Body Size Differences between Foraging and Intranidal Workers of the Monomorphic Ant Lasius niger


Abstract
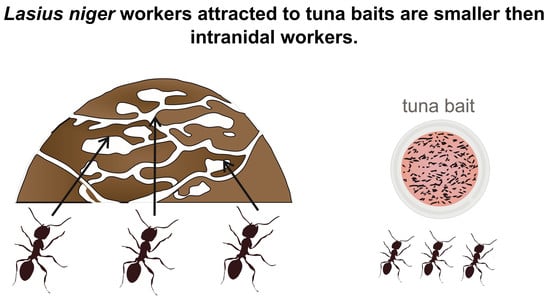
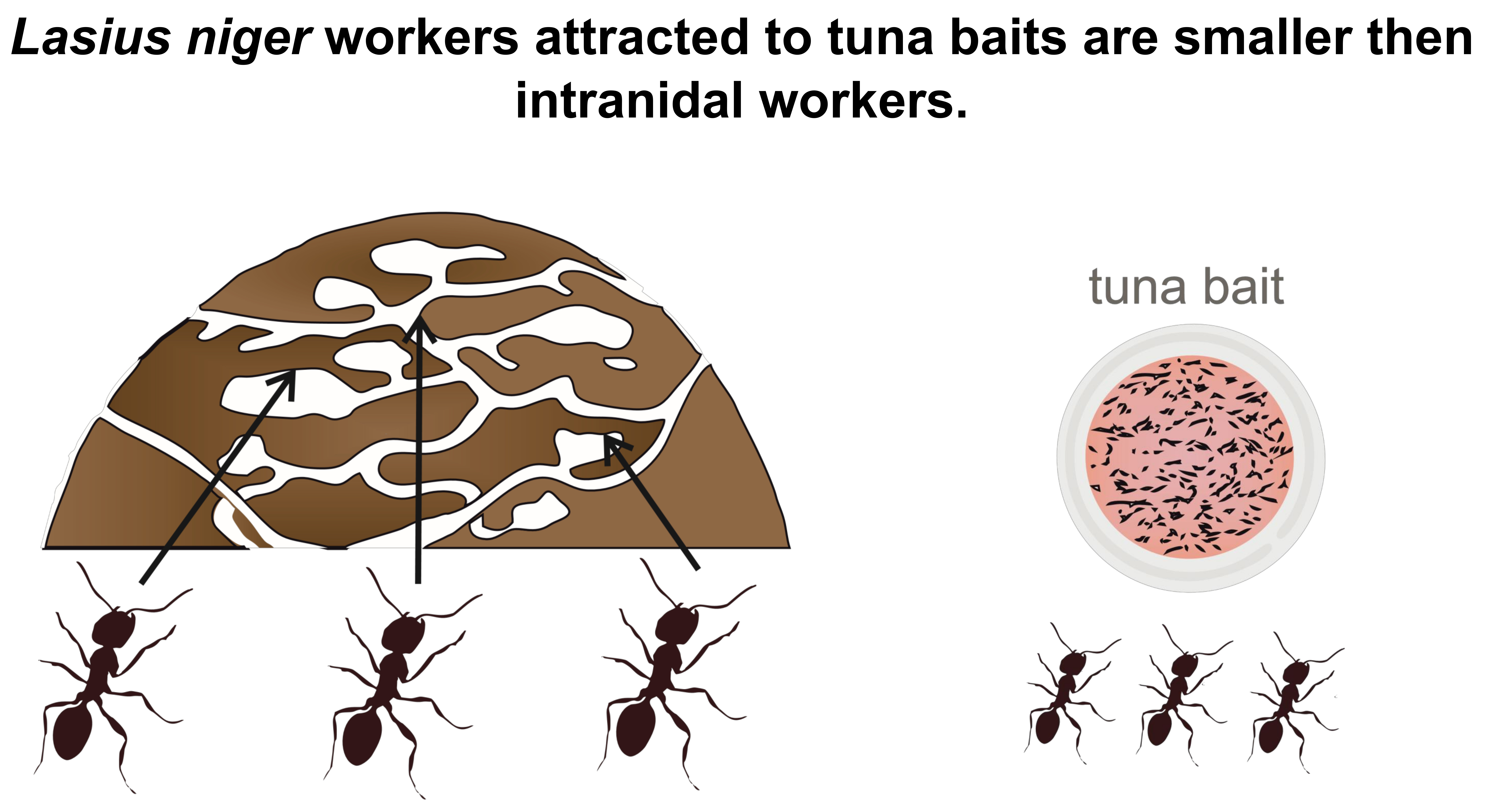
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Sampling
2.3. Morphological Measurements
2.4. Statistical Procedures
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Billick, I. The relationship between the distribution of worker sizes and new worker production in the ant Formica neorufibarbis. Oecologia 2002, 132, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Sagili, R.R.; Pankiw, T.; Metz, B.N. Division of labor associated with brood rearing in the honey bee: How does it translate to colony fitness? PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Colin, T.; Doums, C.; Peronnet, R.; Molet, M. Decreasing worker size diversity does not affect colony performance during laboratory challenges in the ant Temnothorax nylanderi. Behav. Ecol. Sociobiol. 2017, 71, 92. [Google Scholar] [CrossRef]
- Gruter, C.; Menezes, C.; Imperatriz-Fonseca, V.L.; Ratnieks, F.L.W. A morphologically specialized soldier caste improves colony defense in a neotropical eusocial bee. Proc. Natl. Acad. Sci. USA 2012, 109, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Peat, J.; Stout, J.C.; Tucker, J.; Darvill, B.; Derwent, L.C.; Hughes, W.O.H. Can alloethism in workers of the bumblebee, Bombus terrestris, be explained in terms of foraging efficiency? Anim. Behav. 2002, 64, 123–130. [Google Scholar] [CrossRef]
- Spaethe, J.; Weidenmuller, A. Size variation and foraging rate in bumblebees (Bombus terrestris). Insectes Soc. 2002, 49, 142–146. [Google Scholar] [CrossRef]
- Jandt, J.M.; Dornhaus, A. Spatial organization and division of labour in the bumblebee Bombus impatiens. Anim. Behav. 2009, 77, 641–651. [Google Scholar] [CrossRef]
- O’donnell, S.; Jeanne, R.L. The roles of body size and dominance in division of labor among workers of the eusocial wasp Polybia occidentalis (Olivier) (Hymenoptera, Vespidae). J. Kans. Entomol. Soc. 1995, 68, 43–50. [Google Scholar]
- Haifig, I.; Jost, C.; Janei, V.; Costa-Leonardo, A.M. The size of excavators within a polymorphic termite species governs tunnel topology. Anim. Behav. 2011, 82, 1409–1414. [Google Scholar] [CrossRef]
