zoological Society. Entomol. News 1947, 58, 227–230. [Google Scholar]
- Asahina, S. Dragonflies taken by Dr. Sekiguchi in the Galapagos Islands. Publ. Entomol. Lab. Univ. Osaka Prefect. 1961, 6, 1–3. [Google Scholar]
- Gloger, H. Bemerkungen über die Odonaten-Fauna der Galapagos-Inseln nach der Ausbeute von Juan Foerster, 1959. Opusc. Zool. 1964, 74, 1–6. [Google Scholar]
- Palumbi, S.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G.R. The Simple Fool’s Guide to PCR, 2nd ed.; University of Hawaii Press: Honolulu, HI, USA, 1991; p. 45. [Google Scholar]
- Futahashi, R. A revisional study of Japanese dragonflies based on DNA analysis. TOMBO Acta Odonatol. Jpn. 2011, 53, 67–74. [Google Scholar]
- Dijkstra, K.D.B.; Kalkman, V.J.; Dow, R.A.; Stokvis, F.R.; Van Tol, J. Redefining the damselfly families: A comprehensive molecular phylogeny of Zygoptera (Odonata). Syst. Entomol. 2014, 39, 68–96. [Google Scholar] [CrossRef]
- Dmitriev, D.A.; Rakitov, R.A. Decoding of superimposed traces produced by direct sequencing of heterozygous indels. PLoS Comput. Biol. 2008, 4, e1000113. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrapping algorithm for the RAxML web-servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Ronquist, F.; John, P.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B 2003, 270, 96–99. [Google Scholar] [CrossRef]
- Muttkowski, R.A. Catalogue of the Odonata of North America. Bull. Public Mus. City Milwaukee 1910, 1, 1–207. [Google Scholar]
- Calvert, P.P. The Odonata of Baja California, Mexico. Proc. Calif. Acad. Sci. 2nd Ser. 1895, 4, 463–558. [Google Scholar]
- Kimmins, D.E. A list of the type-specimens of Libellulidae and Corduliidae (Odonata) in the British Museum (Natural History). Bull. Br. Mus. (Nat. Hist.) Entomol. 1969, 22, 277–305. [Google Scholar] [CrossRef]
- Dunkle, S.W. Dragonflies of the Florida Peninsula, Bermuda, and the Bahamas; Scientific Publishers: Gainesville, FL, USA, 1989; p. 154. [Google Scholar]
- Carpenter, G.H. A contribution towards a list of the dragonflies of Jamaica. J. Inst. Jamaica 1896, 2, 259–263. [Google Scholar]
- Turner, P.E., Jr. Odonata of the Galapagos Islands (Odonata). Pan-Pac. Entomol. 1967, 43, 285–291. [Google Scholar]
- Linsley, E.G. Insects of the Galápagos (Supplement). Occas. Papers Calif. Acad. Sci. 1977, 125, 1–50. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenzo-Carballa, M.O.; Garrison, R.W.; Encalada, A.C.; Cordero-Rivera, A. Darwin Returns to the Galapagos: Genetic and Morphological Analyses Confirm the Presence of Tramea darwini at the Archipelago (Odonata, Libellulidae). Insects 2021, 12, 21. https://doi.org/10.3390/insects12010021
Lorenzo-Carballa MO, Garrison RW, Encalada AC, Cordero-Rivera A. Darwin Returns to the Galapagos: Genetic and Morphological Analyses Confirm the Presence of Tramea darwini at the Archipelago (Odonata, Libellulidae). Insects. 2021; 12(1):21. https://doi.org/10.3390/insects12010021
Chicago/Turabian StyleLorenzo-Carballa, María Olalla, Rosser W. Garrison, Andrea C. Encalada, and Adolfo Cordero-Rivera. 2021. "Darwin Returns to the Galapagos: Genetic and Morphological Analyses Confirm the Presence of Tramea darwini at the Archipelago (Odonata, Libellulidae)" Insects 12, no. 1: 21. https://doi.org/10.3390/insects12010021
APA StyleLorenzo-Carballa, M. O., Garrison, R. W., Encalada, A. C., & Cordero-Rivera, A. (2021). Darwin Returns to the Galapagos: Genetic and Morphological Analyses Confirm the Presence of Tramea darwini at the Archipelago (Odonata, Libellulidae). Insects, 12(1), 21. https://doi.org/10.3390/insects12010021