- Beshers, S.N.; Traniello, J.F.A. Polyethism and the adaptiveness of worker size variation in the attine ant Trachymyrmex septentrionalis. J. Insect Behav. 1996, 9, 61–83. [Google Scholar] [CrossRef]
- Cerda, X.; Retana, J. Links between worker polymorphism and thermal biology in a thermophilic ant species. Oikos 1997, 78, 467–474. [Google Scholar] [CrossRef]
- Huang, M.H. Multi-phase defense by the big-headed ant, Pheidole obtusospinosa, against raiding army ants. J. Insect Sci. 2010, 10. [Google Scholar] [CrossRef]
- Medan, V.; Josens, R.B. Nectar foraging behaviour is affected by ant body size in Camponotus mus. J. Insect Physiol. 2005, 51, 853–860. [Google Scholar] [CrossRef]
- Arnan, X.; Ferrandiz-Rovira, M.; Pladevall, C.; Rodrigo, A. Worker size-related task partitioning in the foraging strategy of a seed-harvesting ant species. Behav. Ecol. Sociobiol. 2011, 65, 1881–1890. [Google Scholar] [CrossRef]
- Fisher, P.J.; Stradling, D.J.; Sutton, B.C.; Petrini, L.E. Microfungi in the fungus gardens of the leaf-cutting ant Atta cephalotes: A preliminary study. Mycol. Res. 1996, 100, 541–546. [Google Scholar] [CrossRef]
- Bot, A.N.M.; Currie, C.R.; Hart, A.G.; Boomsma, J. Waste management in leaf-cutting ants. Ethol. Ecol. Evol. 2001, 13, 225–237. [Google Scholar] [CrossRef]
- Hart, A.G.; Ratnieks, F.L.W. Task partitioning, division of labour and nest compartmentalisation collectively isolate hazardous waste in the leafcutting ant Atta cephalotes. Behav. Ecol. Sociobiol. 2001, 49, 387–392. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Superorganism: The Beauty, Elegance, and Strangeness of Insect Societies; WW Norton & Company: New York, NY, USA, 2008. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Belknap Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Patel, A.D. An unusually broad behavioral repertory for a major worker in a dimorphic ant species: Pheidole morrisi (Hymenoptera, Formicidae). Psyche A J. Entomol. 1990, 97. [Google Scholar] [CrossRef]
- Herbers, J.M.; Cunningham, M. Social organization in Lepthotorax longispinosus Mayr. Anim. Behav. 1983, 31, 759–771. [Google Scholar] [CrossRef]
- Westling, J.N.; Harrington, K.; Bengston, S.; Dornhaus, A. Morphological differences between extranidal and intranidal workers in the ant Temnothorax rugatulus, but no effect of body size on foraging distance. Insectes Soc. 2014, 61, 367–369. [Google Scholar] [CrossRef]
- Grześ, I.M.; Okrutniak, M.; Grzegorzek, J. The size-dependent division of labour in monomorphic ant Lasius niger. Eur. J. Soil Biol. 2016, 77, 1–3. [Google Scholar] [CrossRef]
- Vele, A.; Modlinger, R. Body size of wood ant workers affects their work division. Sociobiology 2019, 66, 614–618. [Google Scholar] [CrossRef]
- Czechowski, W.; Radchenko, A.; Czechowska, W.; Vepsäläinen, K. The Ants of Poland with Reference to the Myrmecofauna of Europe; Museum and Institute of Zoology of the Polish Academy of Sciences and Natura optima dux Foundation: Warszawa, Poland, 2012. [Google Scholar]
- Offenberg, J. Balancing between mutualism and exploitation: The symbiotic interaction between Lasius ants and aphids. Behav. Ecol. Sociobiol. 2001, 49, 304–310. [Google Scholar] [CrossRef]
- Engel, V.; Fischer, M.K.; Wackers, F.L.; Volkl, W. Interactions between extrafloral nectaries, aphids and ants: Are there competition effects between plant and homopteran sugar sources? Oecologia 2001, 129, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Chadab, R.; Rettenmeyer, C.W. Mass recruitment by army ants. Science 1975, 188, 1124–1125. [Google Scholar] [CrossRef]
- Deslippe, R.J.; Savolainen, R. Role of food-supply in structuring a population of Formica ants. J. Anim. Ecol. 1994, 63, 756–764. [Google Scholar] [CrossRef]
- Yamauchi, K.; Yoshida, T.; Ogawa, T.; Itoh, S.; Ogawa, Y.; Jimbo, S.; Imai, H.T. Spermatogenesis of diploid males in the formicine ant, Lasius sakagamii. Insectes Soc. 2001, 48, 28–32. [Google Scholar] [CrossRef]
- Fjerdingstad, E.J. Control of body size of Lasius niger ant sexuals—Worker interests, genes and environment. Mol. Ecol. 2005, 14, 3123–3132. [Google Scholar] [CrossRef]
- Aron, S.; Steinhauer, N.; Fournier, D. Influence of queen phenotype, investment and maternity apportionment on the outcome of fights in cooperative foundations of the ant Lasius niger. Anim. Behav. 2009, 77, 1067–1074. [Google Scholar] [CrossRef]
- Mailleux, A.C.; Deneubourg, J.L.; Detrain, C. How do ants assess food volume? Anim. Behav. 2000, 59, 1061–1069. [Google Scholar] [CrossRef]
- Detrain, C.; Prieur, J. Sensitivity and feeding efficiency of the black garden ant Lasius niger to sugar resources. J. Insect Physiol. 2014, 64, 74–80. [Google Scholar] [CrossRef]
- Portha, S.; Deneubourg, J.L.; Detrain, C. How food type and brood influence foraging decisions of Lasius niger scouts. Anim. Behav. 2004, 68, 115–122. [Google Scholar] [CrossRef]
- Beckers, R.; Deneubourg, J.L.; Goss, S. Modulation of trail laying in the ant Lasius niger (Hymenoptera, formicidae) and its role in the collective selection of a food source. J. Insect Behav. 1993, 6, 751–759. [Google Scholar] [CrossRef]
- Portha, S.; Deneubourg, J.L.; Detrain, C. Self-organized asymmetries in ant foraging: A functional response to food type and colony needs. Behav. Ecol. 2002, 13, 776–781. [Google Scholar] [CrossRef]
- Detrain, C.; Verheggen, F.J.; Diez, L.; Wathelet, B.; Haubruge, E. Aphid-ant mutualism: How honeydew sugars influence the behaviour of ant scouts. Physiol. Entomol. 2010, 35, 168–174. [Google Scholar] [CrossRef]
- Kay, A.; Rissing, S.W. Division of foraging labor in ants can mediate demands for food and safety. Behav. Ecol. Sociobiol. 2005, 58, 165–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
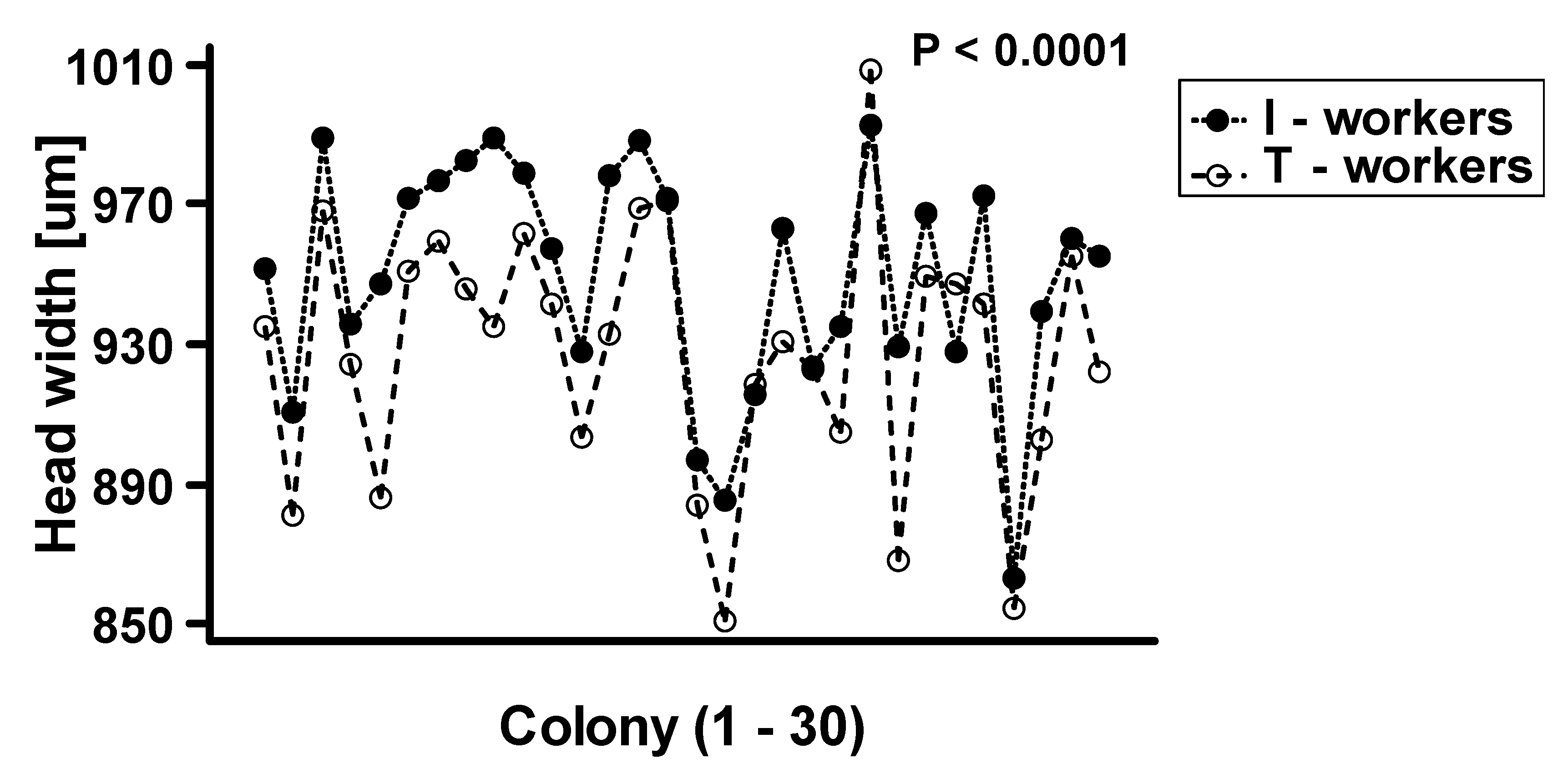
| Source | Df | F-Ratio | p-Value |
|---|---|---|---|
| Main Effects | |||
| Colony | 29 | 51.36 | p < 0.0001 |
| Worker Class | 1 | 162.7 | p < 0.0001 |
| Interaction | |||
| Colony*Worker Class | 29 | 4.294 | p < 0.0001 |
| Residual | 2845 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okrutniak, M.; Rom, B.; Turza, F.; Grześ, I.M. Body Size Differences between Foraging and Intranidal Workers of the Monomorphic Ant Lasius niger. Insects 2020, 11, 433. https://doi.org/10.3390/insects11070433
Okrutniak M, Rom B, Turza F, Grześ IM. Body Size Differences between Foraging and Intranidal Workers of the Monomorphic Ant Lasius niger. Insects. 2020; 11(7):433. https://doi.org/10.3390/insects11070433
Chicago/Turabian StyleOkrutniak, Mateusz, Bartosz Rom, Filip Turza, and Irena M. Grześ. 2020. "Body Size Differences between Foraging and Intranidal Workers of the Monomorphic Ant Lasius niger" Insects 11, no. 7: 433. https://doi.org/10.3390/insects11070433
APA StyleOkrutniak, M., Rom, B., Turza, F., & Grześ, I. M. (2020). Body Size Differences between Foraging and Intranidal Workers of the Monomorphic Ant Lasius niger. Insects, 11(7), 433. https://doi.org/10.3390/insects11070433